
Laminin α4 Expression in Human Adipose Tissue Depots and Its Association with Obesity and Obesity Related Traits
 , , , ,
, , , ,  and
and 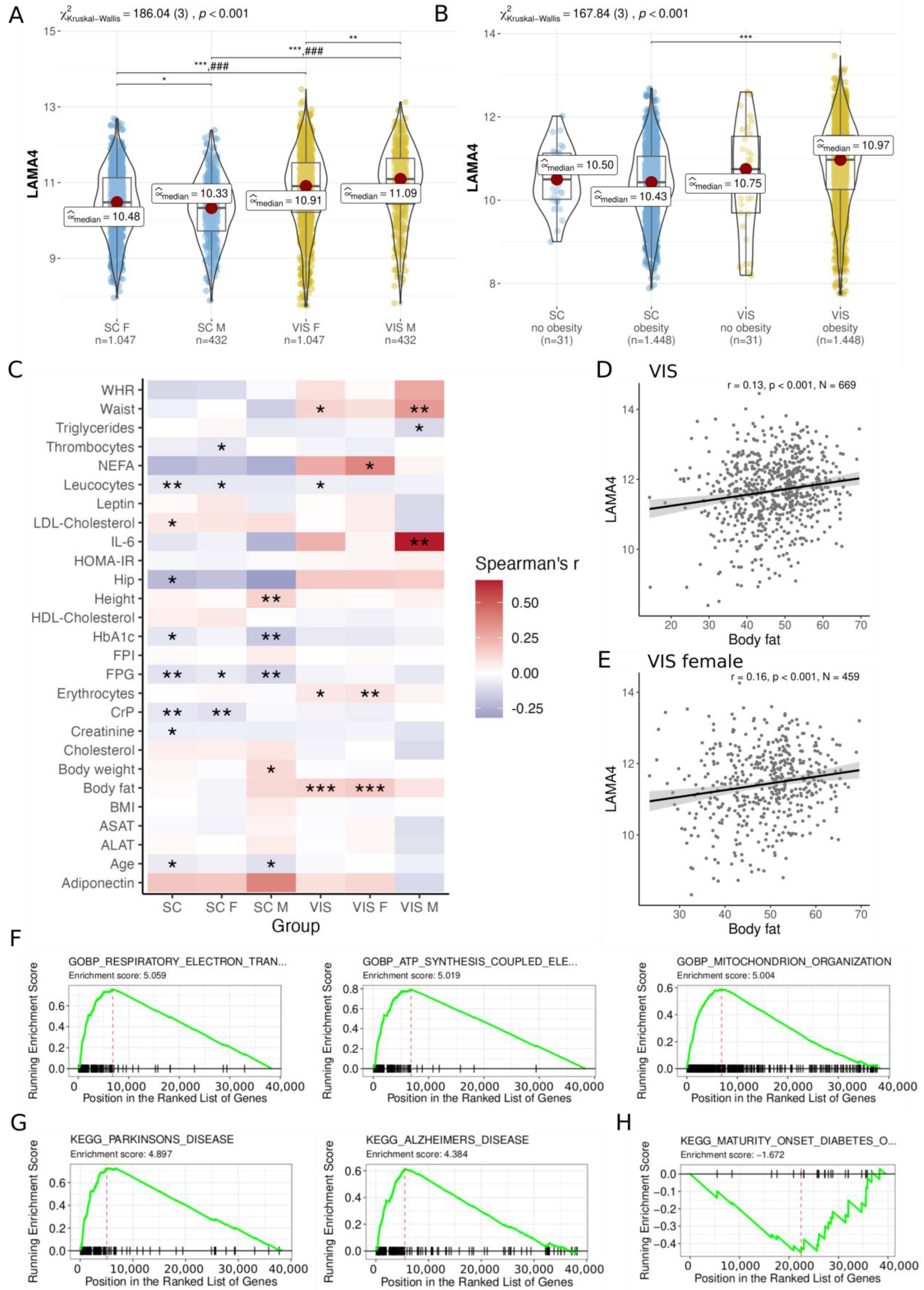{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design of the Leipzig Obesity BioBank
2.2. Bulk RNA-Sequencing and Analysis
2.3. Statistical Analyses
3. Results
3.1. Association of LAMA4 Gene Expression with Adiposity and Gene Set Pathway Alterations
3.2. Extensive Weight Loss Does Not Strongly Affect AT LAMA4 Expression
3.3. AT LAMA4 Expression Is Not Related to Metabolically Healthy Obesity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO European Regional Obesity Report 2022; Regional Office for Europe: Copenhagen, Denmark, 2022. [Google Scholar]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Noncommunicable Diseases Progress Monitor. 2017. Available online: https://www.who.int/publications-detail-redirect/WHO-NMH-NVI-17.9 (accessed on 23 March 2023).
