
Effect of D1- and D2-like Dopamine Receptor Antagonists on the Rewarding and Anxiolytic Effects of Neurotensin in the Ventral Pallidum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Surgery
2.3. Drugs
2.4. Microinjection Procedure
2.5. Behavioural Testing
2.5.1. Conditioned Place Preference (CPP) Test
2.5.2. Elevated plus Maze (EPM) Test
2.6. Histology
2.7. Statistical Analysis
3. Results
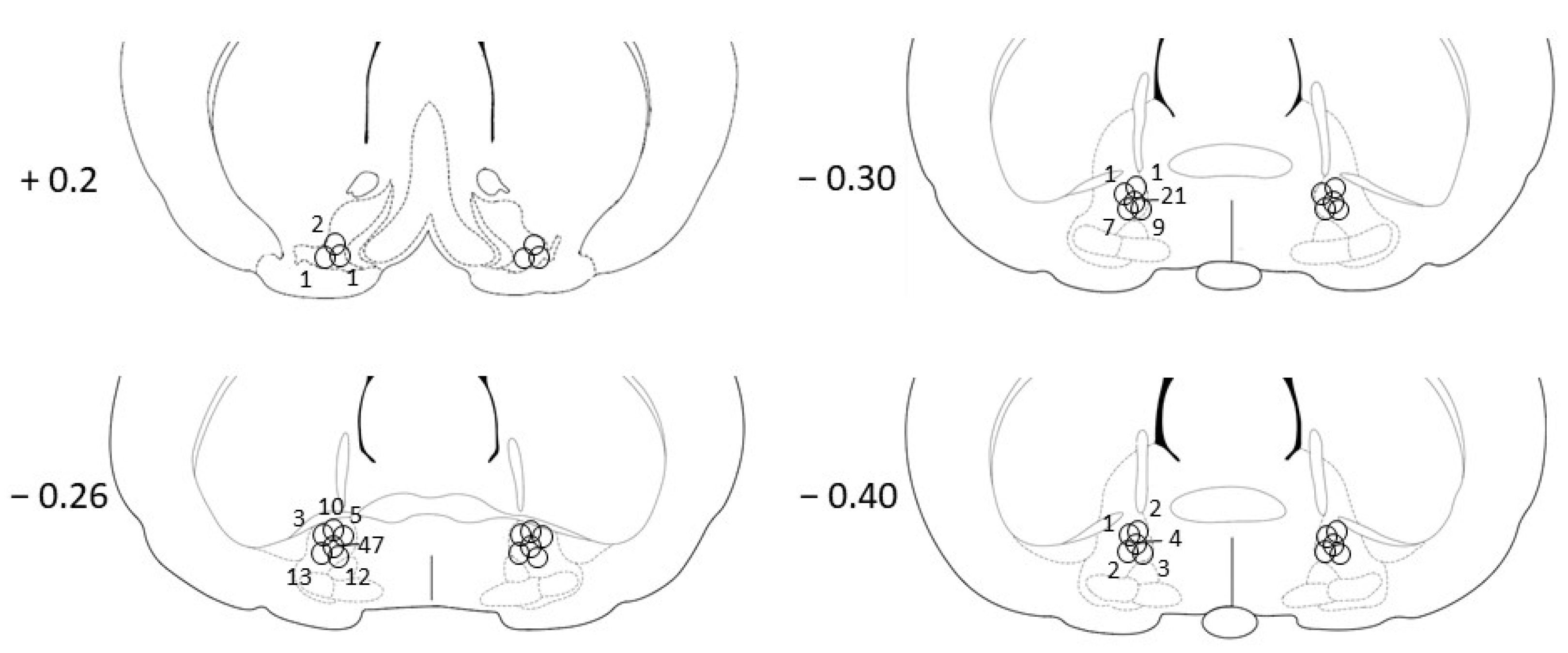
3.1. Histology
