


Proteomic Approaches in the Study of Placenta of Pregnancy Complicated by Gestational Diabetes Mellitus


Abstract
:1. Introduction
2. Placenta Protein Profile
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Desoye, G.; Wells, J.C.K. Pregnancies in Diabetes and Obesity: The Capacity-Load Model of Placental Adaptation. Diabetes 2021, 70, 823–830. [Google Scholar] [CrossRef] [PubMed]
- International Association of Diabetes and Pregnancy Study Groups Consensus Panel. International Association of Diabetes and Pregnancy Study Groups: Recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Koning, S.H.; Hoogenberg, K.; Lutgers, H.L.; van den Berg, P.P.; Wolffenbuttel, B.H. Gestational Diabetes Mellitus: Current knowledge and unmet needs. J. Diabetes 2016, 8, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Burlina, S.; Dalfrà, M.G.; Lapolla, A. Short- and long-term consequences for offspring exposed to maternal diabetes: A review. J. Matern.-Fetal Neonatal Med. 2019, 32, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Burlina, S.; Dalfrà, M.G.; Lapolla, A. Long-term cardio-metabolic effects after gestational diabetes: A review. J. Matern.-Fetal Neonatal Med. 2021, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kampmann, U.; Knorr, S.; Fuglsang, J.; Ovesen, P. Determinants of Maternal Insulin Resistance during Pregnancy: An Updated Overview. J. Diabetes Res. 2019, 19, 5320156. [Google Scholar] [CrossRef]
- Lapolla, A.; Dalfrà, M.G.; Mello, G.; Parretti, E.; Cioni, R.; Marzari, C.; Masin, M.; Ognibene, A.; Messeri, G.; Fedele, D.; et al. Early detection of insulin sensitivity and beta-cell function with simple tests indicates future derangements in late pregnancy. J. Clin. Endocrinol. Metab. 2008, 93, 876–880. [Google Scholar] [CrossRef]
- Dalfrà, M.G.; Burlina, S.; Del Vescovo, G.G.; Lapolla, A. Genetics and Epigenetics: New Insight on Gestational Diabetes Mellitus. Front. Endocrinol. 2020, 11, 602477. [Google Scholar] [CrossRef]
- Hauguel-de Mouzon, S.; Desoye, G. The placenta in diabetic pregnancy: New methodological approaches. In Gestational Diabetes: A Decade after the HAPO Study; Lapolla, A., Metzger, B.E., Eds.; Frontier in Diabetes, Karger: Basel, Switzerland, 2020; pp. 145–154. [Google Scholar]
- Florens, L.; Washburn, M.P. Proteomic analysis by multidimensional protein identification technology. Methods Mol. Biol. 2006, 328, 159–175. [Google Scholar]
- Navajas, R.; Corrales, F.; Paradela, A. Quantitative proteomics-based analyses performed on pre-eclampsia samples in the 2004–2020 period: A systematic review. Clin. Proteom. 2021, 18, 3–13. [Google Scholar] [CrossRef]
