
Hydroxycinnamic Acid Derivatives from Coffee Extracts Prevent Amyloid Transformation of Alpha-Synuclein
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Coffee Extracts
2.2. Purification of Recombinant Alpha-Synuclein
2.3. Fibrillation of Alpha-Synuclein
2.4. Thioflavin T Fluorescence Analysis
2.5. Determination of the Half-Maximum Inhibitory Concentration (IC50)
2.6. Determination of Inhibitory Constant (Ki) for Coffee Extracts
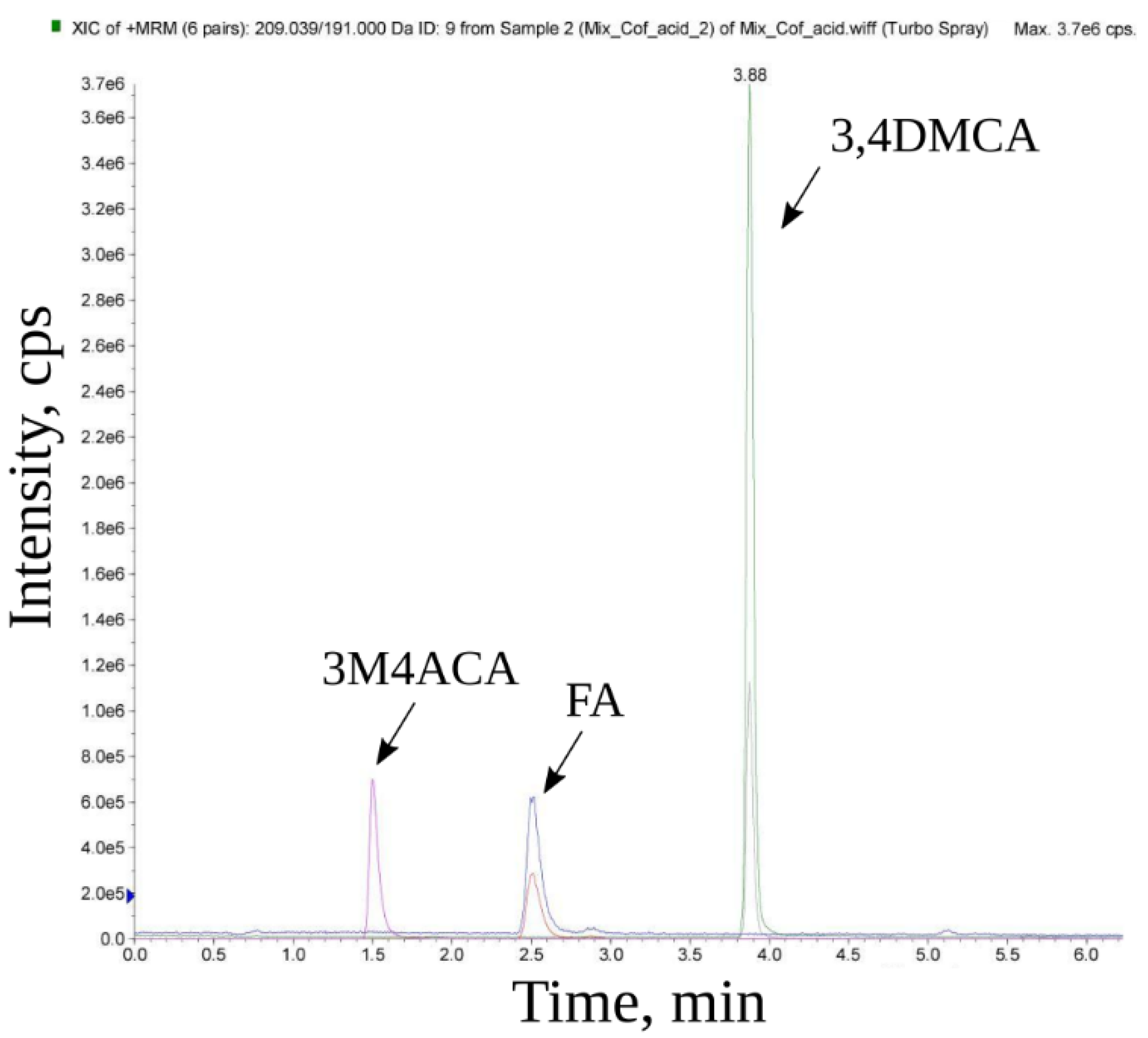
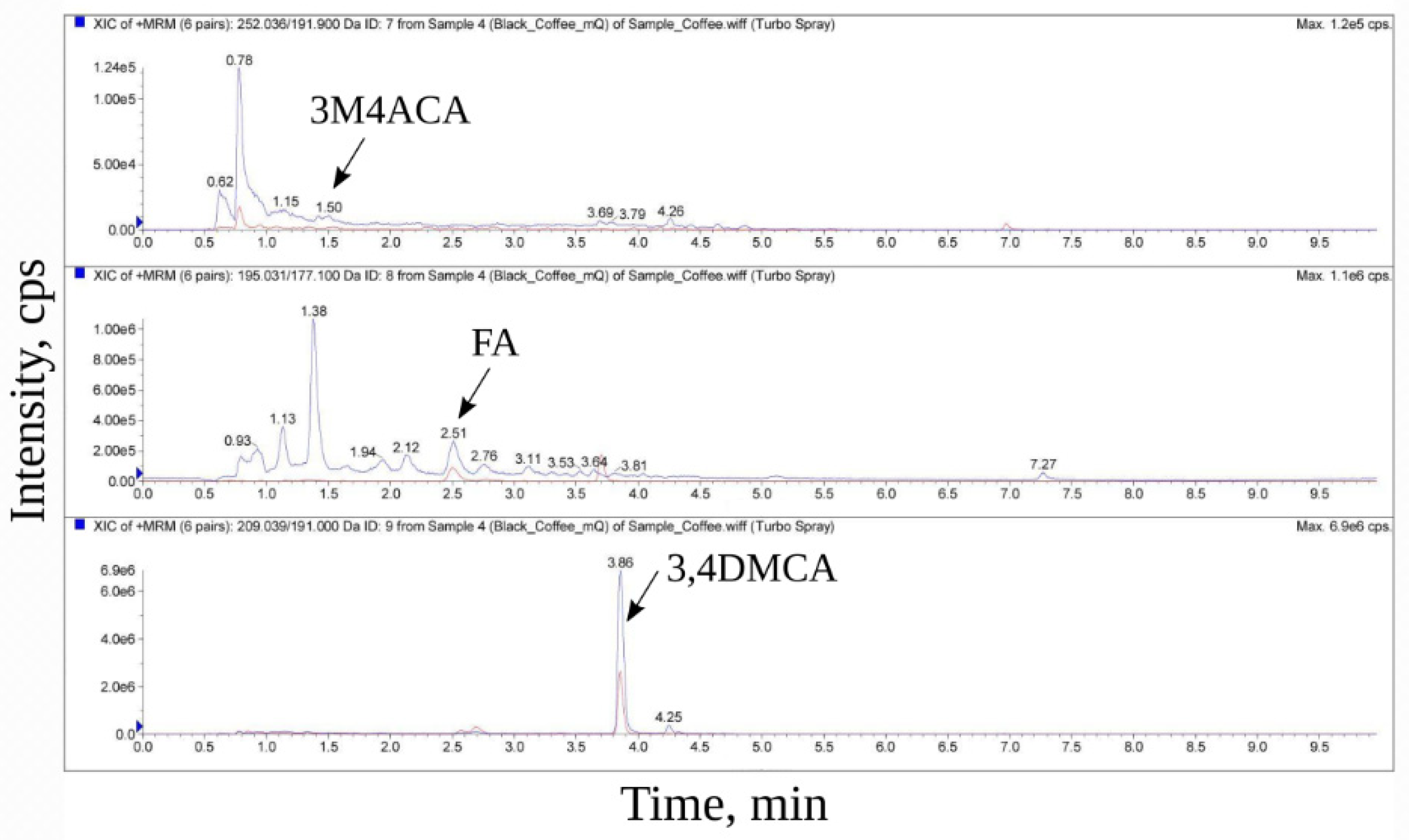
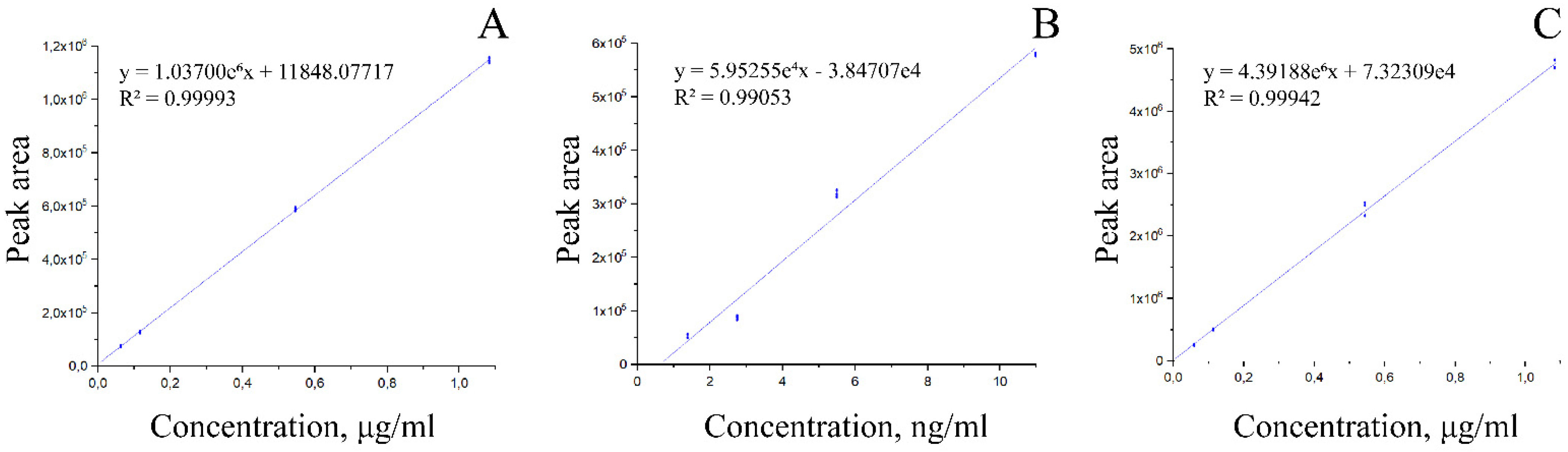
2.7. Mass Spectroscopic Analysis
3. Results
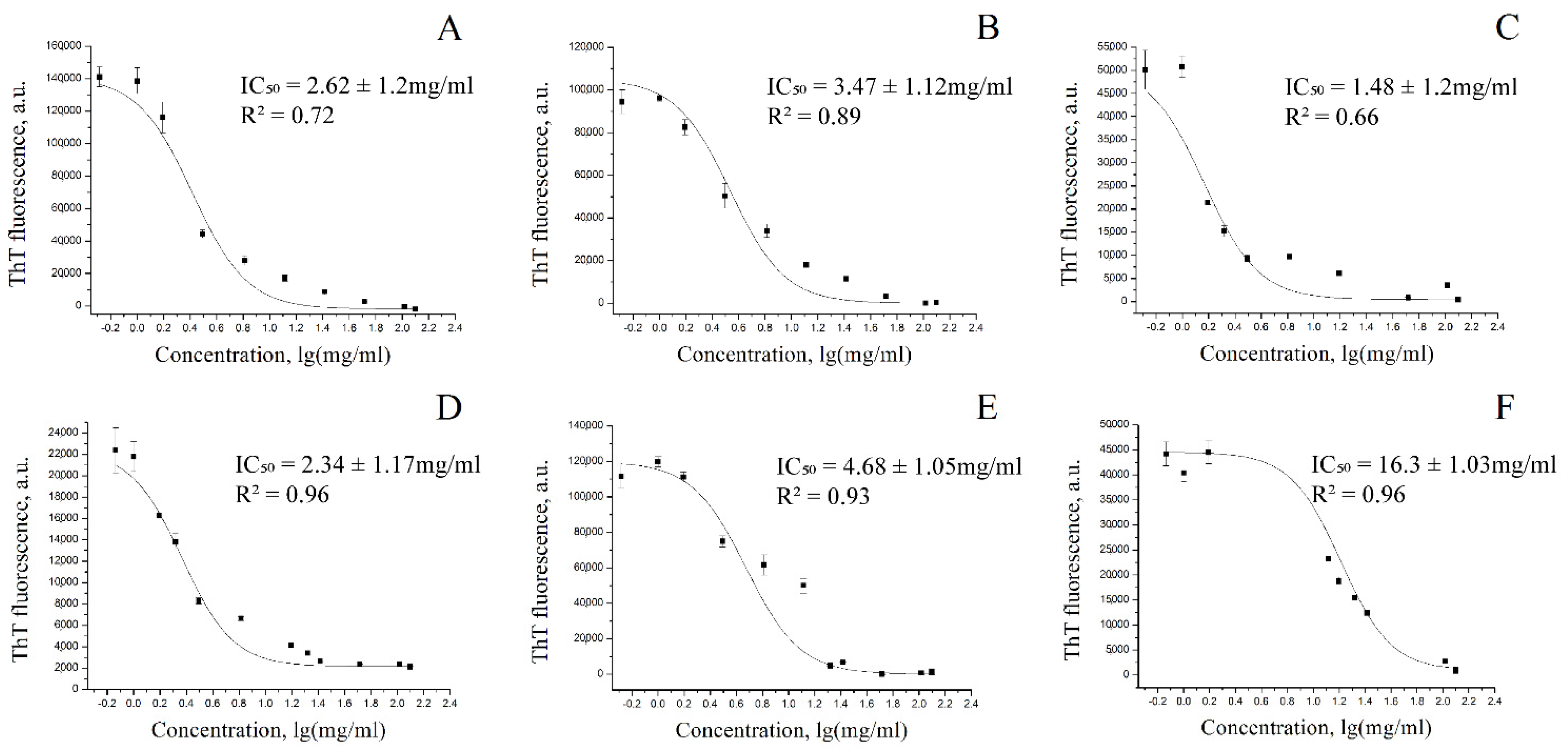
3.1. Determination of the Half-Maximal Inhibition Constant of Alpha-Synuclein Fibrillization for Coffee Extracts
3.2. Determination of the Inhibition Constant Ki
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharifi-Rad, M.; Lankatillake, C.; Dias, D.A.; Docea, A.O.; Mahomoodally, M.F.; Lobine, D.; Chazot, P.L.; Kurt, B.; Boyunegmez Tumer, T.; Catarina Moreira, A.; et al. Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics. J. Clin. Med. 2020, 9, 1061. [Google Scholar] [CrossRef] [PubMed]
- Ruwizhi, N.; Aderibigbe, B.A. Cinnamic Acid Derivatives and Their Biological Efficacy. Int. J. Mol. Sci. 2020, 21, 5712. [Google Scholar] [CrossRef]