- Hubert, H.B.; Feinleib, M.; McNamara, P.M.; Castelli, W.P. Obesity as an independent risk factor for cardiovascular disease: A 26-year follow-up of participants in the Framingham Heart Study. Circulation 1983, 67, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, K.; Douglas, I.; Forbes, H.; dos-Santos-Silva, I.; Leon, D.A.; Smeeth, L. Body-mass index and risk of 22 specific cancers: A population-based cohort study of 5·24 million UK adults. Lancet 2014, 384, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.C. Multiple risk factors for cardiovascular disease and diabetes mellitus. Am. J. Med. 2007, 120 (Suppl. 1), S3–S11. [Google Scholar] [CrossRef] [PubMed]
- Popkin, B.M.; Du, S.; Green, W.D.; Beck, M.A.; Algaith, T.; Herbst, C.H.; Alsukait, R.F.; Alluhidan, M.; Alazemi, N.; Shekar, M. Individuals with obesity and COVID-19: A global perspective on the epidemiology and biological relationships. Obes. Rev. 2020, 21, e13128. [Google Scholar] [CrossRef] [PubMed]
- Vimercati, L.; De Maria, L.; Quarato, M.; Caputi, A.; Gesualdo, L.; Migliore, G.; Cavone, D.; Sponselli, S.; Pipoli, A.; Inchingolo, F.; et al. Association between Long COVID and Overweight/Obesity. J. Clin. Med. 2021, 10, 4143. [Google Scholar] [CrossRef]
- Yurchenco, P.D. Basement Membranes: Cell Scaffoldings and Signaling Platforms. Cold Spring Harb. Perspect. Biol. 2011, 3, a004911. [Google Scholar] [CrossRef]
- LeBleu, V.S.; MacDonald, B.; Kalluri, R. Structure and Function of Basement Membranes. Exp. Biol. Med. 2007, 232, 1121–1129. [Google Scholar] [CrossRef]
- Reggio, S.; Rouault, C.; Poitou, C.; Bichet, J.-C.; Prifti, E.; Bouillot, J.-L.; Rizkalla, S.; Lacasa, D.; Tordjman, J.; Clément, K. Increased Basement Membrane Components in Adipose Tissue During Obesity: Links with TGFβ and Metabolic Phenotypes. J. Clin. Endocrinol. Metab. 2016, 101, 2578–2587. [Google Scholar] [CrossRef]
- Yang, X.; Scott, H.A.; Monickaraj, F.; Xu, J.; Ardekani, S.; Nitta, C.F.; Cabrera, A.; McGuire, P.G.; Mohideen, U.; Das, A.; et al. Basement membrane stiffening promotes retinal endothelial activation associated with diabetes. FASEB J. 2016, 30, 601–611. [Google Scholar] [CrossRef]
- Vaicik, M.K.; Blagajcevic, A.; Ye, H.; Morse, M.C.; Yang, F.; Goddi, A.; Brey, E.M.; Cohen, R.N. The Absence of Laminin α4 in Male Mice Results in Enhanced Energy Expenditure and Increased Beige Subcutaneous Adipose Tissue. Endocrinology 2018, 159, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Noro, A.; Sillat, T.; Virtanen, I.; Ingerpuu, S.; Bäck, N.; Konttinen, Y.T.; Korhonen, M. Laminin Production and Basement Membrane Deposition by Mesenchymal Stem Cells upon Adipogenic Differentiation. J. Histochem. Cytochem. 2013, 61, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Moest, H.; Frei, A.P.; Bhattacharya, I.; Geiger, M.; Wollscheid, B.; Wolfrum, C. Malfunctioning of adipocytes in obesity is linked to quantitative surfaceome changes. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 1208–1216. [Google Scholar] [CrossRef]
