
Targeting Neurogenesis in Seeking Novel Treatments for Ischemic Stroke

{kind=link}
Abstract
:1. Introduction
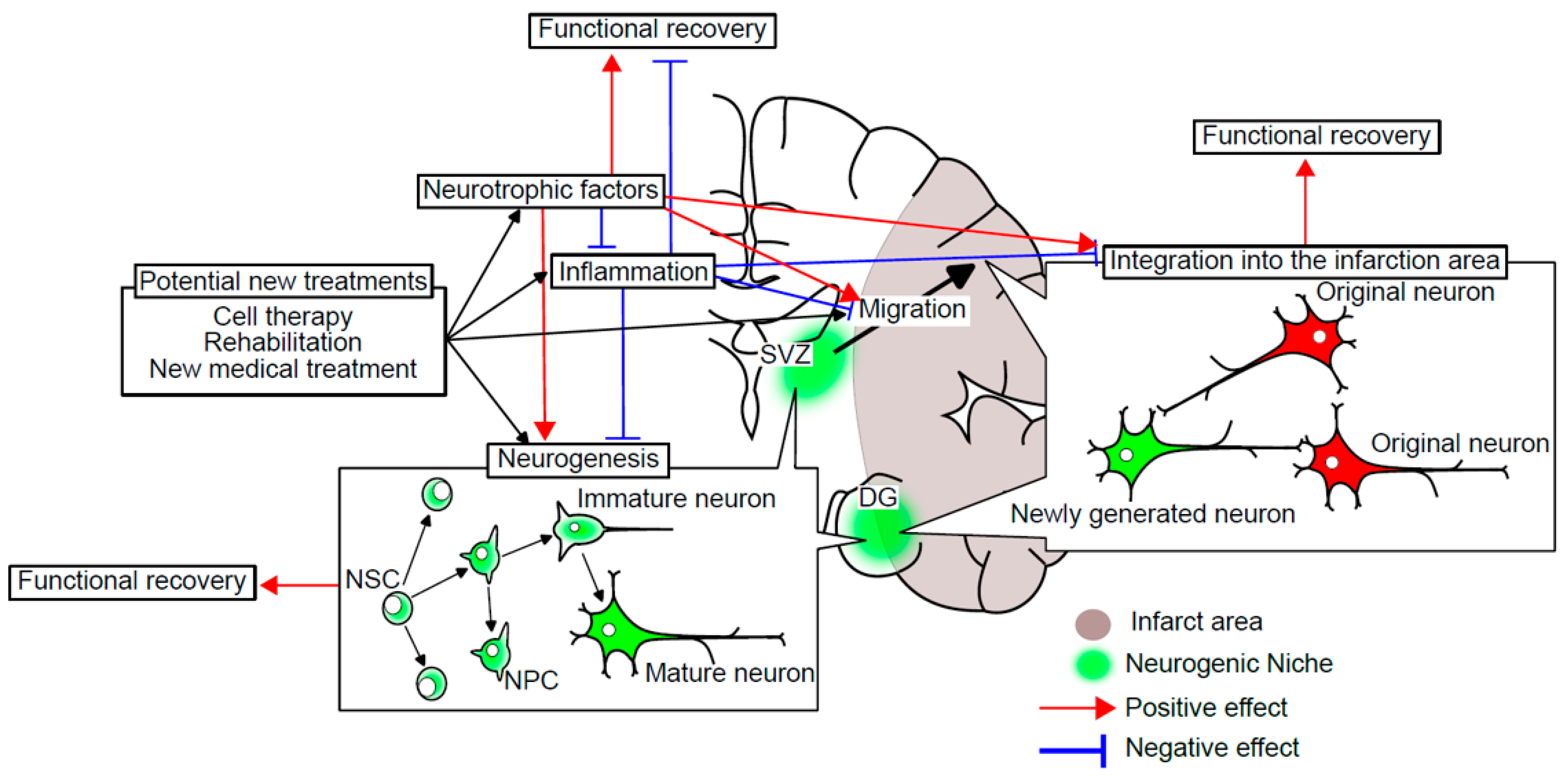
2. The Relationship between Neurogenesis and Ischemic Stroke
2.1. The Mechanism of Ischemic Cell Death
2.2. Change in Neurogenesis after Cerebral Infarction
2.3. How Neurogenesis Contributes to Functional Recovery from Ischemic Stroke
3. Neurogenesis in New Treatments for Ischemic Stroke
3.1. Cell Therapy
3.1.1. MSCs
3.1.2. Neural Stem Cells (NSCs) and NPCs
3.1.3. Other Types of Cell Therapy
3.2. Rehabilitation
3.3. Pharmacotherapy
3.4. Other Types of Interventions
4. Potential and Prospects of Neurogenesis in Treatment for Ischemic Stroke
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goyal, M.; Demchuk, A.M.; Menon, B.K.; Eesa, M.; Rempel, J.L.; Thornton, J.; Roy, D.; Jovin, T.G.; Willinsky, R.A.; Sapkota, B.L.; et al. Randomized Assessment of Rapid Endovascular Treatment of Ischemic Stroke. N. Engl. J. Med. 2015, 372, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Feigin, V.L.; Norrving, B.; Mensah, G.A. Global Burden of Stroke. Circ. Res. 2017, 120, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Z.; Szeto, V.; Bao, M.H.; Sun, H.S.; Feng, Z.P. Pharmacological approaches promoting stem cell-based therapy following ischemic stroke insults. Acta Pharmacol. Sin. 2018, 39, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.-M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Gao, J.; Yao, M.; Chang, D.; Liu, J. mTOR (Mammalian Target of Rapamycin): Hitting the Bull’s Eye for Enhancing Neurogenesis after Cerebral Ischemia? Stroke 2023, 54, 279–285. [Google Scholar] [CrossRef]
- Alvarez-Buylla, A.; GarcíA-Verdugo, J.M. Neurogenesis in Adult Subventricular Zone. J. Neurosci. 2002, 22, 629–634. [Google Scholar] [CrossRef]
- Cheatwood, J.L.; Emerick, A.J.; Kartje, G.L. Neuronal plasticity and functional recovery after ischemic stroke. Top. Stroke Rehabil. 2008, 15, 42–50. [Google Scholar] [CrossRef]
- Luo, C.X.; Lin, Y.H.; Qian, X.D.; Tang, Y.; Zhou, H.H.; Jin, X.; Ni, H.Y.; Zhang, F.Y.; Qin, C.; Li, F.; et al. Interaction of nNOS with PSD-95 negatively controls regenerative repair after stroke. J. Neurosci. 2014, 34, 13535–13548. [Google Scholar] [CrossRef]
- Göritz, C.; Frisén, J. Neural Stem Cells and Neurogenesis in the Adult. Cell Stem Cell 2012, 10, 657–659. [Google Scholar] [CrossRef]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The science of stroke: Mechanisms in search of treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef]
- Lipton, P. Ischemic cell death in brain neurons. Physiol. Rev. 1999, 79, 1431–1568. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—From mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- O’Collins, V.E.; Macleod, M.R.; Donnan, G.A.; Horky, L.L.; van der Worp, B.H.; Howells, D.W. 1,026 experimental treatments in acute stroke. Ann. Neurol. 2006, 59, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.T. The 3 Rs of Stroke Biology: Radial, Relayed, and Regenerative. Neurotherapeutics 2016, 13, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.L.; Zhao, M.; Zhao, S.S.; Xiao, T.; Song, C.G.; Cao, Y.P.; Jolkkonen, J.; Zhao, C.S. Forced limb-use enhanced neurogenesis and behavioral recovery after stroke in the aged rats. Neuroscience 2015, 286, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Parent, J.M.; Vexler, Z.S.; Gong, C.; Derugin, N.; Ferriero, D.M. Rat forebrain neurogenesis and striatal neuron replacement after focal stroke. Ann. Neurol. 2002, 52, 802–813. [Google Scholar] [CrossRef]
