
Understanding Intervertebral Disc Degeneration: Background Factors and the Role of Initial Injury


{kind=link}
Abstract
1. Introduction
2. Embryonic Development and Maturation of IVD
3. Normal Anatomy of IVD
3.1. Nucleus Pulposus
3.2. Annulus Fibrosus
3.3. Endplate
4. Normal Physiology of IVD
5. IVD as an Organ of Immune Privilege
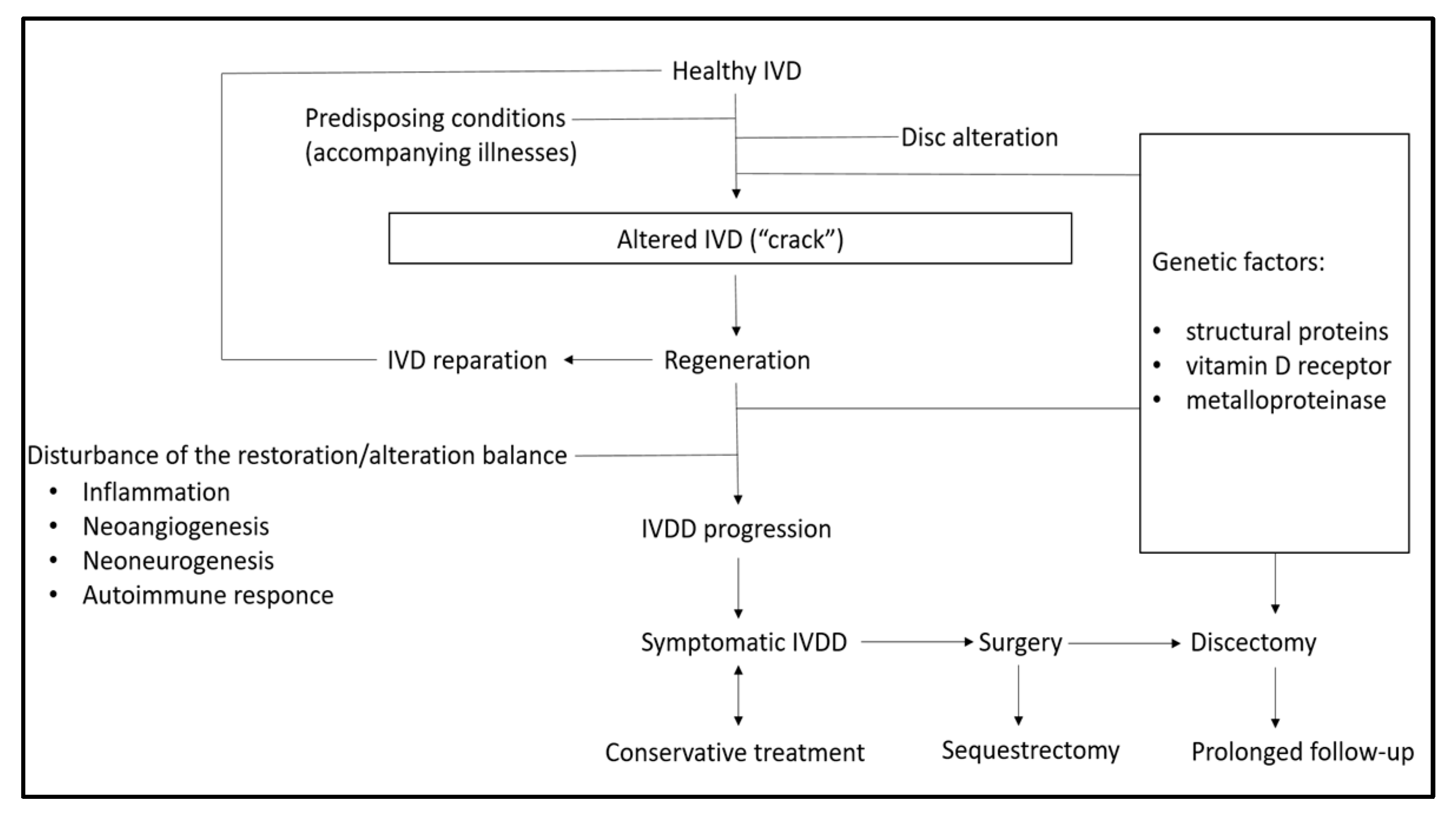
6. Pathophysiology of IVDD
6.1. Inflammation
6.2. Neoangiogenesis in IVDD Pathogenesis
6.3. Neoneurogenesis and Microglial Role in the Pathophysiology of IVDD
6.4. Metabolic Syndrome and Diabetis Mellitus
6.5. Autoimmune Response in IVDD
7. Role of Genetic Factors in the Development of IVDD
7.1. MiRNA as an Integral Indicator of IVDD Development
7.2. Structural Protein Alterations
7.3. Aggrecans
7.4. Vitamin D Receptor
7.5. Metalloproteinase
7.6. Interleukin
8. Discussion
8.1. Meniscus
8.2. Articular Cartilage
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hartvigsen, J.; Hancock, M.J.; Kongsted, A.; Louw, Q.; Ferreira, M.L.; Genevay, S.; Hoy, D.; Karppinen, J.; Pransky, G.; Sieper, J.; et al. What low back pain is and why we need to pay attention. Lancet 2018, 391, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Manchikanti, L.; Kaye, A.D.; Abd-Elsayed, A. Treatment of Discogenic Low Back Pain: Current Treatment Strategies and Future Options—A Literature Review. Curr. Pain Headache Rep. 2019, 23, 86. [Google Scholar] [CrossRef] [PubMed]
- Kos, N.; Gradisnik, L.; Velnar, T. A Brief Review of the Degenerative Intervertebral Disc Disease. Med. Arch. 2019, 73, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, D.; Pillai, J.; Krishnamurthy, K. Genetics of intervertebral disc disease: A review. Clin. Anat. 2021, 35, 116–120. [Google Scholar] [CrossRef]
- Frapin, L.; Clouet, J.; Delplace, V.; Fusellier, M.; Guicheux, J.; Le Visage, C. Lessons learned from intervertebral disc pathophysiology to guide rational design of sequential delivery systems for therapeutic biological factors. Adv. Drug Deliv. Rev. 2019, 149–150, 49–71. [Google Scholar] [CrossRef]
- Michael, M.G.; Zakariah, S.; Samuel, R.; Dino, S. The Intervertebral Disc: Anatomy, Pathophysiology, and Aging. In Benzel’s Spine Surgery: Techniques, Complication Avoidance and Management, 5th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Raj, P.P. Intervertebral Disc: Anatomy-Physiology-Pathophysiology-Treatment. Pain Pract. 2008, 8, 18–44. [Google Scholar] [CrossRef]
- Priyadarshani, P.; Li, Y.; Yao, L. Advances in biological therapy for nucleus pulposus regeneration. Osteoarthr. Cartil. 2015, 24, 206–212. [Google Scholar] [CrossRef]
- Navone, S.E.; Marfia, G.; Giannoni, A.; Beretta, M.; Guarnaccia, L.; Gualtierotti, R.; Nicoli, D.; Rampini, P.; Campanella, R. Inflammatory mediators and signalling pathways controlling intervertebral disc degeneration. Histol. Histopathol. 2017, 32, 523–542. [Google Scholar] [CrossRef]