- Roca-Rivada, A.; Belen Bravo, S.; Pérez-Sotelo, D.; Alonso, J.; Isabel Castro, A.; Baamonde, I.; Baltar, J.; Casanueva, F.F.; Pardo, M. CILAIR-Based Secretome Analysis of Obese Visceral and Subcutaneous Adipose Tissues Reveals Distinctive ECM Remodeling and Inflammation Mediators. Sci. Rep. 2015, 5, 12214. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yan, X.; Sun, A.; Zhang, L.; Zhang, J.; Yan, Y. Adipose extracellular matrix deposition is an indicator of obesity and metabolic disorders. J. Nutr. Biochem. 2023, 111, 109159. [Google Scholar] [CrossRef] [PubMed]
- Vaicik, M.K.; Thyboll Kortesmaa, J.; Movérare-Skrtic, S.; Kortesmaa, J.; Soininen, R.; Bergström, G.; Ohlsson, C.; Chong, L.Y.; Rozell, B.; Emont, M.; et al. Laminin α4 Deficient Mice Exhibit Decreased Capacity for Adipose Tissue Expansion and Weight Gain. PLoS ONE 2014, 9, e109854. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y. Laminin: Loss-of-function studies. Cell. Mol. Life Sci. 2017, 74, 1095–1115. [Google Scholar] [CrossRef]
- Bailey, J.L.; Burk, D.H.; Burke, S.J.; Reed, S.D.; Ghosh, S.; Elks, C.M. Adipocyte-Specific Laminin Alpha 4 Deletion Preserves Adipose Tissue Health despite Increasing Adiposity. Biomedicines 2022, 10, 2077. [Google Scholar] [CrossRef]
- Goddi, A.; Carmona, A.; Schroedl, L.; White, J.M.; Piron, M.J.; De Leon, A.; Casimiro, I.; Hoffman, A.; Porras, M.A.G.; Brey, E.M.; et al. Laminin-α4 Is Upregulated in Both Human and Murine Models of Obesity. Front. Endocrinol. 2021, 12, 698621. [Google Scholar] [CrossRef]
- Wang, M.; Li, C.; Liu, Y.; Wang, Z. Effect of LAMA4 on Prognosis and Its Correlation with Immune Infiltration in Gastric Cancer. BioMed Res. Int. 2021, 2021, 6428873. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Heiker, J.T.; Gärtner, D.; Björnson, E.; Schön, M.R.; Flehmig, G.; Klöting, N.; Krohn, K.; Fasshauer, M.; Stumvoll, M.; et al. Extensive weight loss reveals distinct gene expression changes in human subcutaneous and visceral adipose tissue. Sci. Rep. 2015, 5, 14841. [Google Scholar] [CrossRef] [PubMed]
- Langhardt, J.; Flehmig, G.; Klöting, N.; Lehmann, S.; Ebert, T.; Kern, M.; Schön, M.R.; Gärtner, D.; Lohmann, T.; Dressler, M.; et al. Effects of Weight Loss on Glutathione Peroxidase 3 Serum Concentrations and Adipose Tissue Expression in Human Obesity. Obes. Facts 2018, 11, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Metabolically Healthy Obesity. Endocr. Rev. 2020, 41, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-sensitive obesity. Am. J. Physiol.-Endocrinol. Metab. 2010, 299, E506–E515. [Google Scholar] [CrossRef] [PubMed]
- Yaskolka Meir, A.; Rinott, E.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Rosen, P.; Shelef, I.; Youngster, I.; Shalev, A.; Blüher, M.; et al. Effect of green-Mediterranean diet on intrahepatic fat: The DIRECT PLUS randomised controlled trial. Gut 2021, 70, 2085–2095. [Google Scholar] [CrossRef]