- Morimoto, J.; Yasuhara, T.; Kameda, M.; Umakoshi, M.; Kin, I.; Kuwahara, K.; Kin, K.; Okazaki, M.; Takeuchi, H.; Sasaki, T.; et al. Electrical Stimulation Enhances Migratory Ability of Transplanted Bone Marrow Stromal Cells in a Rodent Ischemic Stroke Model. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 57–68. [Google Scholar] [CrossRef]
- Yu, T.S.; Washington, P.M.; Kernie, S.G. Injury-Induced Neurogenesis: Mechanisms and Relevance. Neuroscientist 2016, 22, 61–71. [Google Scholar] [CrossRef]
- Duan, C.L.; Liu, C.W.; Shen, S.W.; Yu, Z.; Mo, J.L.; Chen, X.H.; Sun, F.Y. Striatal astrocytes transdifferentiate into functional mature neurons following ischemic brain injury. Glia 2015, 63, 1660–1670. [Google Scholar] [CrossRef]
- Becerra-Calixto, A.; Cardona-Gómez, G.P. The Role of Astrocytes in Neuroprotection after Brain Stroke: Potential in Cell Therapy. Front. Mol. Neurosci. 2017, 10, 88. [Google Scholar] [CrossRef]
- Rosenblum, S.; Smith, T.N.; Wang, N.; Chua, J.Y.; Westbroek, E.; Wang, K.; Guzman, R. BDNF Pretreatment of Human Embryonic-Derived Neural Stem Cells Improves Cell Survival and Functional Recovery after Transplantation in Hypoxic-Ischemic Stroke. Cell Transpl. 2015, 24, 2449–2461. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ahlenius, H.; Thored, P.; Kobayashi, R.; Kokaia, Z.; Lindvall, O. Intracerebral infusion of glial cell line-derived neurotrophic factor promotes striatal neurogenesis after stroke in adult rats. Stroke 2006, 37, 2361–2367. [Google Scholar] [CrossRef]
- Zhu, W.; Cheng, S.; Xu, G.; Ma, M.; Zhou, Z.; Liu, D.; Liu, X. Intranasal nerve growth factor enhances striatal neurogenesis in adult rats with focal cerebral ischemia. Drug Deliv. 2011, 18, 338–343. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, K.; Xie, L.; Childs, J.; Mao, X.O.; Logvinova, A.; Greenberg, D.A. VEGF-induced neuroprotection, neurogenesis, and angiogenesis after focal cerebral ischemia. J. Clin. Investig. 2003, 111, 1843–1851. [Google Scholar] [CrossRef]
- Daynac, M.; Morizur, L.; Chicheportiche, A.; Mouthon, M.A.; Boussin, F.D. Age-related neurogenesis decline in the subventricular zone is associated with specific cell cycle regulation changes in activated neural stem cells. Sci. Rep. 2016, 6, 21505. [Google Scholar] [CrossRef]
- Nada, S.E.; Tulsulkar, J.; Shah, Z.A. Heme oxygenase 1-mediated neurogenesis is enhanced by Ginkgo biloba (EGb 761®) after permanent ischemic stroke in mice. Mol. Neurobiol. 2014, 49, 945–956. [Google Scholar] [CrossRef]
- Jin, K.; Wang, X.; Xie, L.; Mao, X.O.; Zhu, W.; Wang, Y.; Shen, J.; Mao, Y.; Banwait, S.; Greenberg, D.A. Evidence for stroke-induced neurogenesis in the human brain. Proc. Natl. Acad. Sci. USA 2006, 103, 13198–13202. [Google Scholar] [CrossRef]
- Kreuzberg, M.; Kanov, E.; Timofeev, O.; Schwaninger, M.; Monyer, H.; Khodosevich, K. Increased subventricular zone-derived cortical neurogenesis after ischemic lesion. Exp. Neurol. 2010, 226, 90–99. [Google Scholar] [CrossRef]