- Hewlings, S.; Kalman, D. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.; Alsahli, M.; Aly, S.; Khan, M.; Aldebasi, Y. Role of Curcumin in Disease Prevention and Treatment. Adv. Biomed. Res. 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Darvesh, A.S.; Carroll, R.T.; Bishayee, A.; Novotny, N.A.; Geldenhuys, W.J.; Van der Schyf, C.J. Curcumin and Neurodegenerative Diseases: A Perspective. Expert Opin. Investig. Drugs 2012, 21, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Maiti, P.; Ma, Q.; Zuo, X.; Jones, M.R.; Cole, G.M.; Frautschy, S.A. Clinical Development of Curcumin in Neurodegenerative Disease. Expert Rev. Neurother. 2015, 15, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Mythri, R.B.; Bharath, M.M.S. Curcumin: A Potential Neuroprotective Agent in Parkinson’s Disease. Curr. Pharm. Des. 2012, 18, 91–99. [Google Scholar] [CrossRef]
- Lee, W.-H.; Loo, C.-Y.; Bebawy, M.; Luk, F.; Mason, R.; Rohanizadeh, R. Curcumin and Its Derivatives: Their Application in Neuropharmacology and Neuroscience in the 21st Century. Curr. Neuropharmacol. 2013, 11, 338–378. [Google Scholar] [CrossRef]
- Zanyatkin, I.; Stroylova, Y.; Tishina, S.; Stroylov, V.; Melnikova, A.; Haertle, T.; Muronetz, V. Inhibition of Prion Propagation by 3,4-Dimethoxycinnamic Acid: Inhibition of Prion Propagation by 3,4-Dimethoxycinnamic Acid. Phytother. Res. 2017, 31, 1046–1055. [Google Scholar] [CrossRef]
- Tishina, S.A.; Stroylov, V.S.; Zanyatkin, I.A.; Melnikova, A.K.; Muronetz, V.I.; Stroylova, Y.Y. Cinnamic Acid Derivatives as the Potential Modulators of Prion Aggregation. Mendeleev Commun. 2017, 27, 493–494. [Google Scholar] [CrossRef]