- Amelot, A.; Mazel, C. The Intervertebral Disc: Physiology and Pathology of a Brittle Joint. World Neurosurg. 2018, 120, 265–273. [Google Scholar] [CrossRef]
- Dowdell, J.; Erwin, M.; Choma, T.; Vaccaro, A.; Iatridis, J.; Cho, S.K. Intervertebral Disk Degeneration and Repair. Neurosurgery 2017, 80, S46. [Google Scholar] [CrossRef]
- Sharan, A.D.; Tang, S.Y.; Vaccaro, A.R. Basic Science of Spinal Disease; JP Medical Ltd.: London, UK, 2013. [Google Scholar]
- Sun, Z.; Zhao, H.; Liu, B.; Gao, Y.; Tang, W.-H.; Liu, Z.-H.; Luo, Z.-J. AF cell derived exosomes regulate endothelial cell migration and inflammation: Implications for vascularization in intervertebral disc degeneration. Life Sci. 2020, 265, 118778. [Google Scholar] [CrossRef] [PubMed]
- Bergknut, N.; Smolders, L.A.; Grinwis, G.C.; Hagman, R.; Lagerstedt, A.-S.; Hazewinkel, H.A.; Tryfonidou, M.A.; Meij, B.P. Intervertebral disc degeneration in the dog. Part 1: Anatomy and physiology of the intervertebral disc and characteristics of intervertebral disc degeneration. Veter. J. 2013, 195, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2013, 10, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Jian, X.; Sun, H.; Qin, J.; Wang, Y.; Zhang, J.; Shen, Z.; Yang, D.; Li, C.; Zhao, P.; et al. Cartilage endplate stem cells inhibit intervertebral disc degeneration by releasing exosomes to nucleus pulposus cells to activate Akt/autophagy. Stem Cells 2021, 39, 467–481. [Google Scholar] [CrossRef]
- Martins, D.E.; de Medeiros, V.P.; Wajchenberg, M.; Paredes-Gamero, E.J.; Lima, M.; Reginato, R.D.; Nader, H.B.; Puertas, E.B.; Faloppa, F. Changes in human intervertebral disc biochemical composition and bony end plates between middle and old age. PLoS ONE 2018, 13, e0203932. [Google Scholar] [CrossRef] [PubMed]
- Geiss, A.; Larsson, K.; Junevik, K.; Rydevik, B.; Olmarker, K. Autologous nucleus pulposus primes T cells to develop into interleukin-4-producing effector cells: An experimental study on the autoimmune properties of nucleus pulposus. J. Orthop. Res. 2008, 27, 97–103. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, X.-S. Pre-operative blood test for antibody to nucleus pulposus may distinguish types of lumbar disc herniation. Med. Hypotheses 2010, 75, 464–465. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, B.; Luo, Z.-J. The Immune Privilege of the Intervertebral Disc: Implications for Intervertebral Disc Degeneration Treatment. Int. J. Med. Sci. 2020, 17, 685–6925. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, B.; Liu, Z.-H.; Song, W.; Wang, D.; Chen, B.-Y.; Fan, J.; Xu, Z.; Geng, D.; Luo, Z.-J. Notochordal-Cell-Derived Exosomes Induced by Compressive Load Inhibit Angiogenesis via the miR-140-5p/Wnt/β-Catenin Axis. Mol. Ther. Nucleic Acids 2020, 22, 1092–1106. [Google Scholar] [CrossRef]
- Takada, T.; Nishida, K.; Doita, M.; Kurosaka, M. Fas Ligand Exists on Intervertebral Disc Cells: A Potential Molecular Mechanism for Immune Privilege of the Disc. Spine 2002, 27, 1526–1530. [Google Scholar] [CrossRef]