- Ohira, K.; Furuta, T.; Hioki, H.; Nakamura, K.C.; Kuramoto, E.; Tanaka, Y.; Funatsu, N.; Shimizu, K.; Oishi, T.; Hayashi, M.; et al. Ischemia-induced neurogenesis of neocortical layer 1 progenitor cells. Nat. Neurosci. 2010, 13, 173–179. [Google Scholar] [CrossRef]
- Puderbaugh, M.; Emmady, P.D. Neuroplasticity. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Rahman, A.A.; Amruta, N.; Pinteaux, E.; Bix, G.J. Neurogenesis after Stroke: A Therapeutic Perspective. Transl. Stroke Res. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Koh, S.H.; Park, H.H. Neurogenesis in Stroke Recovery. Transl. Stroke Res. 2017, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.H.; Lo, E.H. The Role of the PI3K Pathway in the Regeneration of the Damaged Brain by Neural Stem Cells after Cerebral Infarction. J. Clin. Neurol. 2015, 11, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Varela-Nallar, L.; Inestrosa, N.C. Wnt signaling in the regulation of adult hippocampal neurogenesis. Front. Cell Neurosci. 2013, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chopp, M.; Meier, D.H.; Winter, S.; Wang, L.; Szalad, A.; Lu, M.; Wei, M.; Cui, Y.; Zhang, Z.G. Sonic hedgehog signaling pathway mediates cerebrolysin-improved neurological function after stroke. Stroke 2013, 44, 1965–1972. [Google Scholar] [CrossRef]
- Zhang, X.L.; Zhang, X.G.; Huang, Y.R.; Zheng, Y.Y.; Ying, P.J.; Zhang, X.J.; Lu, X.; Wang, Y.J.; Zheng, G.Q. Stem Cell-Based Therapy for Experimental Ischemic Stroke: A Preclinical Systematic Review. Front. Cell Neurosci. 2021, 15, 628908. [Google Scholar] [CrossRef]
- Jin, J.; Tang, Y.; Li, K.; Zuo, X.; Zhan, L.; Sun, W.; Xu, E. Bone Marrow Stromal Cells Alleviate Secondary Damage in the Substantia Nigra after Focal Cerebral Infarction in Rats. Front. Cell Neurosci. 2019, 13, 338. [Google Scholar] [CrossRef]
- Oh, S.H.; Choi, C.; Chang, D.J.; Shin, D.A.; Lee, N.; Jeon, I.; Sung, J.H.; Lee, H.; Hong, K.S.; Ko, J.J.; et al. Early neuroprotective effect with lack of long-term cell replacement effect on experimental stroke after intra-arterial transplantation of adipose-derived mesenchymal stromal cells. Cytotherapy 2015, 17, 1090–1103. [Google Scholar] [CrossRef]
- Toyoshima, A.; Yasuhara, T.; Kameda, M.; Morimoto, J.; Takeuchi, H.; Wang, F.; Sasaki, T.; Sasada, S.; Shinko, A.; Wakamori, T.; et al. Intra-Arterial Transplantation of Allogeneic Mesenchymal Stem Cells Mounts Neuroprotective Effects in a Transient Ischemic Stroke Model in Rats: Analyses of Therapeutic Time Window and Its Mechanisms. PLoS ONE 2015, 10, e0127302. [Google Scholar] [CrossRef]
- Kawauchi, S.; Yasuhara, T.; Kin, K.; Yabuno, S.; Sugahara, C.; Nagase, T.; Hosomoto, K.; Okazaki, Y.; Tomita, Y.; Umakoshi, M.; et al. Transplantation of modified human bone marrow-derived stromal cells affords therapeutic effects on cerebral ischemia in rats. CNS Neurosci. Ther. 2022, 28, 1974–1985. [Google Scholar] [CrossRef]
- Wu, J.; Sun, Z.; Sun, H.S.; Wu, J.; Weisel, R.D.; Keating, A.; Li, Z.H.; Feng, Z.P.; Li, R.K. Intravenously administered bone marrow cells migrate to damaged brain tissue and improve neural function in ischemic rats. Cell Transpl. 2008, 16, 993–1005. [Google Scholar] [CrossRef]