- Medvedeva, M.; Barinova, K.; Melnikova, A.; Semenyuk, P.; Kolmogorov, V.; Gorelkin, P.; Erofeev, A.; Muronetz, V. Naturally Occurring Cinnamic Acid Derivatives Prevent Amyloid Transformation of Alpha-Synuclein. Biochimie 2020, 170, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Muronetz, V.I.; Barinova, K.; Kudryavtseva, S.; Medvedeva, M.; Melnikova, A.; Sevostyanova, I.; Semenyuk, P.; Stroylova, Y.; Sova, M. Natural and Synthetic Derivatives of Hydroxycinnamic Acid Modulating the Pathological Transformation of Amyloidogenic Proteins. Molecules 2020, 25, 4647. [Google Scholar] [CrossRef] [PubMed]
- Freyssin, A.; Page, G.; Fauconneau, B.; Bilan, A. Natural Polyphenols Effects on Protein Aggregates in Alzheimer’s and Parkinson’s Prion-like Diseases. Neural Regen. Res. 2018, 13, 955. [Google Scholar] [CrossRef] [PubMed]
- Marchiani, A.; Mammi, S.; Siligardi, G.; Hussain, R.; Tessari, I.; Bubacco, L.; Delogu, G.; Fabbri, D.; Dettori, M.A.; Sanna, D.; et al. Small Molecules Interacting with α-Synuclein: Antiaggregating and Cytoprotective Properties. Amino Acids 2013, 45, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yamada, M. Antioxidant Compounds Have Potent Anti-Fibrillogenic and Fibril-Destabilizing Effects for Alpha-Synuclein Fibrils In Vitro. J. Neurochem. 2006, 97, 105–115. [Google Scholar] [CrossRef]
- Caruana, M.; Högen, T.; Levin, J.; Hillmer, A.; Giese, A.; Vassallo, N. Inhibition and Disaggregation of α-Synuclein Oligomers by Natural Polyphenolic Compounds. FEBS Lett. 2011, 585, 1113–1120. [Google Scholar] [CrossRef]