- Robinson, J.M.; Ackerman, W.E., 4th; Kniss, D.A.; Takizawa, T.; Vandré, D.D. Proteomics of the human placenta: Promises and realities. Placenta 2008, 29, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapolla, A.; Porcu, S.; Roverso, M.; Desoye, G.; Cosma, C.; Nardelli, G.B.; Bogana, G.; Carozzini, M.; Traldi, P. A preliminary investigation on placenta protein profile reveals only modest changes in well controlled gestational diabetes mellitus. Eur. J. Mass Spectrom. 2013, 19, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Hillenkamp, F.; Karas, M.; Beavis, R.C.; Chait, B.T. Matrix-assisted laser desorption/ionization mass spectrometry of biopolymers. Anal Chem. 1991, 63, 1193A–1203A. [Google Scholar] [CrossRef] [PubMed]
- Francese, S.; Dani, F.R.; Traldi, P.; Mastrobuoni, G.; Pieraccini, G.; Moneti, G. MALDI mass spectrometry imaging, from its origins up to today: The state of the art. Comb. Chem. High Throughput Screen. 2009, 12, 156–174. [Google Scholar] [CrossRef]
- Roverso, M.; Lapolla, A.; Cosma, C.; Seraglia, R.; Galvan, E.; Visentin, S.; Cosmi, E.; Desoye, G.; Traldi, P. Some preliminary matrix-assisted laser desorption/ionization imaging experiments on maternal and fetal sides of human placenta. Eur. J. Mass Spectrom. 2014, 20, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Cristoni, S.; Molin, L.; Crotti, S.; Rossi Bernardi, L.; Seraglia, R.; Traldi, P. Sieve-based device for MALDI sample preparation. II. Instrumental parameterization. J. Mass Spectrom. 2009, 44, 1579–1586. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, L.; Xu, Z.; Liang, G. Identification and analysis of multi-protein complexes in placenta. PLoS ONE 2013, 8, e62988. [Google Scholar] [CrossRef]
- Roverso, M.; Brioschi, M.; Banfi, C.; Visentin, S.; Burlina, S.; Seraglia, R.; Traldi, P.; Lapolla, A. A preliminary study on human placental tissue impaired by gestational diabetes: A comparison of gel-based versus gel-free proteomics approaches. Eur. J. Mass Spectrom. 2016, 22, 71–82. [Google Scholar] [CrossRef]
- Burlina, S.; Banfi, C.; Brioschi, M.; Visentin, S.; Dalfrà, M.G.; Traldi, P.; Lapolla, A. Is the placental proteome impaired in well-controlled gestational diabetes? J. Mass Spectrom. 2019, 54, 359–365. [Google Scholar] [CrossRef]
- Blois, S.M.; Gueuvoghlanian-Silva, B.Y.; Tirado-González, I.; Torloni, M.R.; Freitag, N.; Mattar, R.; Conrad, M.L.; Unverdorben, L.; Barrientos, G.; Knabl, J.; et al. Getting too sweet: Galectin-1 dysregulation in gestational diabetes mellitus. Mol. Hum. Reprod. 2014, 20, 644–649. [Google Scholar] [CrossRef]