- Yang, Z.; Cai, X.; Xu, A.; Xu, F.; Liang, Q. Bone marrow stromal cell transplantation through tail vein injection promotes angiogenesis and vascular endothelial growth factor expression in cerebral infarct area in rats. Cytotherapy 2015, 17, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, S.; Andrzejewska, A.; Lukomska, B.; Janowski, M. Neuroinflammation as a target for treatment of stroke using mesenchymal stem cells and extracellular vesicles. J. Neuroinflammation 2019, 16, 178. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, C.T.; Claasen, J.H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef]
- Monje, M.L.; Toda, H.; Palmer, T.D. Inflammatory blockade restores adult hippocampal neurogenesis. Science 2003, 302, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Wu, S.; Li, F.; Lv, L.; Han, D.; Zhao, N.; Yan, X.; Hu, S.; Xu, T. Transplantation of VEGF-mediated bone marrow mesenchymal stem cells promotes functional improvement in a rat acute cerebral infarction model. Brain Res. 2017, 1676, 9–18. [Google Scholar] [CrossRef]
- Reynolds, B.A.; Weiss, S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef]
- Gage, F.H. Mammalian neural stem cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef]
- Mattsson, B.; Sørensen, J.C.; Zimmer, J.; Johansson, B.B. Neural grafting to experimental neocortical infarcts improves behavioral outcome and reduces thalamic atrophy in rats housed in enriched but not in standard environments. Stroke 1997, 28, 1225–1231; discussion 1222–1231. [Google Scholar] [CrossRef]
- Mine, Y.; Tatarishvili, J.; Oki, K.; Monni, E.; Kokaia, Z.; Lindvall, O. Grafted human neural stem cells enhance several steps of endogenous neurogenesis and improve behavioral recovery after middle cerebral artery occlusion in rats. Neurobiol. Dis. 2013, 52, 191–203. [Google Scholar] [CrossRef]
- Zhang, B.; Joseph, B.; Saatman, K.E.; Chen, L. Intra-Arterial Delivery of Neural Stem Cells to the Rat and Mouse Brain: Application to Cerebral Ischemia. J. Vis. Exp. 2020. [Google Scholar] [CrossRef]
- Boese, A.C.; Le, Q.E.; Pham, D.; Hamblin, M.H.; Lee, J.P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.M.; Andres, R.H.; Steinberg, G.K. Optimizing the success of cell transplantation therapy for stroke. Neurobiol. Dis. 2010, 37, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Rao, M.S. Stem Cell and Precursor Cell Therapy. NeuroMolecular Med. 2002, 2, 233–250. [Google Scholar] [CrossRef] [PubMed]
- Minnerup, J.; Kim, J.B.; Schmidt, A.; Diederich, K.; Bauer, H.; Schilling, M.; Strecker, J.K.; Ringelstein, E.B.; Sommer, C.; Schöler, H.R.; et al. Effects of neural progenitor cells on sensorimotor recovery and endogenous repair mechanisms after photothrombotic stroke. Stroke 2011, 42, 1757–1763. [Google Scholar] [CrossRef]
- Chekhonin, V.P.; Lebedev, S.V.; Volkov, A.I.; Pavlov, K.A.; Ter-Arutyunyants, A.A.; Volgina, N.E.; Savchenko, E.A.; Grinenko, N.F.; Lazarenko, I.P. Activation of expression of brain-derived neurotrophic factor at the site of implantation of allogenic and xenogenic neural stem (progenitor) cells in rats with ischemic cortical stroke. Bull. Exp. Biol. Med. 2011, 150, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Horie, N.; Pereira, M.P.; Niizuma, K.; Sun, G.; Keren-Gill, H.; Encarnacion, A.; Shamloo, M.; Hamilton, S.A.; Jiang, K.; Huhn, S.; et al. Transplanted stem cell-secreted vascular endothelial growth factor effects poststroke recovery, inflammation, and vascular repair. Stem Cells 2011, 29, 274–285. [Google Scholar] [CrossRef]