- Fazili, N.A.; Naeem, A. Anti-Fibrillation Potency of Caffeic Acid against an Antidepressant Induced Fibrillogenesis of Human α-Synuclein: Implications for Parkinson’s Disease. Biochimie 2015, 108, 178–185. [Google Scholar] [CrossRef]
- Hase, T.; Shishido, S.; Yamamoto, S.; Yamashita, R.; Nukima, H.; Taira, S.; Toyoda, T.; Abe, K.; Hamaguchi, T.; Ono, K.; et al. Rosmarinic Acid Suppresses Alzheimer’s Disease Development by Reducing Amyloid β Aggregation by Increasing Monoamine Secretion. Sci. Rep. 2019, 9, 8711. [Google Scholar] [CrossRef]
- Yang, L.; Wang, N.; Zheng, G. Enhanced Effect of Combining Chlorogenic Acid on Selenium Nanoparticles in Inhibiting Amyloid β Aggregation and Reactive Oxygen Species Formation In Vitro. Nanoscale Res. Lett. 2018, 13, 303. [Google Scholar] [CrossRef]
- Mancini, R.S.; Wang, Y.; Weaver, D.F. Phenylindanes in Brewed Coffee Inhibit Amyloid-Beta and Tau Aggregation. Front. Neurosci. 2018, 12, 735. [Google Scholar] [CrossRef]
- Ma, L.; Zheng, J.; Chen, H.; Zeng, X.; Wang, S.; Yang, C.; Li, X.; Xiao, Y.; Zheng, L.; Chen, H.; et al. A Systematic Screening of Traditional Chinese Medicine Identifies Two Novel Inhibitors against the Cytotoxic Aggregation of Amyloid Beta. Front. Pharmacol. 2021, 12, 637766. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Corona, G.; Spencer, J.P.E. Caffeic Acid, Tyrosol and p-Coumaric Acid Are Potent Inhibitors of 5-S-Cysteinyl-Dopamine Induced Neurotoxicity. Arch. Biochem. Biophys. 