
Artemisia annua L. Extracts Irreversibly Inhibit the Activity of CYP2B6 and CYP3A4 Enzymes

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Extraction
2.3. Determination of Residual Activity
2.4. Hemochromopyridine Assay
2.5. Reversible and Pseudo-Irreversible Inhibition Assay
2.6. Statistical Analysis
3. Results
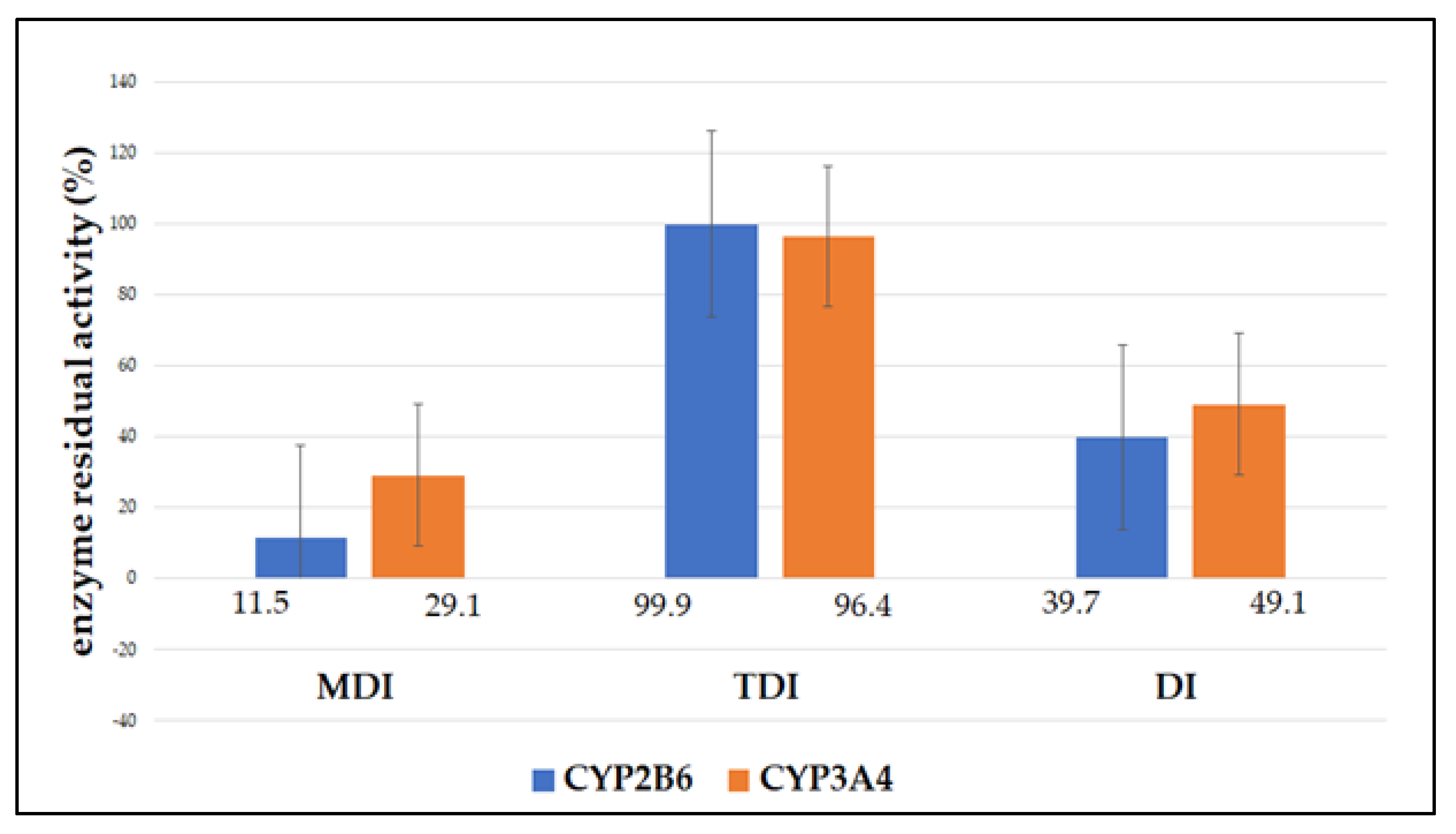
3.1. Enzyme Inhibition