- Chen, B.; Gao, X.Q.; Yang, C.X.; Tan, S.K.; Sun, Z.L.; Yan, N.H.; Pang, Y.G.; Yuan, M.; Chen, G.J.; Xu, G.T.; et al. Neuroprotective effect of grafting GDNF gene-modified neural stem cells on cerebral ischemia in rats. Brain Res. 2009, 1284, 1–11. [Google Scholar] [CrossRef]
- Duan, R.; Gao, Y.; He, R.; Jing, L.; Li, Y.; Gong, Z.; Yao, Y.; Luan, T.; Zhang, C.; Li, L.; et al. Induced Pluripotent Stem Cells for Ischemic Stroke Treatment. Front. Neurosci. 2021, 15, 628663. [Google Scholar] [CrossRef]
- Eckert, A.; Huang, L.; Gonzalez, R.; Kim, H.S.; Hamblin, M.H.; Lee, J.P. Bystander Effect Fuels Human Induced Pluripotent Stem Cell-Derived Neural Stem Cells to Quickly Attenuate Early Stage Neurological Deficits after Stroke. Stem Cells Transl. Med. 2015, 4, 841–851. [Google Scholar] [CrossRef]
- Oh, S.H.; Jeong, Y.W.; Choi, W.; Noh, J.E.; Lee, S.; Kim, H.S.; Song, J. Multimodal Therapeutic Effects of Neural Precursor Cells Derived from Human-Induced Pluripotent Stem Cells through Episomal Plasmid-Based Reprogramming in a Rodent Model of Ischemic Stroke. Stem Cells Int. 2020, 2020, 4061516. [Google Scholar] [CrossRef]
- Matsumura, H.; Marushima, A.; Ishikawa, H.; Toyomura, J.; Ohyama, A.; Watanabe, M.; Takaoka, S.; Bukawa, H.; Matsumura, A.; Matsumaru, Y.; et al. Induced Neural Cells from Human Dental Pulp Ameliorate Functional Recovery in a Murine Model of Cerebral Infarction. Stem Cell Rev. Rep. 2022, 18, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.W.; Platt, S.R.; Lau, V.W.; Grace, H.E.; Holmes, S.P.; Wang, L.; Duberstein, K.J.; Howerth, E.W.; Kinder, H.A.; Stice, S.L.; et al. Induced Pluripotent Stem Cell-Derived Neural Stem Cell Therapy Enhances Recovery in an Ischemic Stroke Pig Model. Sci. Rep. 2017, 7, 10075. [Google Scholar] [CrossRef] [PubMed]
- Abeysinghe, H.C.; Bokhari, L.; Quigley, A.; Choolani, M.; Chan, J.; Dusting, G.J.; Crook, J.M.; Kobayashi, N.R.; Roulston, C.L. Pre-differentiation of human neural stem cells into GABAergic neurons prior to transplant results in greater repopulation of the damaged brain and accelerates functional recovery after transient ischemic stroke. Stem Cell Res. Ther. 2015, 6, 186. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, Y.H.; Kim, Y.J.; Yoon, B.W. Enforced physical training promotes neurogenesis in the subgranular zone after focal cerebral ischemia. J. Neurol. Sci. 2008, 269, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.; Sandel, M.E.; Jette, A.M.; Appelman, J.; Brandt, D.E.; Cheng, P.; Teselle, M.; Delmonico, R.; Terdiman, J.F.; Rasch, E.K. Does postacute care site matter? A longitudinal study assessing functional recovery after a stroke. Arch. Phys. Med. Rehabil. 2013, 94, 622–629. [Google Scholar] [CrossRef]
- Rogers, J.M.; Duckworth, J.; Middleton, S.; Steenbergen, B.; Wilson, P.H. Elements virtual rehabilitation improves motor, cognitive, and functional outcomes in adult stroke: Evidence from a randomized controlled pilot study. J. Neuroeng. Rehabil. 2019, 16, 56. [Google Scholar] [CrossRef]
- Biernaskie, J.; Corbett, D. Enriched rehabilitative training promotes improved forelimb motor function and enhanced dendritic growth after focal ischemic injury. J. Neurosci. 2001, 21, 5272–5280. [Google Scholar] [CrossRef]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H. More hippocampal neurons in adult mice living in an enriched environment. Nature 1997, 386, 493–495. [Google Scholar] [CrossRef]