- Han, D.; Ding, Y.; Liu, S.-L.; Wang, G.; Si, I.-C.; Wang, X.; Cui, L.; Huang, D. Double role of Fas ligand in the apoptosis of intervertebral disc cells in vitro. Acta Biochim. Biophys. Sin. 2009, 41, 938–947. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sun, Z.; Wan, Z.-Y.; Guo, Y.-S.; Wang, H.-Q.; Luo, Z.-J. FasL on human nucleus pulposus cells prevents angiogenesis in the disc by inducing Fas-mediated apoptosis of vascular endothelial cells. Int. J. Clin. Exp. Pathol. 2013, 6, 2376. [Google Scholar] [CrossRef] [PubMed]
- Woods, B.I.; Vo, N.; Sowa, G.; Kang, J.D. Gene Therapy for Intervertebral Disk Degeneration. Orthop. Clin. N. Am. 2011, 42, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Molinos, M.; Almeida, C.R.; Caldeira, J.; Cunha, C.; Goncalves, R.M.; Barbosa, M.A. Inflammation in intervertebral disc degeneration and regeneration. J. R. Soc. Interface 2015, 12, 20141191. [Google Scholar] [CrossRef]
- Feng, C.; Yang, M.; Lan, M.; Liu, C.; Zhang, Y.; Huang, B.; Liu, H.; Zhou, Y. ROS: Crucial Intermediators in the Pathogenesis of Intervertebral Disc Degeneration. Oxidative Med. Cell. Longev. 2017, 2017, 5601593. [Google Scholar] [CrossRef]
- Kuo, Y.-J.; Wu, L.-C.; Sun, J.-S.; Chen, M.-H.; Sun, M.-G.; Tsuang, Y.-H. Mechanical stress-induced apoptosis of nucleus pulposus cells: An in vitro and in vivo rat model. J. Orthop. Sci. 2014, 19, 313–322. [Google Scholar] [CrossRef]
- Wang, W.-J.; Yu, X.-H.; Wang, C.; Yang, W.; He, W.-S.; Zhang, S.-J.; Yan, Y.-G.; Zhang, J. MMPs and ADAMTSs in intervertebral disc degeneration. Clin. Chim. Acta 2015, 448, 238–246. [Google Scholar] [CrossRef]
- Wang, Y.; Che, M.; Xin, J.; Zheng, Z.; Li, J.; Zhang, S. The role of IL-1β and TNF-α in intervertebral disc degeneration. BioMedicine 2020, 131, 110660. [Google Scholar] [CrossRef]
- Wuertz, K.; Haglund, L. Inflammatory Mediators in Intervertebral Disk Degeneration and Discogenic Pain. Glob. Spine J. 2013, 3, 175–184. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Y.; Li, J.; Wang, S.; Liu, Y.; Nie, L.; Cheng, L. Interleukin-9 Promotes TNF-α and PGE2 Release in Human Degenerated Intervertebral Disc Tissues. Spine 2016, 41, 1631–1640. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, M.; Chen, H.; Wu, Z.; Gao, Y.; Ma, Z.; He, X.; Kang, X. NF-κB signalling pathways in nucleus pulposus cell function and intervertebral disc degeneration. Cell Prolif. 2021, 54, e13057. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-J.; Liao, H.-Y.; Bai, D.-Y.; Wang, Z.-Q.; Xie, X.-W. MAPK /ERK signaling pathway: A potential target for the treatment of intervertebral disc degeneration. BioMedicine 2021, 143, 112170. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Liu, H.; Yang, M.; Zhang, Y.; Huang, B.; Zhou, Y. Disc cell senescence in intervertebral disc degeneration: Causes and molecular pathways. Cell Cycle 2016, 15, 1674–1684. [Google Scholar] [CrossRef]