2010, 501, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.M. The Shikimate Pathway as an Entry to Aromatic Secondary Metabolism. Plant Physiol. 1995, 107, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.; Gaspar, A.; Garrido, E.M.; Garrido, J.; Borges, F. Hydroxycinnamic Acid Antioxidants: An Electrochemical Overview. BioMed Res. Int. 2013, 2013, 251754. [Google Scholar] [CrossRef]
- Stalmach, A.; Mullen, W.; Barron, D.; Uchida, K.; Yokota, T.; Cavin, C.; Steiling, H.; Williamson, G.; Crozier, A. Metabolite Profiling of Hydroxycinnamate Derivatives in Plasma and Urine after the Ingestion of Coffee by Humans: Identification of Biomarkers of Coffee Consumption. Drug Metab. Dispos. 2009, 37, 1749–1758. [Google Scholar] [CrossRef]
- Sova, M.; Saso, L. Natural Sources, Pharmacokinetics, Biological Activities and Health Benefits of Hydroxycinnamic Acids and Their Metabolites. Nutrients 2020, 12, 2190. [Google Scholar] [CrossRef]
- Farrell, T.L.; Gomez-Juaristi, M.; Poquet, L.; Redeuil, K.; Nagy, K.; Renouf, M.; Williamson, G. Absorption of Dimethoxycinnamic Acid Derivatives In Vitro and Pharmacokinetic Profile in Human Plasma Following Coffee Consumption. Mol. Nutr. Food Res. 2012, 56, 1413–1423. [Google Scholar] [CrossRef]
- Grosso, G.; Stepaniak, U.; Topor-Mądry, R.; Szafraniec, K.; Pająk, A. Estimated Dietary Intake and Major Food Sources of Polyphenols in the Polish Arm of the HAPIEE Study. Nutrition 2014, 30, 1398–1403. [Google Scholar] [CrossRef]