- Fabel, K.; Wolf, S.A.; Ehninger, D.; Babu, H.; Leal-Galicia, P.; Kempermann, G. Additive effects of physical exercise and environmental enrichment on adult hippocampal neurogenesis in mice. Front. Neurosci. 2009, 3, 50. [Google Scholar] [CrossRef]
- Komitova, M.; Mattsson, B.; Johansson, B.B.; Eriksson, P.S. Enriched environment increases neural stem/progenitor cell proliferation and neurogenesis in the subventricular zone of stroke-lesioned adult rats. Stroke 2005, 36, 1278–1282. [Google Scholar] [CrossRef]
- Venna, V.R.; Xu, Y.; Doran, S.J.; Patrizz, A.; McCullough, L.D. Social interaction plays a critical role in neurogenesis and recovery after stroke. Transl. Psychiatry 2014, 4, e351. [Google Scholar] [CrossRef] [PubMed]
- Hicks, A.; Jolkkonen, J. Challenges and possibilities of intravascular cell therapy in stroke. Acta Neurobiol. Exp. (Wars) 2009, 69, 1–10. [Google Scholar] [PubMed]
- Zai, L.; Ferrari, C.; Dice, C.; Subbaiah, S.; Havton, L.A.; Coppola, G.; Geschwind, D.; Irwin, N.; Huebner, E.; Strittmatter, S.M.; et al. Inosine augments the effects of a Nogo receptor blocker and of environmental enrichment to restore skilled forelimb use after stroke. J. Neurosci. 2011, 31, 5977–5988. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Li, M.Z.; Yang, L.; Feng, X.F.; Lei, J.F.; Zhang, N.; Zhao, Y.Y.; Zhao, H. The three-phase enriched environment paradigm promotes neurovascular restorative and prevents learning impairment after ischemic stroke in rats. Neurobiol. Dis. 2020, 146, 105091. [Google Scholar] [CrossRef]
- Maldonado, M.A.; Allred, R.P.; Felthauser, E.L.; Jones, T.A. Motor skill training, but not voluntary exercise, improves skilled reaching after unilateral ischemic lesions of the sensorimotor cortex in rats. Neurorehabil. Neural Repair. 2008, 22, 250–261. [Google Scholar] [CrossRef]
- Cho, K.J.; Cheon, S.Y.; Kim, G.W. Statins Promote Long-Term Recovery after Ischemic Stroke by Reconnecting Noradrenergic Neuronal Circuitry. Neural Plast. 2015, 2015, 585783. [Google Scholar] [CrossRef]
- Sakurai, K.; Isahaya, K.; Takaishi, S.; Kato, B.; Shimizu, K.; Shimomura, K.; Tokuyama, Y.; Hasegawa, Y. Effects of early statin treatment on inflammatory biomarkers and clinical deterioration in patients with acute ischemic stroke. Rinsho Shinkeigaku 2011, 51, 6–13. [Google Scholar] [CrossRef]
- Lei, S.; Zhang, P.; Li, W.; Gao, M.; He, X.; Zheng, J.; Li, X.; Wang, X.; Wang, N.; Zhang, J.; et al. Pre- and posttreatment with edaravone protects CA1 hippocampus and enhances neurogenesis in the subgranular zone of dentate gyrus after transient global cerebral ischemia in rats. ASN Neuro 2014, 6, 1759091414558417. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Zhu, X.; Jiao, W.; Yang, Y.; Wu, Y.; Yang, L.; Wang, Y. Metformin enhances neural precursor cells migration and functional recovery after ischemic stroke in mice. Exp. Brain Res. 2023, 241, 505–515. [Google Scholar] [CrossRef]
- Jin, Q.; Cheng, J.; Liu, Y.; Wu, J.; Wang, X.; Wei, S.; Zhou, X.; Qin, Z.; Jia, J.; Zhen, X. Improvement of functional recovery by chronic metformin treatment is associated with enhanced alternative activation of microglia/macrophages and increased angiogenesis and neurogenesis following experimental stroke. Brain Behav. Immun. 2014, 40, 131–142. [Google Scholar] [CrossRef]