- Zhang, G.-Z.; Deng, Y.-J.; Xie, Q.-Q.; Ren, E.-H.; Ma, Z.-J.; He, X.-G.; Gao, Y.-C.; Kang, X.-W. Sirtuins and intervertebral disc degeneration: Roles in inflammation, oxidative stress, and mitochondrial function. Clin. Chim. Acta 2020, 508, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Kalamegam, G.; Pushparaj, P.N.; Matta, C.; Memic, A.; Khademhosseini, A.; Mobasheri, R.; Poletti, F.L.; Hoyland, J.A.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Focus on articular cartilage and intervertebral disc regeneration. Methods 2016, 99, 69–80. [Google Scholar] [CrossRef]
- Kaneyama, S.; Nishida, K.; Takada, T.; Suzuki, T.; Shimomura, T.; Maeno, K.; Kurosaka, M.; Doita, M. Fas ligand expression on human nucleus pulposus cells decreases with disc degeneration processes. J. Orthop. Sci. 2008, 13, 130–135. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, T.; Tian, W.; Hu, H.; Xin, Z.; Ma, X.; Ye, C.; Hang, K.; Han, X.; Zhao, J.; et al. Loss of TIMP3 expression induces inflammation, matrix degradation, and vascular ingrowth in nucleus pulposus: A new mechanism of intervertebral disc degeneration. FASEB J. 2020, 34, 5483–5498. [Google Scholar] [CrossRef]
- Weber, K.T.; Jacobsen, T.D.; Maidhof, R.; Virojanapa, J.; Overby, C.; Bloom, O.; Quraishi, S.; Levine, M.; Chahine, N.O. Developments in intervertebral disc disease research: Pathophysiology, mechanobiology, and therapeutics. Curr. Rev. Musculoskelet. Med. 2015, 8, 18–31. [Google Scholar] [CrossRef]
- Yu, H.; Liu, Y.; Xie, W.; Xie, Q.; Liu, Q.; Cheng, L. IL-38 alleviates the inflammatory response and the degeneration of nucleus pulposus cells via inhibition of the NF-κB signaling pathway in vitro. Int. Immunopharmacol. 2020, 85, 106592. [Google Scholar] [CrossRef]
- Liu, Z.-H.; Sun, Z.; Wang, H.-Q.; Ge, J.; Jiang, T.-S.; Chen, Y.-F.; Ma, Y.; Wang, C.; Hu, S.; Samartzis, D.; et al. FasL Expression on Human Nucleus Pulposus Cells Contributes to the Immune Privilege of Intervertebral Disc by Interacting with Immunocytes. Int. J. Med. Sci. 2013, 10, 1053. [Google Scholar] [CrossRef]
- Sun, Z.; Wan, Z.-Y.; Liu, Z.-H.; Guo, Y.-S.; Yin, J.-B.; Duan, C.-G.; Gao, Y.; Li, T.; Wang, H.-Q.; Luo, Z.-J. Expression of soluble Fas and soluble FasL in human nucleus pulposus cells. Int. J. Clin. Exp. Pathol. 2013, 6, 1567. [Google Scholar] [PubMed]
- Du, X.-F.; Cui, H.-T.; Pan, H.-H.; Long, J.; Cui, H.-W.; Chen, S.-L.; Wang, J.-R.; Li, Z.-M.; Liu, H.; Huang, Y.-C.; et al. Role of the miR-133a-5p/FBXO6 axis in the regulation of intervertebral disc degeneration. J. Orthop. Transl. 2021, 29, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, Y.; Liao, Z.; Liu, H.; Zhang, S.; Zhong, D.; Qiu, X.; Chen, T.; Su, D.; Ke, X.; et al. Self-amplifying loop of NF-κB and periostin initiated by PIEZO1 accelerates mechano-induced senescence of nucleus pulposus cells and intervertebral disc degeneration. Mol. Ther. 2022, 30, 3241–3256. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Leng, P.; Song, M.; Li, D.; Guo, P.; Xu, X.; Gao, H.; Li, Z.; Li, C.; Zhang, H. Piezo1 activates the NLRP3 inflammasome in nucleus pulposus cell-mediated by Ca2+/NF-κB pathway. Int. Immunopharmacol. 2020, 85, 106681. [Google Scholar] [CrossRef]