- Clifford, M.N. Chlorogenic Acids and Other Cinnamates—Nature, Occurrence, Dietary Burden, Absorption and Metabolism. J. Sci. Food Agric. 2000, 80, 1033–1043. [Google Scholar] [CrossRef]
- Kroon, P.A.; Clifford, M.N.; Crozier, A.; Day, A.J.; Donovan, J.L.; Manach, C.; Williamson, G. How Should We Assess the Effects of Exposure to Dietary Polyphenols In Vitro? Am. J. Clin. Nutr. 2004, 80, 15–21. [Google Scholar] [CrossRef]
- Nagy, K.; Redeuil, K.; Williamson, G.; Rezzi, S.; Dionisi, F.; Longet, K.; Destaillats, F.; Renouf, M. First Identification of Dimethoxycinnamic Acids in Human Plasma after Coffee Intake by Liquid Chromatography–Mass Spectrometry. J. Chromatogr. A 2011, 1218, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Dohmae, N.; Nonaka, T.; Oikawa, T.; Hisanaga, S.; Goedert, M.; Hasegawa, M. Cysteine Misincorporation in Bacterially Expressed Human Alpha-Synuclein. FEBS Lett. 2006, 580, 1775–1779. [Google Scholar] [CrossRef] [PubMed]
- Barinova, K.V.; Kuravsky, M.L.; Arutyunyan, A.M.; Serebryakova, M.V.; Schmalhausen, E.V.; Muronetz, V.I. Dimerization of Tyr136Cys Alpha-Synuclein Prevents Amyloid Transformation of Wild Type Alpha-Synuclein. Int. J. Biol. Macromol. 2017, 96, 35–43. [Google Scholar] [CrossRef] [PubMed]
- OriginLab. Available online: https://Www.Originlab.Com/Index.Aspx?Go=Support&pid=1838 (accessed on 18 April 2022).