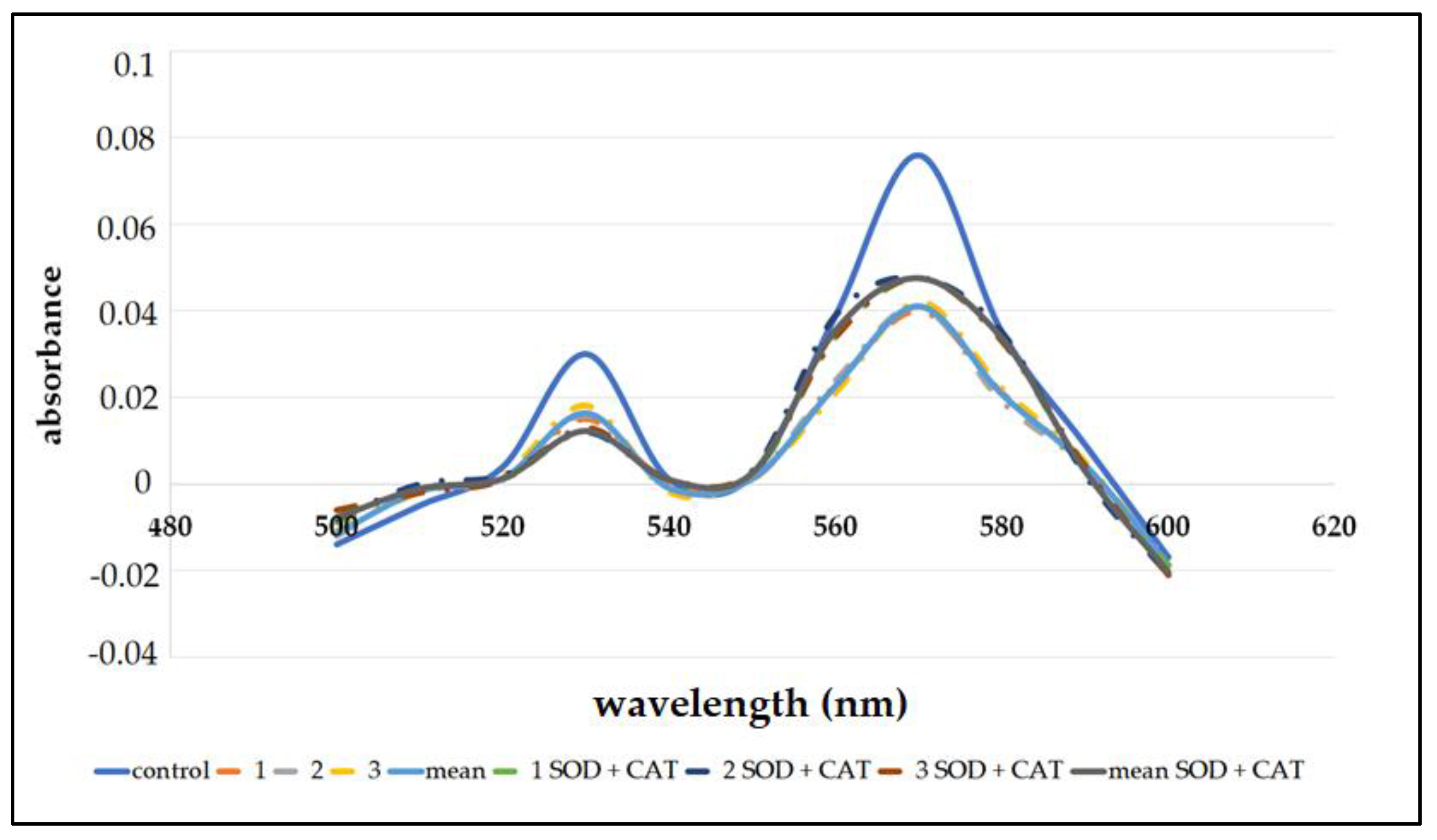
3.2. Heme Destruction
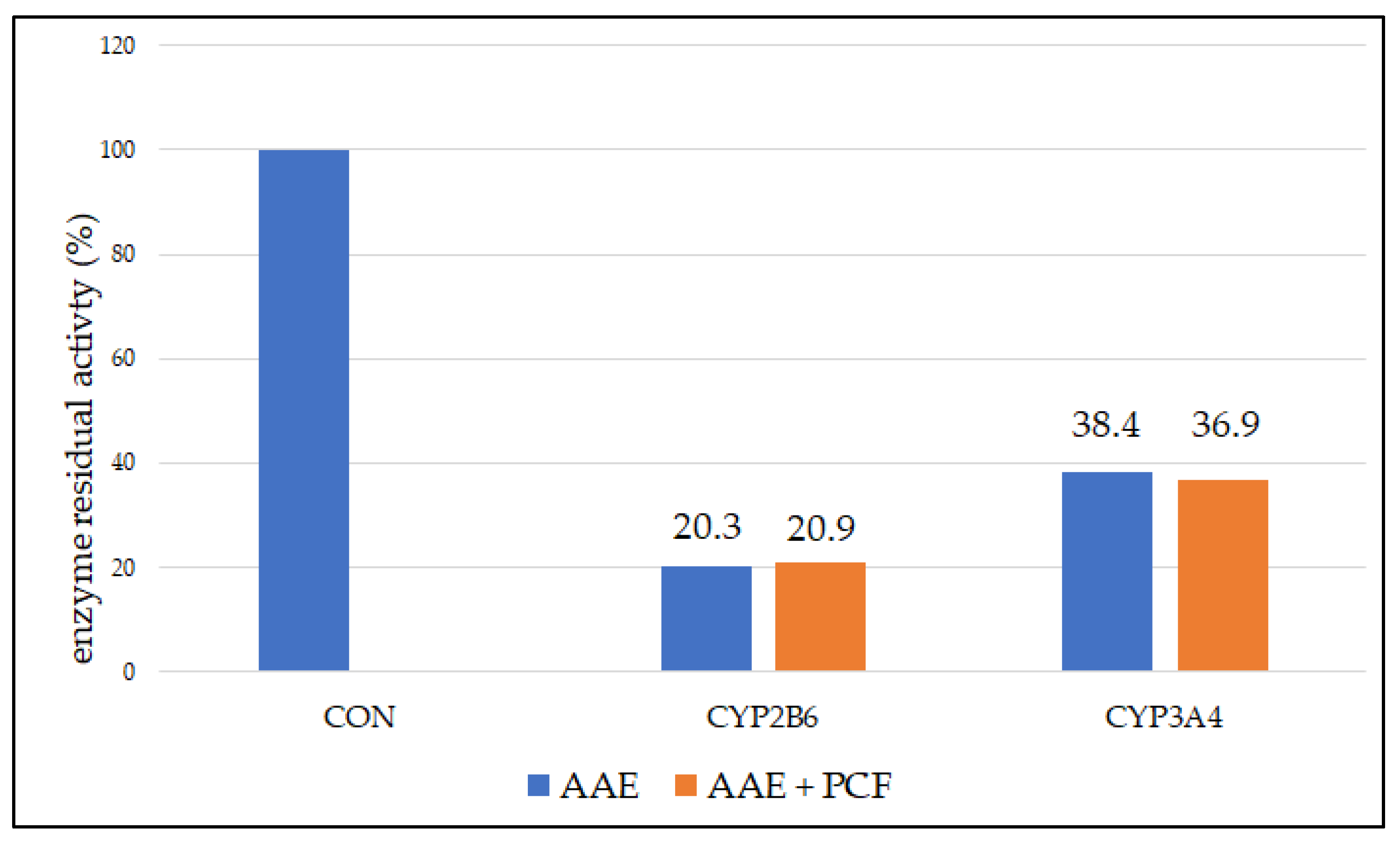
3.3. Reversable and Pseudo-Irreversible Inhibition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef] [PubMed]
- Alesaeidi, S.; Miraj, S. A Systematic Review of Anti-Malarial Properties, Immunosuppressive Properties, Anti-Inflammatory Properties, and Anti-Cancer Properties of Artemisia Annua. Electron. Phys. 2016, 8, 3150–3155. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M. Artemisia Species: From Traditional Medicines to Modern Antimalarials—And Back Again. J. Altern. Complement. Med. 2009, 15, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Store Norske Leksikon. Available online: https://snl.no/malurt (accessed on 12 November 2022).