- Zhao, K.; An, R.; Xiang, Q.; Li, G.; Wang, K.; Song, Y.; Liao, Z.; Li, S.; Hua, W.; Feng, X.; et al. Acid-sensing ion channels regulate nucleus pulposus cell inflammation and pyroptosis via the NLRP3 inflammasome in intervertebral disc degeneration. Cell Prolif. 2020, 54, e12941. [Google Scholar] [CrossRef]
- Liao, Z.; Li, S.; Liu, R.; Feng, X.; Shi, Y.; Wang, K.; Li, S.; Zhang, Y.; Wu, X.; Yang, C. Autophagic Degradation of Gasdermin D Protects against Nucleus Pulposus Cell Pyroptosis and Retards Intervertebral Disc Degeneration In Vivo. Oxidative Med. Cell. Longev. 2021, 2021, 5584447. [Google Scholar] [CrossRef]
- Navone, S.; Peroglio, M.; Guarnaccia, L.; Beretta, M.; Grad, S.; Paroni, M.; Cordiglieri, C.; Locatelli, M.; Pluderi, M.; Rampini, P.; et al. Mechanical loading of intervertebral disc modulates microglia proliferation, activation, and chemotaxis. Osteoarthr. Cartil. 2018, 26, 978–987. [Google Scholar] [CrossRef]
- Rider, S.M.; Mizuno, S.; Kang, J.D. Molecular Mechanisms of Intervertebral Disc Degeneration. Spine Surg. Relat. Res. 2019, 3, 1–11. [Google Scholar] [CrossRef]
- Capossela, S.; Schläfli, P.; Bertolo, A.; Janner, T.; Stadler, B.; Pötzel, T.; Baur, M.; Stoyanov, J. Degenerated human intervertebral discs contain autoantibodies against extracellular matrix proteins. Eur. Cells Mater. 2014, 27, 251–263. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, M.; Zhao, X.H.; Liu, Z.H.; Gao, Y.; Samartzis, D.; Wang, H.Q.; Luo, Z.J. Immune cascades in human intervertebral disc: The pros and cons. Int. J. Clin. Exp. Pathol. 2013, 6, 1009. [Google Scholar]
- Ye, F.; Lyu, F.; Wang, H.; Zheng, Z. The involvement of immune system in intervertebral disc herniation and degeneration. JOR Spine 2022, 5, e1196. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, Z.-H.; Chen, Y.-F.; Zhang, Y.-Z.; Wan, Z.-Y.; Zhang, W.-L.; Che, L.; Liu, X.; Wang, H.-Q.; Luo, Z.-J. Molecular immunotherapy might shed a light on the treatment strategies for disc degeneration and herniation. Med. Hypotheses 2013, 81, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Yao, Y.; Wang, X.; Zhang, F.; Jiang, X.; Liu, J.; Wang, H.; Li, Y.; Wang, X.; Li, H.; et al. Interleukin-21 is associated with the pathogenesis of lumbar disc herniation. Iran. J. Allergy Asthma Immunol. 2015, 509–518. [Google Scholar]
- Zhang, Y.; Liu, L.; Wang, S.; Zhao, Y.; Liu, Y.; Li, J.; Nie, L.; Cheng, L. Production of CCL20 on nucleus pulposus cells recruits IL-17-producing cells to degenerated IVD tissues in rat models. Histochem. J. 2015, 47, 81–89. [Google Scholar] [CrossRef]