- Lapidus, L.J.; Eaton, W.A.; Hofrichter, J. Measuring the Rate of Intramolecular Contact Formation in Polypeptides. Proc. Natl. Acad. Sci. USA 2000, 97, 7220–7225. [Google Scholar] [CrossRef]
- Ahmad, B.; Lapidus, L.J. Curcumin Prevents Aggregation in α-Synuclein by Increasing Reconfiguration Rate. J. Biol. Chem. 2012, 287, 9193–9199. [Google Scholar] [CrossRef]
- Lee, K.-W.; Im, J.-Y.; Woo, J.-M.; Grosso, H.; Kim, Y.-S.; Cristovao, A.C.; Sonsalla, P.K.; Schuster, D.S.; Jalbut, M.M.; Fernandez, J.R.; et al. Neuroprotective and Anti-Inflammatory Properties of a Coffee Component in the MPTP Model of Parkinson’s Disease. Neurotherapeutics 2013, 10, 143–153. [Google Scholar] [CrossRef]
- Tsai, S.; Chao, C.; Yin, M. Preventive and Therapeutic Effects of Caffeic Acid against Inflammatory Injury in Striatum of MPTP-Treated Mice. Eur. J. Pharmacol. 2011, 670, 441–447. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Zhang, L.; Wang, Q.; Yang, Z.; Liu, J.; Feng, L. Caffeic Acid Reduces A53T α-Synuclein by Activating JNK/Bcl-2-Mediated Autophagy In Vitro and Improves Behaviour and Protects Dopaminergic Neurons in a Mouse Model of Parkinson’s Disease. Pharmacol. Res. 2019, 150, 104538. [Google Scholar] [CrossRef]
- Ishida, K.; Yamamoto, M.; Misawa, K.; Nishimura, H.; Misawa, K.; Ota, N.; Shimotoyodome, A. Coffee polyphenols prevent cognitive dysfunction and suppress amyloid β plaques in APP/PS2 transgenic mouse. Neurosci. Res. 2020, 154, 35–44. [Google Scholar] [CrossRef]
- Gardener, S.L.; Rainey-Smith, S.R.; Villemagne, V.L.; Fripp, J.; Doré, V.; Bourgeat, P.; Taddei, K.; Fowler, C.; Masters, C.L.; Maruff, P.; et al. Higher Coffee Consumption Is Associated With Slower Cognitive Decline and Less Cerebral Aβ-Amyloid Accumulation over 126 Months: Data from the Australian Imaging, Biomarkers, and Lifestyle Study. Front. Aging Neurosci. 2021, 3, 744872. [Google Scholar] [CrossRef]
- Kim, J.W.; Byun, M.S.; Yi, D.; Lee, J.H.; Jeon, S.Y.; Jung, G.; Lee, H.N.; Sohn, B.K.; Lee, J.; Kim, Y.K.; et al. Coffee intake and decreased amyloid pathology in human brain. Transl. Psychiatry 2019, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.L. Wine Phenolics. Ann. N. Y. Acad. Sci. 2002, 957, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Maurya, D.K.; Devasagayam, T.P.A. Antioxidant and Prooxidant Nature of Hydroxycinnamic Acid Derivatives Ferulic and Caffeic Acids. Food Chem. Toxicol. 2010, 48, 3369–3373. [Google Scholar] [CrossRef] [PubMed]
- Basli, A.; Soulet, S.; Chaher, N.; Mérillon, J.-M.; Chibane, M.; Monti, J.-P.; Richard, T. Wine Polyphenols: Potential Agents in Neuroprotection. Oxidative Med. Cell. Longev. 2012, 2012, 805762. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Neuroprotective Potential of Ferulic Acid in the Rotenone Model of Parkinson’s Disease. Drug Des. Dev. Ther. 2015, 9, 5499–5510. [Google Scholar] [CrossRef]
- Wang, E.-J.; Wu, M.-Y.; Lu, J.-H. Ferulic Acid in Animal Models of Alzheimer’s Disease: A Systematic Review of Preclinical Studies. Cells 2021, 10, 2653. [Google Scholar] [CrossRef]