- Mueller, M.S.; Karhagomba, I.B.; Hirt, H.M.; Wemakor, E. The Potential of Artemisia annua L. as a Locally Produced Remedy for Malaria in the Tropics: Agricultural, Chemical and Clinical Aspects. J. Ethnopharmacol. 2000, 73, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.C.; Dutta, B.; Pant, D.; Joshi, P.; Lohar, D.R. In Vitro Antibacterial Activity of Artemisia annua Linn. Growing in India. Int. J. Green Pharm. 2009, 3, 255–258. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A Review of Bioactive Essential Oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef]
- Wang, D.; Cui, L.; Chang, X.; Guan, D. Biosynthesis and Characterization of Zinc Oxide Nanoparticles from Artemisia annua and Investigate Their Effect on Proliferation, Osteogenic Differentiation and Mineralization in Human Osteoblast-like MG-63 Cells. J. Photochem. Photobiol. B 2020, 202, 111652. [Google Scholar] [CrossRef]
- Lubbe, A.; Seibert, I.; Klimkait, T.; van der Kooy, F. Ethnopharmacology in Overdrive: The Remarkable Anti-HIV Activity of Artemisia annua. J. Ethnopharmacol. 2012, 141, 854–859. [Google Scholar] [CrossRef]
- Ho, W.E.; Peh, H.Y.; Chan, T.K.; Wong, W.S.F. Artemisinins: Pharmacological Actions beyond Anti-Malarial. Pharmacol. Ther. 2014, 142, 126–139. [Google Scholar] [CrossRef]
- Kim, M.H.; Seo, J.Y.; Liu, K.H.; Kim, J.-S. Protective Effect of Artemisia annua L. Extract against Galactose-Induced Oxidative Stress in Mice. PLoS ONE 2014, 9, e101486. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Zhang, D.; Zhang, Y.; Wen, Y.; Li, L.; Zheng, L. Tumoricidal Effects of a Selenium (Se)-Polysaccharide from Ziyang Green Tea on Human Osteosarcoma U-2 OS Cells. Carbohydr. Polym. 2013, 98, 1186–1190. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Lalarizo Rakoto, M.; Marodon, C.; Bedoui, Y.; Nakab, J.; Simon, E.; Hoarau, L.; Savriama, S.; Strasberg, D.; Guiraud, P.; et al. Artemisia annua, a Traditional Plant Brought to Light. Int. J. Mol. Sci. 2020, 21, 4986. [Google Scholar] [CrossRef]
- Omura, T.; Sato, R. The Carbon Monoxide-binding Pigment of Liver Microsomes: II. Solubilization, Purification and Properties. J. Biol. Chem. 1946, 239, 2379–2385. [Google Scholar] [CrossRef]
- Hakkola, J.; Hukkanen, J.; Turpeinen, M.; Pelkonen, O. Inhibition and induction of CYP enzymes in humans: An update. Arch. Toxicol. 2020, 11, 3671–3722. [Google Scholar] [CrossRef]
- Hukkanen, J.; Pelkonen, A.; Hakkola, J.; Raunio, H. Expression and rcgulation of xcnobiotic-mctabolizing cytochromc P450 (CYP) cnzymes in human lung. Crit. Rcv. Toxicol. 2002, 32, 391–411. [Google Scholar] [CrossRef]
- Rendic, S.; Guengcrich, F.P. Survcy of Human Oxidoreductases and Cytochrome P450 Enzymes lnvolved in thc Metabolism of Xenobiotic and Natural Chcmicals. Chem. Res. Toxicol. 2015, 28, 38–42. [Google Scholar] [CrossRef]
- Lozić, M.; Rimac, H.; Bojić, M. Citokrom P450 i metabolizam lijekova—Značenje i novosti. Farm. Glasnik. 2016, 72, 747–760. [Google Scholar]
- Drug Interactions. Available online: https://drug-interactions.medicine.iu.edu/MainTable.aspx (accessed on 30 November 2022).