- Picelli, S.; Faridani, O.R.; Björklund, Å.K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 2014, 9, 171–181. [Google Scholar] [CrossRef]
- Song, Y.; Milon, B.; Ott, S.; Zhao, X.; Sadzewicz, L.; Shetty, A.; Boger, E.T.; Tallon, L.J.; Morell, R.J.; Mahurkar, A.; et al. A comparative analysis of library prep approaches for sequencing low input translatome samples. BMC Genom. 2018, 19, 696. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Frankish, A.; Diekhans, M.; Ferreira, A.-M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 2019, 47, D766–D773. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Nie, J.; Sicotte, H.; Li, Y.; Eckel-Passow, J.E.; Dasari, S.; Vedell, P.T.; Barman, P.; Wang, L.; Weinshiboum, R.; et al. Measure transcript integrity using RNA-seq data. BMC Bioinform. 2016, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.I.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Lauss, M. swamp: Visualization, Analysis and Adjustment of High-Dimensional Data in Respect to Sample Annotations. R Package Version 1.5.1. 2019. Available online: https://CRAN.R-project.org/package=swamp (accessed on 23 March 2022).
- Miller, H.E.; Bishop, A.J.R. Correlation AnalyzeR: Functional predictions from gene co-expression correlations. BMC Bioinform. 2021, 22, 206. [Google Scholar] [CrossRef]
- Patil, I. Visualizations with statistical details: The “ggstatsplot” approach. J. Open Source Softw. 2021, 6, 3167. [Google Scholar] [CrossRef]
- Herve, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-81-2. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 23 March 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Wang, W.; Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagemann, T.; Czechowski, P.; Ghosh, A.; Sun, W.; Dong, H.; Noé, F.; Wolfrum, C.; Blüher, M.; Hoffmann, A. Laminin α4 Expression in Human Adipose Tissue Depots and Its Association with Obesity and Obesity Related Traits. Biomedicines 2023, 11, 2806. https://doi.org/10.3390/biomedicines11102806
Hagemann T, Czechowski P, Ghosh A, Sun W, Dong H, Noé F, Wolfrum C, Blüher M, Hoffmann A. Laminin α4 Expression in Human Adipose Tissue Depots and Its Association with Obesity and Obesity Related Traits. Biomedicines. 2023; 11(10):2806. https://doi.org/10.3390/biomedicines11102806
Chicago/Turabian StyleHagemann, Tobias, Paul Czechowski, Adhideb Ghosh, Wenfei Sun, Hua Dong, Falko Noé, Christian Wolfrum, Matthias Blüher, and Anne Hoffmann. 2023. "Laminin α4 Expression in Human Adipose Tissue Depots and Its Association with Obesity and Obesity Related Traits" Biomedicines 11, no. 10: 2806. https://doi.org/10.3390/biomedicines11102806
APA StyleHagemann, T., Czechowski, P., Ghosh, A., Sun, W., Dong, H., Noé, F., Wolfrum, C., Blüher, M., & Hoffmann, A. (2023). Laminin α4 Expression in Human Adipose Tissue Depots and Its Association with Obesity and Obesity Related Traits. Biomedicines, 11(10), 2806. https://doi.org/10.3390/biomedicines11102806