- Favaro, R.R.; Salgado, R.M.; Covarrubias, A.C.; Bruni, F.; Lima, C.; Fortes, Z.B.; Zorn, T.M. Long-term type 1 diabetes impairs decidualization and extracellular matrix remodeling during early embryonic development in mice. Placenta 2013, 34, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Abdulsid, A.; Fletcher, A.; Lyall, F. Heat shock protein 27 is spatially distributed in the human placenta and decreased during labor. PLoS ONE 2013, 8, e71127. [Google Scholar]
- Liu, B.; Xu, Y.; Voss, C.; Qiu, F.H.; Zhao, M.Z.; Liu, Y.D.; Nie, J.; Wang, Z.L. Altered protein expression in gestational diabetes mellitus placentas provides insight into insulin resistance and coagulation/fibrinolysis pathways. PLoS ONE 2012, 7, e44701. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Qu, T.; He, X.; Yang, X.; Guo, N.; Mao, Y.; Xu, X.; Sun, X.; Zhang, X.; Wang, W. Screening of differentially expressed proteins from syncytiotrophoblast for severe early-onset preeclampsia in women with gestational diabetes mellitus using tandem mass tag quantitative proteomics. BMC Pregnancy Childbirth 2018, 18, 437. [Google Scholar] [CrossRef]
- Nirupama, R.; Divyashree, S.; Janhavi, P.; Muthukumar, S.P.; Ravindra, P.V. Preeclampsia: Pathophysiology and management. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 101975. [Google Scholar] [CrossRef]
- Szabo, S.; Karaszi, K.; Romero, R.; Toth, E.; Szilagyi, A.; Gelencser, Z.; Xu, Y.; Balogh, A.; Szalai, G.; Hupuczi, P.; et al. Proteomic identification of Placental Protein 1 (PP1), PP8, and PP22 and characterization of their placental expression in healthy pregnancies and in preeclampsia. Placenta 2020, 99, 197–207. [Google Scholar] [CrossRef]
- Assi, E.; D’Addio, F.; Mandò, C.; Maestroni, A.; Loretelli, C.; Ben Nasr, M.; Usuelli, V.; Abdelsalam, A.; Seelam, A.J.; Pastore, I.; et al. Placental proteome abnormalities in women with gestational diabetes and large-for-gestational-age newborns. BMJ Open Diabetes Res. Care 2020, 8, e001586. [Google Scholar] [CrossRef]
- Pettitt, D.J.; Nelson, R.G.; Saad, M.F.; Bennett, P.H.; Knowler, W.C. Diabetes and obesity in the offspring of Pima Indian women with diabetes during pregnancy. Diabetes Care 1993, 16, 310–314. [Google Scholar] [CrossRef]
- Jansson, N.; Greenwood, S.L.; Johansson, B.R.; Powell, T.L.; Jansson, T. Leptin stimulates the activity of the system A amino acid transporter in human placental villous fragments. J. Clin. Endocrinol. Metab. 2003, 88, 1205–1211. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Lee, J.E.; Moon, P.G.; Lee, I.K.; Baek, M.C. Proteomic Analysis of Extracellular Vesicles Released by Adipocytes of Otsuka Long-Evans Tokushima Fatty (OLETF) Rats. Protein J. 2015, 34, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Jayabalan, N.; Lai, A.; Ormazabal, V.; Adam, S.; Guanzon, D.; Palma, C.; Scholz-Romero, K.; Lim, R.; Jansson, T.; McIntyre, H.D.; et al. Adipose Tissue Exosomal Proteomic Profile Reveals a Role on Placenta Glucose Metabolism in Gestational Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2019, 104, 1735–1752. [Google Scholar] [CrossRef] [PubMed]
- Barrozo, E.; Aagaard, K.M. Human placental biology at single-cell resolution: A contemporaneous review. BJOG 2022, 129, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, Q.; Liu, Y.; Garmire, L.X. Single cell transcriptome research in human placenta. Reproduction 2020, 160, R155–R167. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, F.; Peng, Y.; Chen, R.; Zhou, W.; Wang, H.; Ouyang, J.; Yu, B.; Xu, Z. Transcriptomic Profiling of Human Placenta in Gestational Diabetes Mellitus at the Single-Cell Level. Front. Endocrinol. 2021, 7, 679582. [Google Scholar] [CrossRef] [PubMed]
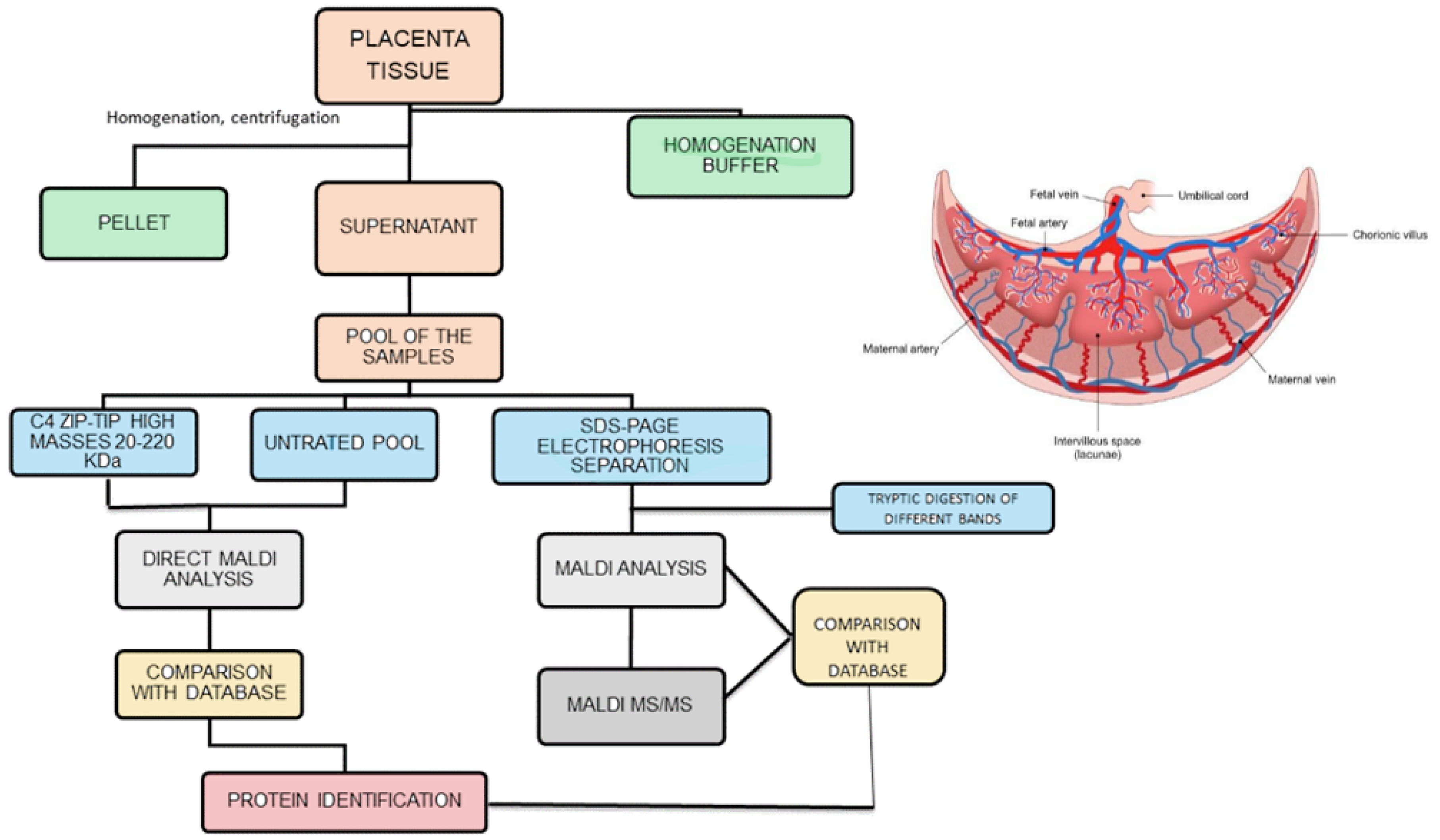

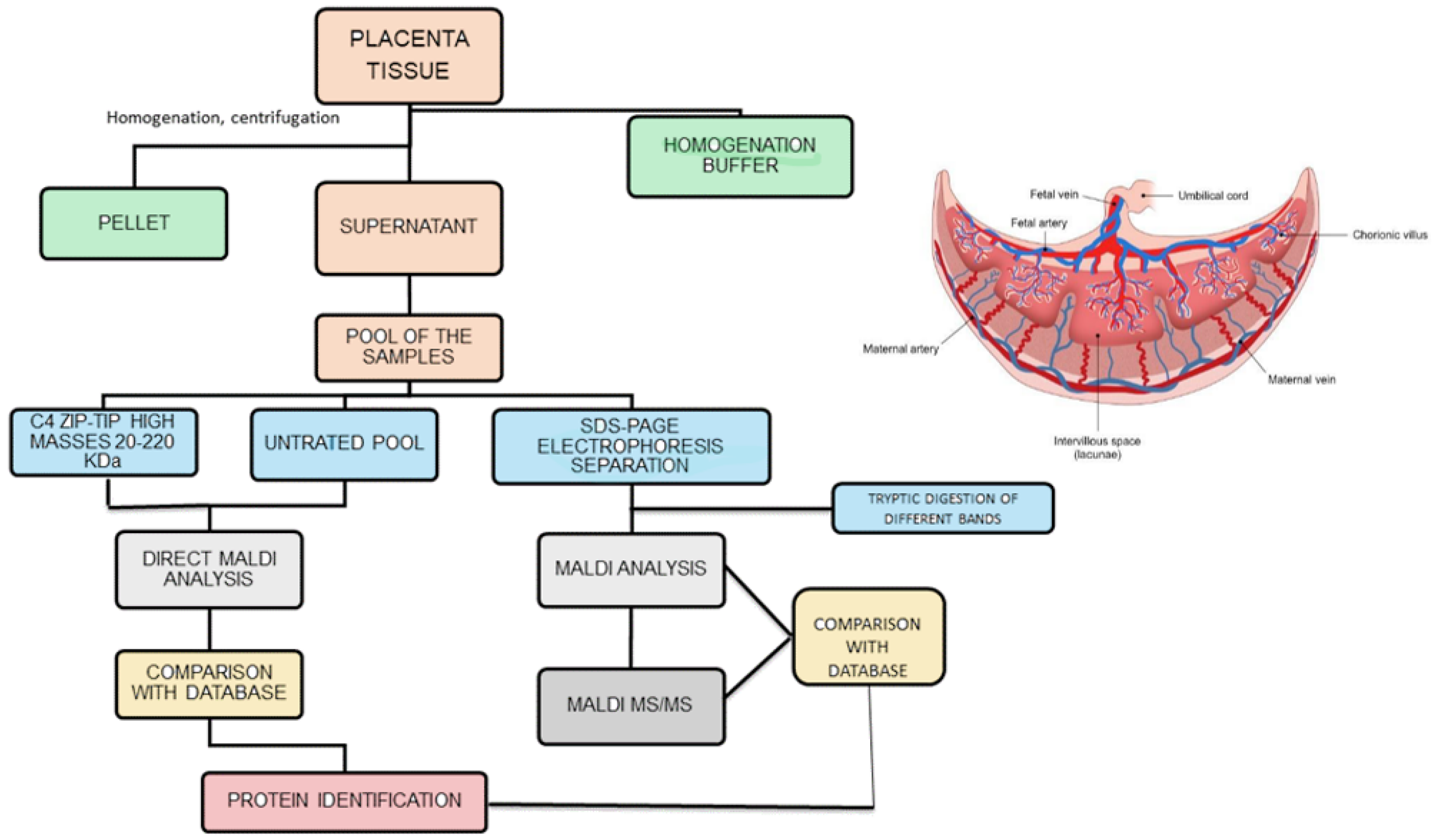{kind=link}
| PROTEIN DETECTED | NCBI | PROTEIN ROLE | REF. |
|---|---|---|---|
| Serine/arginine repetitive matrix protein 1 | SRRM1 | Transcription factor involved in pre mRNA proccessing | [13] |
| NADH dehydrogenase iron-sulfur protein 3 mitochondrial | NDUFS3 | Core subunit of mitochondrial complex 1 | [16] |
| Transcription cofactor vestigial-like protein 4 | Vgll4 | Nuclear transcription factor | [16] |
| Voltage dependent anion-selective channel protein 2 | VDAC2 | Channel through mitochondrial membrane allows diffusion of small molecules | [16] |
| Galectin-1 | Gal-1 | Regulator T-cell homeostasis and survival, T-cell immune disorders, inflammation | [19,20,21] |
| Bcl2-associated transcriptor factor1 | BCLAF1 | Transcriptional repressor, its over expression induces apoptosis | [13] |
| Collagen alpha 2 VI chain | COL6A2 | Human collagen | [19] |
| Collagen alpha-1 | COLA1 | Extracellular matrix collagen | [20,21] |
| Actin related protein 2 3 complex subunit 2 | ARPCS | Protein involved in cell migration and invasion | [19] |
| Phosphatidylcholine transfer protein | PCTP | Member of the steroidogenic acute regulatory protein-related transfer superfamily. | [19] |
| Voltage dependent anion selective channel protein 2 2 | VDAC22 | Transporter of ions and metabolites across the mitochondrial membrane | [19] |
| Periostin | POSTN | Matricellular protein involved in regulation of cell adhesion, cell differentiation, and organization of extracellular matrix. | [19] |