- Murai, K.; Sakai, D.; Nakamura, Y.; Nakai, T.; Igarashi, T.; Seo, N.; Murakami, T.; Kobayashi, E.; Mochida, J. Primary immune system responders to nucleus pulposus cells: Evidence for immune response in disc herniation. Eur. Cell Mater. 2010, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, J.; Chen, Z.; Yang, Y.; Zhang, W.; Guo, S.; Yang, Q. Role of microRNAs in intervertebral disc degeneration (Review). Exp. Ther. Med. 2021, 22, 860. [Google Scholar] [CrossRef] [PubMed]
- Cazzanelli, P.; Wuertz-Kozak, K. MicroRNAs in Intervertebral Disc Degeneration, Apoptosis, Inflammation, and Mechanobiology. Int. J. Mol. Sci. 2020, 21, 3601. [Google Scholar] [CrossRef]
- Lin, Z.; Lu, F.; Ma, X.; Xia, X.; Zou, F.; Jiang, J. Roles of circular RNAs in the pathogenesis of intervertebral disc degeneration (Review). Exp. Ther. Med. 2021, 22, 1221. [Google Scholar] [CrossRef]
- Higashino, K.; Matsui, Y.; Yagi, S.; Takata, Y.; Goto, T.; Sakai, T.; Katoh, S.; Yasui, N. The alpha2 type IX collagen tryptophan polymorphism is associated with the severity of disc degeneration in younger patients with herniated nucleus pulposus of the lumbar spine. Int. Orthop. 2006, 31, 107–111. [Google Scholar] [CrossRef]
- Seki, S.; Kawaguchi, Y.; Mori, M.; Mio, F.; Chiba, K.; Mikami, Y.; Ikegawa, S. Association study of COL9A2 with lumbar disc disease in the Japanese population. J. Hum. Genet. 2006, 51, 1063–1067. [Google Scholar] [CrossRef]
- Annunen, S.; Paassilta, P.; Lohiniva, J.; Perälä, M.; Pihlajamaa, T.; Karppinen, J.; Tervonen, O.; Kröger, H.; Lähde, S.; Vanharanta, H.; et al. An Allele of COL9A2 Associated with Intervertebral Disc Disease. Science 1999, 285, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Pluijm, S.M.F.; Van Essen, H.W.; Bravenboer, N.; Uitterlinden, A.G.; Smit, J.H.; Pols, H.A.P.; Lips, P. Collagen type aplha I 1 Sp1 polymorphism, osteoporosis, and intervertebral disc degeneration in older men and women. Ann. Rheum. Dis. 2004, 63, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Tilkeridis, C.; Bei, T.; Garantziotis, S.; A Stratakis, C. Association of a COL1A1 polymorphism with lumbar disc disease in young military recruits. J. Med. Genet. 2005, 42, e44. [Google Scholar] [CrossRef]
- Nerlich, A.G.; Schleicher, E.D.; Boos, N. Volvo Award Winner in Basic Science Studies: Immunohistologic markers for age-related changes of human lumbar intervertebral discs. Spine 1997, 22, 2781–2795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Z.; Liu, J.; Guo, X. Advances in Susceptibility Genetics of Intervertebral Degenerative Disc Disease. Int. J. Biol. Sci. 2008, 4, 283–290. [Google Scholar] [CrossRef]
- Mio, F.; Chiba, K.; Hirose, Y.; Kawaguchi, Y.; Mikami, Y.; Oya, T.; Mori, M.; Kamata, M.; Matsumoto, M.; Ozaki, K.; et al. A Functional Polymorphism in COL11A1, Which Encodes the α1 Chain of Type XI Collagen, Is Associated with Susceptibility to Lumbar Disc Herniation. Am. J. Hum. Genet. 2007, 81, 1271–1277. [Google Scholar] [CrossRef]
- Kalb, S.; Martirosyan, N.L.; Kalani, M.Y.S.; Broc, G.G.; Theodore, N. Genetics of the Degenerated Intervertebral Disc. World Neurosurg. 2012, 77, 491–501. [Google Scholar] [CrossRef]