- Yim, D.; Kim, M.J.; Shin, Y.; Lee, S.J.; Shin, J.G.; Kim, D.H. Inhibition of Cytochrome P450 Activities by Sophora flavescens Extract and Its Prenylated Flavonoids in Human Liver Microsomes. Evid. Based. Complement. Alternat. Med. 2019, 13, 2673769. [Google Scholar] [CrossRef]
- Kondža, M.; Bojić, M.; Tomić, I.; Maleš, Ž.; Rezić, V.; Ćavar, I. Characterization of the CYP3A4 Enzyme Inhibition Potential of Selected Flavonoids. Molecules 2021, 26, 3018. [Google Scholar] [CrossRef]
- Hernandez-Maldonado, J.; Grundmann, O. Drug-Drug Interactions of Artemisinin-Based Combination Therapies in Malaria Treatment: A Narrative Review of the Literature. J. Clin. Pharmacol. 2022, 62, 1197–1205. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Martin, M.V.; Sohl, C.D.; Cheng, Q. Measurement of cytochrome P450 and NADPH-cytochrome P450 reductase. Nat. Protoc. 2009, 4, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Mashati, P.; Esmaeili, S.; Dehghan-Nayeri, N.; Bashash, D.; Darvishi, M.; Gharehbaghian, A. Methanolic Extract from Aerial Parts of Artemisia annua L. Induces Cytotoxicity and Enhances Vincristine-Induced Anticancer Effect in Pre-B Acute Lymphoblastic Leukemia Cells. Int. J. Hematol. Oncol. Stem. Cell. Res. 2019, 13, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.R.; Cheng, Y.S.; Ding, D.Y. Determination of mephenytoin and 4’-hydroxymephenytoin in urine by high performance liquid chromatography. Yao Xue Xue Bao 1994, 29, 624–628. [Google Scholar] [PubMed]
- Flink, E.B.; Watson, C.J. A Method for the Quantitative Determination of Hemoglonin and Related Heme Pigments in Feces, Urine, and Blood Plasa. J. Biol. Chem. 1942, 146, 171–178. [Google Scholar] [CrossRef]
- Paul, K.G.; Theorell, H.; Åkeson, Å.; Virtanen, A.I.; Sörensen, N.A. The Molar Light Absorption of Pyridine Ferroprotoporphrin (Pyridine Haemochromogen). Acta Chem. Scand. 1953, 7, 1284–1287. [Google Scholar] [CrossRef]
- Bojić, M.; Barbero, L.; Dolgos, H.; Freisleben, A.; Gallemann, D.; Riva, S.; Guengerich, F.P. Time- and NADPH-dependent Inhibition of Cytochrome P450 3A4 by the Cyclopentapeptide Cilengitide: Significance of the Guanidine Group and Accompanying Spectral Changes. Drug Metab. Dispos. 2014, 42, 1438–1446. [Google Scholar] [CrossRef]
- Zhou, S.; Gao, Y.; Jiang, W.; Huang, M.; Xu, A.; Paxton, J.W. Interactions of herbs with cytochrome P450. Drug. Metab. Rev. 2003, 35, 35–98. [Google Scholar] [CrossRef]
- Usia, T.; Watabe, T.; Kadota, S.; Tezuka, Y. Mechanism-based inhibition of CYP3A4 by constituents of Zingiber aromaticum. Biol. Pharm. Bull. 2005, 28, 495–499. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Aburatani, M.; Yoshida, T.; Yamashita, Y.; El-Beih, A.A.; Ohta, T. CYP3A4 inhibitors isolated from Licorice. Biol. Pharm. Bull. 2005, 28, 2000–2002. [Google Scholar] [CrossRef]
- Melillo de Magalhães, P.; Dupont, I.; Hendrickx, A.; Joly, A.; Raas, T.; Dessy, S.; Sergent, T.; Schneider, Y.J. Anti-inflammatory effect and modulation of cytochrome P450 activities by Artemisia annua tea infusions in human intestinal Caco-2 cells. Food. Chem. 2012, 134, 864–871. [Google Scholar] [CrossRef]
- Špičáková, A.; Bazgier, V.; Skálová, L.; Otyepka, M.; Anzenbacher, P. Beta-caryophyllene oxide and trans-nerolidol affect enzyme activity of CYP3A4—In vitro and in silico studies. Physiol. Res. 2019, 68, S51–S58. [Google Scholar] [CrossRef]
- Šarić Mustapić, D.; Debeljak, Ž.; Maleš, Ž.; Bojić, M. The Inhibitory Effect of Flavonoid Aglycones on the Metabolic Activity of CYP3A4 Enzyme. Molecules 2018, 23, 2553. [Google Scholar] [CrossRef]
- Ueng, Y.F.; Chen, C.C.; Yamazaki, H.; Kiyotani, K.; Chang, Y.P.; Lo, W.S.; Li, D.T.; Tsai, P.L. Mechanism-based inhibition of CYP1A1 and CYP3A4 by the furanocoumarin chalepensin. Drug. Metab. Pharmacokinet. 2013, 28, 229–238. [Google Scholar] [CrossRef]
- National Library of Medicine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Luteolin (accessed on 1 December 2022).