| Ig gamma-2 chain C region | IGHG2 | Constant region of immunoglobulin heavy chain | [19] |
| Moesin | MSN | Cytoskeletal adaptor protein that plays an important role in modification of the actin cytoskeleton. | [19] |
| Triosephosphate isomerase | TPI | Enzyme that catalyzes the reversible interconversion of the triose phosphate isomers dihydroxyacetone phosphate and D-glyceraldehyde 3-phosphate. | [16,19] |
| Protein disulfide-isomerase | PDI | Enzymes that catalyze exchange reactions between -S-H and S-S- groups, the formation or breaking of sulfur bridges | [19] |
| Vimentin | VIM | Filament proteinresponsible for maintaining cell shape, integrity of the cytoplasm, and stabilizing cytoskeletal interactions | [19] |
| 14-3-3 protein beta/alpha | GB | Key regulators of processes of mitosis and apoptosis. | [19] |
| Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2 | GNB2 | Transducer | [24] |
| 26S proteasome non-ATPase regulatory subunit 14 | PSDE | Hydrolase Metalloprotease Protease | [24] |
| Annexin A5 | A5 | Blood coagulation Signal transduction | [24] |
| Annexin A4 | A4 | Anti-apoptosis Signal transduction | [24] |
| 14-3-3 protein zeta/delta | YWHAZ | Anti-apoptosis Signal transduction | [24] |
| Glycyl-tRNA synthetase | GARS | ATP binding | [24] |
| Myosin regulatory light polypeptide 9 | MYL 9 | Calcium ion binding | [24] |
| Annexin A2 | A2 | Calcium-dependent phospholipid binding | [24] |
| Fumarate hydratase mitochondrial | FHM | Fumarate hydratase activity | [24] |
| Laminin subunit gamma 3 | LAMC3 | Extracellular marix protein | [19] |
| L lactate deydrogenase B chain | LDHB | Dehydrogenase protein | [19] |
| Pyridoxal kinase | PLK | Kinase involved in vitamin B metabolism | [19] |
| Proteasome subunit beta type 4 | PSMB4 | Subunit of proteasome involved in cellular proteolisis | [19] |
| Complement C4A | CG4 | Component of human complement | [19] |
| Trifuncional enzyme subunit alpha mitocondrial | TP | Catalizes the last three of the four reactions of the mitochondrial beta-oxidation pathway | [19] |
| Calcium binding mitochondrial carrier protein Aralar 2 | CMC2 | Catalyzes the calcium dependent exchange of cytoplasmic glutamate with mitochondrial aspartate across the mitochondrial membrane | [19] |
| Csh-like fibrillar surface protein | CSH | Fibrillar surface protein | [19] |
| Bone marrow proteoglycan | PGR2 | Placenta regulation of peptide hormone and growth factor activity | [28] |
| Amiloride-sensitive amine oxidase | DAO2 | Control of histamine levels | [28] |
| Glia-derived nexin | SERPINE 2 | Placental remodelling | [28] |
| Complement factor H-related protein 1 | CFHR1 | Complement regulation and lipid metabolism | [28] |