- Videman, T.; Gibbons, L.E.; Battié, M.C.; Maravilla, K.; Vanninen, E.; Leppävuori, J.; Kaprio, J.; Peltonen, L. The Relative Roles of Intragenic Polymorphisms of the Vitamin D Receptor Gene in Lumbar Spine Degeneration and Bone Density. Spine 2001, 26, A1–A6. [Google Scholar] [CrossRef]
- Feng, Y.; Egan, B.; Wang, J. Genetic factors in intervertebral disc degeneration. Genes Dis. 2016, 3, 178–185. [Google Scholar] [CrossRef]
- Hanaei, S.; Abdollahzade, S.; Sadr, M.; Mirbolouk, M.H.; Fattahi, E.; Khoshnevisan, A.; Rezaei, N. Association of interleukin 2, interleukin 12, and interferon-γ with intervertebral disc degeneration in Iranian population. BMC Med. Genet. 2020, 21, 143. [Google Scholar] [CrossRef]
- Hanaei, S.; Abdollahzade, S.; Sadr, M.; Mirbolouk, M.H.; Fattahi, E.; Khoshnevisan, A.; Rezaei, N. The role of interleukin 4 and IL-4RA in intervertebral disc degeneration: Investigation of single nucleotide polymorphisms in genes and a systematic review & meta-analysis of IL-4 expression level. Br. J. Neurosurg. 2019, 34, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Silva, A.J.; Pereira, P.; Vaz, R.; Gonçalves, R.M.; Barbosa, M.A. The inflammatory response in the regression of lumbar disc herniation. Arthritis Res. Ther. 2018, 20, 251. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, P.; Wu, H.; Pei, M. Meniscus, articular cartilage and nucleus pulposus: A comparative review of cartilage-like tissues in anatomy, development and function. Cell Tissue Res. 2017, 370, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Arzi, B.; DuRaine, G.; Lee, C.; Huey, D.; Borjesson, D.; Murphy, B.; Hu, J.; Baumgarth, N.; Athanasiou, K. Cartilage immunoprivilege depends on donor source and lesion location. Acta Biomater. 2015, 23, 72–81. [Google Scholar] [CrossRef]
- Jordon, J.; Konstantinou, K.; O’Dowd, J. Herniated lumbar disc. BMJ Clin. Evid. 2009. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karchevskaya, A.E.; Poluektov, Y.M.; Korolishin, V.A. Understanding Intervertebral Disc Degeneration: Background Factors and the Role of Initial Injury. Biomedicines 2023, 11, 2714. https://doi.org/10.3390/biomedicines11102714
Karchevskaya AE, Poluektov YM, Korolishin VA. Understanding Intervertebral Disc Degeneration: Background Factors and the Role of Initial Injury. Biomedicines. 2023; 11(10):2714. https://doi.org/10.3390/biomedicines11102714
Chicago/Turabian StyleKarchevskaya, Anna E., Yuri M. Poluektov, and Vasiliy A. Korolishin. 2023. "Understanding Intervertebral Disc Degeneration: Background Factors and the Role of Initial Injury" Biomedicines 11, no. 10: 2714. https://doi.org/10.3390/biomedicines11102714
APA StyleKarchevskaya, A. E., Poluektov, Y. M., & Korolishin, V. A. (2023). Understanding Intervertebral Disc Degeneration: Background Factors and the Role of Initial Injury. Biomedicines, 11(10), 2714. https://doi.org/10.3390/biomedicines11102714