- Hardie, D.G. AMP-activated protein kinase—An energy sensor that regulates all aspects of cell function. Genes. Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef] [PubMed]
- McCullough, L.D.; Zeng, Z.; Li, H.; Landree, L.E.; McFadden, J.; Ronnett, G.V. Pharmacological Inhibition of AMP-activated Protein Kinase Provides Neuroprotection in Stroke. J. Biol. Chem. 2005, 280, 20493–20502. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.L.; He, M.Q.; Han, X.Y.; Sun, J.Y.; Yang, M.F.; Yuan, H.; Fan, C.D.; Zhang, S.; Mao, L.L.; Li, D.W.; et al. Intranasal Delivery of Granulocyte Colony-Stimulating Factor Enhances Its Neuroprotective Effects Against Ischemic Brain Injury in Rats. Mol. Neurobiol. 2016, 53, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Mizuma, A.; Yamashita, T.; Kono, S.; Nakayama, T.; Baba, Y.; Itoh, S.; Asakura, K.; Niimi, Y.; Asahi, T.; Kanemaru, K.; et al. Phase II Trial of Intravenous Low-Dose Granulocyte Colony-Stimulating Factor in Acute Ischemic Stroke. J. Stroke Cerebrovasc. Dis. 2016, 25, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.F.; McQuillen, P.; Mu, D.; Chang, Y.; Wendland, M.; Vexler, Z.; Ferriero, D.M. Erythropoietin enhances long-term neuroprotection and neurogenesis in neonatal stroke. Dev. Neurosci. 2007, 29, 321–330. [Google Scholar] [CrossRef]
- Tsai, T.-H.; Lu, C.-H.; Wallace, C.G.; Chang, W.-N.; Chen, S.-F.; Huang, C.-R.; Tsai, N.-W.; Lan, M.-Y.; Sung, P.-H.; Liu, C.-F.; et al. Erythropoietin improves long-term neurological outcome in acute ischemic stroke patients: A randomized, prospective, placebo-controlled clinical trial. Crit. Care 2015, 19, 49. [Google Scholar] [CrossRef]
- Khodanovich, M.; Kisel, A.; Kudabaeva, M.; Chernysheva, G.; Smolyakova, V.; Krutenkova, E.; Wasserlauf, I.; Plotnikov, M.; Yarnykh, V. Effects of Fluoxetine on Hippocampal Neurogenesis and Neuroprotection in the Model of Global Cerebral Ischemia in Rats. Int. J. Mol. Sci. 2018, 19, 162. [Google Scholar] [CrossRef]
- Chen, X.M.; Wang, N.N.; Zhang, T.Y.; Wang, F.; Wu, C.F.; Yang, J.Y. Neuroprotection by sildenafil: Neuronal networks potentiation in acute experimental stroke. CNS Neurosci. Ther. 2014, 20, 40–49. [Google Scholar] [CrossRef]
- Yu, Y.H.; Kim, S.-W.; Kang, J.; Song, Y.; Im, H.; Kim, S.J.; Yoo, D.Y.; Lee, M.-R.; Park, D.-K.; Oh, J.S.; et al. Phosphodiesterase-5 Inhibitor Attenuates Anxious Phenotypes and Movement Disorder Induced by Mild Ischemic Stroke in Rats. J. Korean Neurosurg. Soc. 2022, 65, 665–679. [Google Scholar] [CrossRef]
- Silver, B.; McCarthy, S.; Lu, M.; Mitsias, P.; Russman, A.N.; Katramados, A.; Morris, D.C.; Lewandowski, C.A.; Chopp, M. Sildenafil treatment of subacute ischemic stroke: A safety study at 25-mg daily for 2 weeks. J. Stroke Cerebrovasc. Dis. 2009, 18, 381–383. [Google Scholar] [CrossRef]
- Meng, Z.; Li, M.; He, Q.; Jiang, S.; Zhang, X.; Xiao, J.; Bai, Y. Ectopic expression of human angiopoietin-1 promotes functional recovery and neurogenesis after focal cerebral ischemia. Neuroscience 2014, 267, 135–146. [Google Scholar] [CrossRef]