| Dypeptidyl peptidase 4 | DPP49 | Regulation of fetal insulin levels and beta-cell development | [28] |
| Fibronectin | FN1 | Placental inflammation | [28] |
| PROTEIN DETECTED | NCBI | PROTEIN ROLE | REF. |
|---|---|---|---|
| Heath Shock protein | HSP70 | Stress-induced protein | [20] |
| Collagen alpha-1 | COLA1 | Extracellular matrix collagen | [20,21] |
| Alpha-1-antitripsin | A1AT | Protease inhibitor | [19] |
| Alpha-2-antiplasmin | ά2AP | Serine protease inhibitor of plasmine | [19] |
| ApolipoproteinA-I | APOA-1 | Protein component of liporotein | [19] |
| Clusterin | CLU | Protein associated with the clearance of cellular debris and apoptosis. | [19] |
| Fibrinogen alpha chain | FGA | Alpha chain of fibrinogen, involved in platelet activation | [19,24] |
| Fibrinogen beta chain | FGB | Beta chain of fibrinogen, involved in platelet activation | [19,24] |
| Fibrinogen gamma chain | FGG | Gamma chain of fibrinogen, involved in platelet activation | [19] |
| Ig gamma-1 chain C region | Ighg1 | Constant region of immunoglobulin heavy chains. | [19] |
| Vitronectin | VN | cell adhesion proteins in plasma. | [19] |
| Ras-related protein Rap-1A | RAP1A | Signal transduction | [24] |
| Tryptophanyl-tRNA synthetase citoplasmic | WRS | Triptophan-Trna ligase activity | [24] |
| NADH-cytocrome b5 reductase 3 | CYB5R3 | Lipid metabolism | [28] |
| Corticosteroid 11-beta-dehydrogenase isozyme 2 | HSD11B2 | Regulation of feto-maternal cortisol levels | [28] |
| Caspase-1 | CASP1 | Activation of placental inflammasome | [28] |
| N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 | DDAH1 | ADMA_NO pathway homeostasis during pregnancy | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapolla, A.; Traldi, P. Proteomic Approaches in the Study of Placenta of Pregnancy Complicated by Gestational Diabetes Mellitus. Biomedicines 2022, 10, 2272. https://doi.org/10.3390/biomedicines10092272
Lapolla A, Traldi P. Proteomic Approaches in the Study of Placenta of Pregnancy Complicated by Gestational Diabetes Mellitus. Biomedicines. 2022; 10(9):2272. https://doi.org/10.3390/biomedicines10092272
Chicago/Turabian StyleLapolla, Annunziata, and Pietro Traldi. 2022. "Proteomic Approaches in the Study of Placenta of Pregnancy Complicated by Gestational Diabetes Mellitus" Biomedicines 10, no. 9: 2272. https://doi.org/10.3390/biomedicines10092272
APA StyleLapolla, A., & Traldi, P. (2022). Proteomic Approaches in the Study of Placenta of Pregnancy Complicated by Gestational Diabetes Mellitus. Biomedicines, 10(9), 2272. https://doi.org/10.3390/biomedicines10092272