- Desrosiers, M.R.; Mittelman, A.; Weathers, P.J. Dried Leaf Artemisia annua Improves Bioavailability of Artemisinin via Cytochrome P450 Inhibition and Enhances Artemisinin Efficacy Downstream. Biomolecules 2020, 7, 254. [Google Scholar] [CrossRef]
- Fu, C.; Zhang, K.; Wang, M.; Qiu, F. Multi-component pharmacokinetics assessment of Artemisia annua L. in rats based on LC-ESI-MS/MS quantification combined with molecular docking. Arab. J. Chem. 2022, 15, 104254. [Google Scholar] [CrossRef]
- Bojić, M. Pretklinička ispitivanja inhibicijskog i interakcijskog potencijala novih lijekova na razini citokroma P450. Farm. Glasnik. 2015, 71, 229–242. [Google Scholar]
- Tang, L.W.T.; Verma, R.K.; Fan, H.; Chan, E.C.Y. Mechanism-Based Inactivation of Cytochrome P450 3A4 by Benzbromarone. Mol. Pharmacol. 2021, 99, 266–276. [Google Scholar] [CrossRef]
- Lee, M.H.; Graham, G.G.; Williams, K.M.; Day, R.O. A benefit-risk assessment of benzbromarone in the treatment of gout. Was its withdrawal from the market in the best interest of patients? Drug. Saf. 2008, 31, 643–665. [Google Scholar] [CrossRef]
- Sevrioukova, I.F.; Poulos, T.L. Structure and mechanism of the complex between cytochrome P4503A4 and ritonavir. Proc. Natl. Acad. Sci. USA 2010, 107, 18422–18427. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monoterpenes | Sesquiterpenes | Phenolic Compounds | Coumarins |
|---|---|---|---|
| 1,8-cineole | artemisinin | quinic acid | scopolin |
| α-and-β-pinene | arteannuin B | caffeic acid | scopoletin |
| camphene | artemisinic acid | luteolin | |
| borneol | quercetin | ||
| camphor | rutin | ||
| carvone | apigenin | ||
| limonene | isorhamnetin | ||
| α-terpinene | kaempferol | ||
| myrtenol | mearnsetin | ||
| artemetin | |||
| eupatorine |
| CYP2B6 | |||
| Incubation | MDI | TDI | DI |
| 1 | 11.1 | 99.8 | 39.9 |
| 2 | 12.1 | 101.2 | 40.4 |
| 3 | 11.5 | 98.9 | 38.9 |
| Mean | 11.5 ± 0.5 | 99.9 ± 1.1 | 39.7 ± 0.7 |
| statistical significance | p < 0.05 | p > 0.05 | p < 0.05 |
| CYP3A4 | |||
| 1 | 29.9 | 95.4 | 48.8 |
| 2 | 29.1 | 96.7 | 48.2 |
| 3 | 28.2 | 97.1 | 50.2 |
| Mean | 29.1 ± 0.8 | 96.4 ± 0.8 | 49.1 ± 0.9 |
| statistical significance | p < 0.05 | p > 0.05 | p < 0.05 |
| Incubation | Heme Concentration (%) | |
|---|---|---|
| Without SOD and CAT | With SOD and CAT | |
| 1 | 52.9 | 62.5 |
| 2 | 53.2 | 61.9 |
| 3 | 53.5 | 60.9 |
| Mean | 53.2 ± 0.3 | 61.8 ± 0.3 |
| statistical significance | p < 0.05 | p < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondža, M.; Mandić, M.; Ivančić, I.; Vladimir-Knežević, S.; Brizić, I. Artemisia annua L. Extracts Irreversibly Inhibit the Activity of CYP2B6 and CYP3A4 Enzymes. Biomedicines 2023, 11, 232. https://doi.org/10.3390/biomedicines11010232
Kondža M, Mandić M, Ivančić I, Vladimir-Knežević S, Brizić I. Artemisia annua L. Extracts Irreversibly Inhibit the Activity of CYP2B6 and CYP3A4 Enzymes. Biomedicines. 2023; 11(1):232. https://doi.org/10.3390/biomedicines11010232
Chicago/Turabian StyleKondža, Martin, Marta Mandić, Ivona Ivančić, Sanda Vladimir-Knežević, and Ivica Brizić. 2023. "Artemisia annua L. Extracts Irreversibly Inhibit the Activity of CYP2B6 and CYP3A4 Enzymes" Biomedicines 11, no. 1: 232. https://doi.org/10.3390/biomedicines11010232
APA StyleKondža, M., Mandić, M., Ivančić, I., Vladimir-Knežević, S., & Brizić, I. (2023). Artemisia annua L. Extracts Irreversibly Inhibit the Activity of CYP2B6 and CYP3A4 Enzymes. Biomedicines, 11(1), 232. https://doi.org/10.3390/biomedicines11010232