- Loris, Z.B.; Pieper, A.A.; Dietrich, W.D. The neuroprotective compound P7C3-A20 promotes neurogenesis and improves cognitive function after ischemic stroke. Exp. Neurol. 2017, 290, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Loris, Z.B.; Hynton, J.R.; Pieper, A.A.; Dietrich, W.D. Beneficial Effects of Delayed P7C3-A20 Treatment after Transient MCAO in Rats. Transl. Stroke Res. 2018, 9, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.Y.; Li, A.; Hoo, R.L.; Ching, Y.P.; Christie, B.R.; Lee, T.M.; Xu, A.; So, K.F. Physical exercise-induced hippocampal neurogenesis and antidepressant effects are mediated by the adipocyte hormone adiponectin. Proc. Natl. Acad. Sci. USA 2014, 111, 15810–15815. [Google Scholar] [CrossRef]
- Yu, L.; Wang, J.; Xia, Y.; Hou, W.; Yao, X.; Guo, Y.; Wang, J.; Wei, H.; Wang, S. Adiponectin Promotes Neurogenesis after Transient Cerebral Ischemia Through STAT3 Mediated BDNF Upregulation in Astrocytes. Neurochem. Res. 2023, 48, 641–657. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-Y.; Yang, Y.-R.; Chang, H.-C. The SDF1-CXCR4 Axis Is Involved in the Hyperbaric Oxygen Therapy-Mediated Neuronal Cells Migration in Transient Brain Ischemic Rats. Int. J. Mol. Sci. 2022, 23, 1780. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Liu, H.; Yan, T.; Zhuang, Z.; Jin, D.; Peng, Y. Functional electrical stimulation-facilitated proliferation and regeneration of neural precursor cells in the brains of rats with cerebral infarction. Neural Regen. Res. 2014, 9, 243–251. [Google Scholar] [CrossRef]
- Luo, J.; Zheng, H.; Zhang, L.; Zhang, Q.; Li, L.; Pei, Z.; Hu, X. High-Frequency Repetitive Transcranial Magnetic Stimulation (rTMS) Improves Functional Recovery by Enhancing Neurogenesis and Activating BDNF/TrkB Signaling in Ischemic Rats. Int. J. Mol. Sci. 2017, 18, 455. [Google Scholar] [CrossRef]
- Peng, J.J.; Sha, R.; Li, M.X.; Chen, L.T.; Han, X.H.; Guo, F.; Chen, H.; Huang, X.L. Repetitive transcranial magnetic stimulation promotes functional recovery and differentiation of human neural stem cells in rats after ischemic stroke. Exp. Neurol. 2019, 313, 1–9. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Zhu, Y.; Li, Y.; Liu, P.; Gao, B.; Tian, S.; Du, L.; Bai, Y. Post-stroke Constraint-induced Movement Therapy Increases Functional Recovery, Angiogenesis, and Neurogenesis with Enhanced Expression of HIF-1α and VEGF. Curr. Neurovasc. Res. 2017, 14, 368–377. [Google Scholar] [CrossRef]
- Sun, C.; Sun, H.; Wu, S.; Lee, C.C.; Akamatsu, Y.; Wang, R.K.; Kernie, S.G.; Liu, J. Conditional ablation of neuroprogenitor cells in adult mice impedes recovery of poststroke cognitive function and reduces synaptic connectivity in the perforant pathway. J. Neurosci. 2013, 33, 17314–17325. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagase, T.; Kin, K.; Yasuhara, T. Targeting Neurogenesis in Seeking Novel Treatments for Ischemic Stroke. Biomedicines 2023, 11, 2773. https://doi.org/10.3390/biomedicines11102773
Nagase T, Kin K, Yasuhara T. Targeting Neurogenesis in Seeking Novel Treatments for Ischemic Stroke. Biomedicines. 2023; 11(10):2773. https://doi.org/10.3390/biomedicines11102773
Chicago/Turabian StyleNagase, Takayuki, Kyohei Kin, and Takao Yasuhara. 2023. "Targeting Neurogenesis in Seeking Novel Treatments for Ischemic Stroke" Biomedicines 11, no. 10: 2773. https://doi.org/10.3390/biomedicines11102773
APA StyleNagase, T., Kin, K., & Yasuhara, T. (2023). Targeting Neurogenesis in Seeking Novel Treatments for Ischemic Stroke. Biomedicines, 11(10), 2773. https://doi.org/10.3390/biomedicines11102773