- Sgarbossa, A.; Giacomazza, D.; di Carlo, M. Ferulic Acid: A Hope for Alzheimer’s Disease Therapy from Plants. Nutrients 2015, 7, 5764–5782. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | MRM Transition | DP (Volts) | EP (Volts) | CE (Volts) | CXP (Volts) | |
|---|---|---|---|---|---|---|
| Ferulic acid | 195.031 | 177.1 | 1 | 10 | 15 | 14 |
| Ferulic acid | 195.031 | 89 | 1 | 10 | 39 | 14 |
| 3,4-Dimethoxycinnamic acid | 209.039 | 191 | 1 | 10 | 15 | 14 |
| 3,4-Dimethoxycinnamic acid | 209.039 | 163 | 1 | 10 | 27 | 20 |
| 3-Methoxy-4-acetamidoxycinnamic acid | 252.036 | 191.9 | 1 | 10 | 29 | 16 |
| 3-Methoxy-4-acetamidoxycinnamic acid | 252.036 | 207 | 1 | 10 | 15 | 18 |
| 1 | 3-Methoxy-4-acetamidoxycinnamic acid | 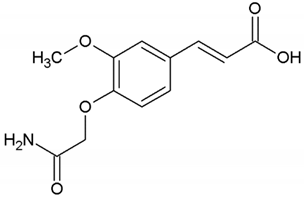 |
| 2 | Ferulic acid (3-methoxy-4-hydroxycinnamic acid) | 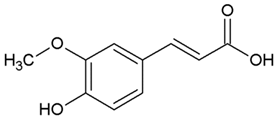 |
| 3 | Dimethoxycinnamic acid (3,4DMCA) | 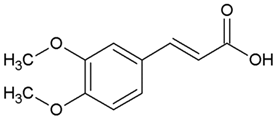 |
| 4 | Curcumin |  |
| Sample Name | 3,4DMCA, µg/mL | 3M4ACA, µg/mL | FA, ng/mL |
|---|---|---|---|
| Black coffee, mQ water | 53.26 | 0.81 | 0.22 |
| Black coffee, mQ water, heating | 48.51 | 0.75 | 0.21 |
| Black coffee, ethanol | 44.06 | 0.40 | 0.13 |
| Green coffee, mQ water | 25.18 | 0.02 | 1.29 |
| Green coffee, mQ water, heating | 25.92 | 0.05 | 3.02 |
| Green coffee, ethanol | 12.96 | 0.01 | 1.55 |
| Sample Name | Ki for 3,4DMCA, μM |
|---|---|
| Black coffee, mQ water | 1.12 ± 0.1 |
| Black coffee, mQ water, heating | 1.81 ± 0.17 |
| Black coffee, ethanol | 0.89 ± 0.04 |
| Green coffee, mQ water | 1.81 ± 0.09 |
| Green coffee, mQ water, heating | 4.47 ± 0.6 |
| Green coffee, ethanol | 2.88 ± 2.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medvedeva, M.; Kitsilovskaya, N.; Stroylova, Y.; Sevostyanova, I.; Saboury, A.A.; Muronetz, V. Hydroxycinnamic Acid Derivatives from Coffee Extracts Prevent Amyloid Transformation of Alpha-Synuclein. Biomedicines 2022, 10, 2255. https://doi.org/10.3390/biomedicines10092255
Medvedeva M, Kitsilovskaya N, Stroylova Y, Sevostyanova I, Saboury AA, Muronetz V. Hydroxycinnamic Acid Derivatives from Coffee Extracts Prevent Amyloid Transformation of Alpha-Synuclein. Biomedicines. 2022; 10(9):2255. https://doi.org/10.3390/biomedicines10092255
Chicago/Turabian StyleMedvedeva, Maria, Natalia Kitsilovskaya, Yulia Stroylova, Irina Sevostyanova, Ali Akbar Saboury, and Vladimir Muronetz. 2022. "Hydroxycinnamic Acid Derivatives from Coffee Extracts Prevent Amyloid Transformation of Alpha-Synuclein" Biomedicines 10, no. 9: 2255. https://doi.org/10.3390/biomedicines10092255
APA StyleMedvedeva, M., Kitsilovskaya, N., Stroylova, Y., Sevostyanova, I., Saboury, A. A., & Muronetz, V. (2022). Hydroxycinnamic Acid Derivatives from Coffee Extracts Prevent Amyloid Transformation of Alpha-Synuclein. Biomedicines, 10(9), 2255. https://doi.org/10.3390/biomedicines10092255