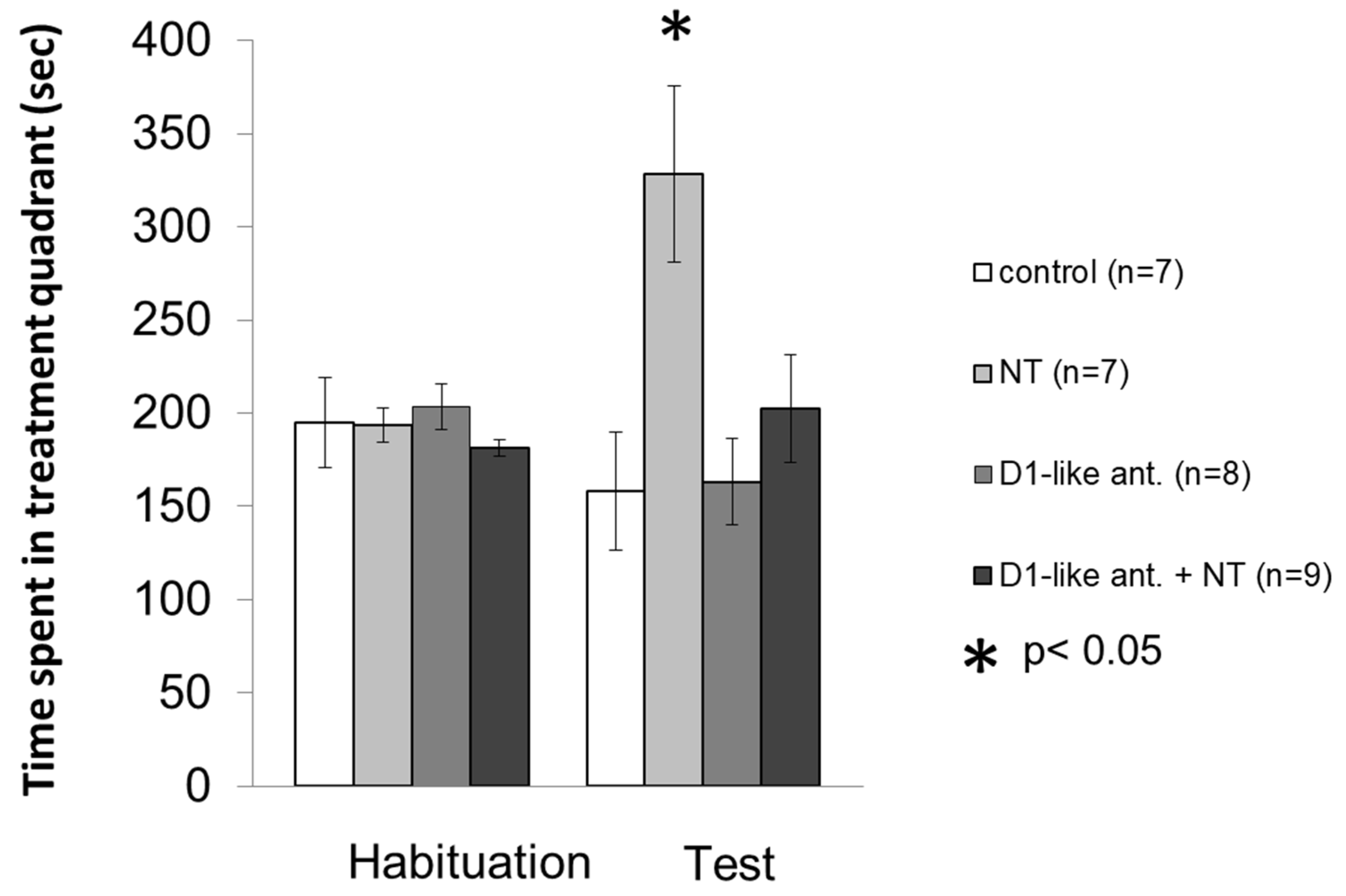
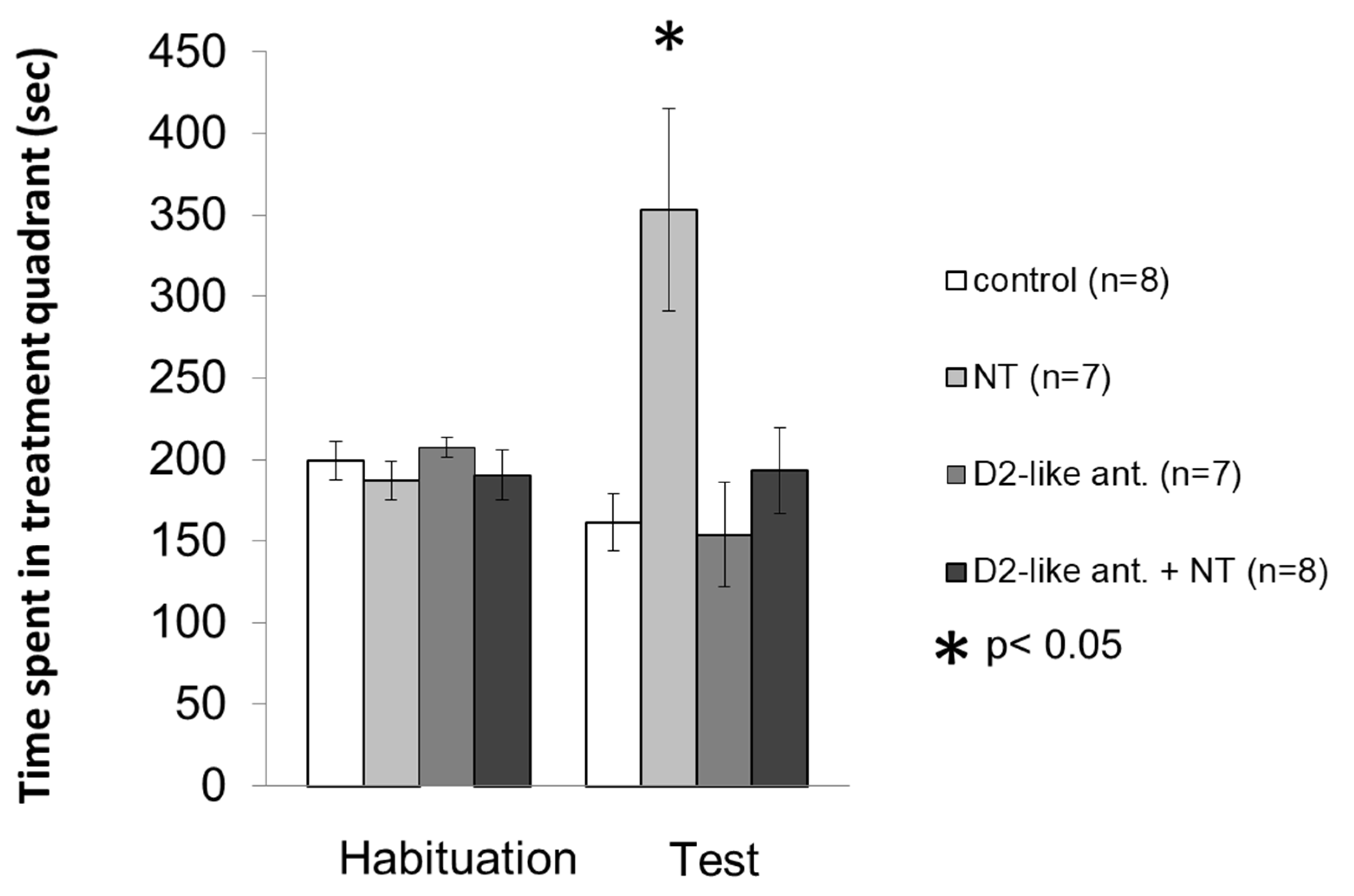
3.2. Conditioned Place Preference Test
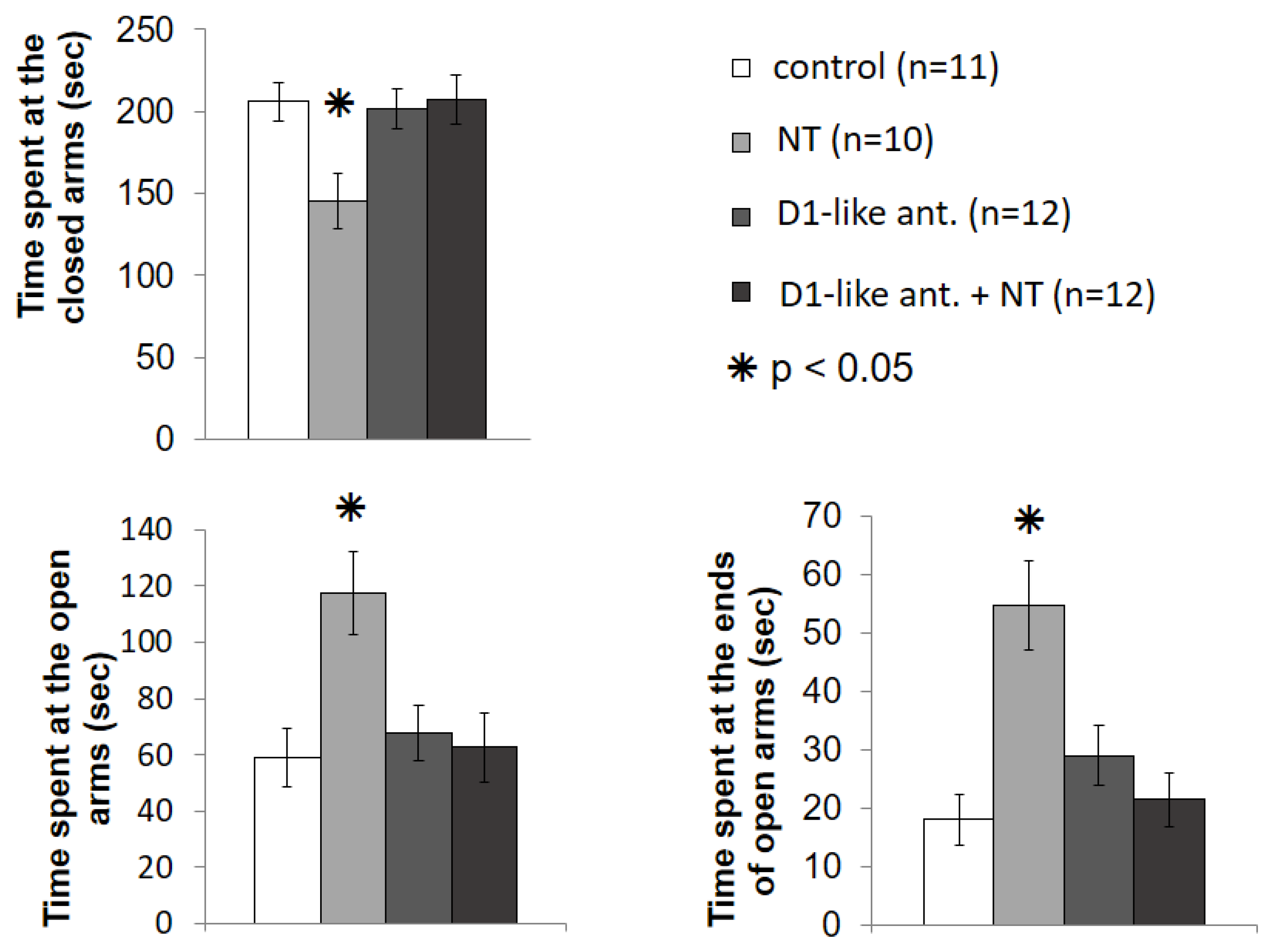
3.3. Elevated plus Maze Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Charlson, F.J.; Ferrari, A.J.; Santomauro, D.F.; Diminic, S.; Stockings, E.; Scott, J.G.; McGrath, J.J.; Whiteford, H.A. Global Epidemiology and Burden of Schizophrenia: Findings from the Global Burden of Disease Study 2016. Schizophr. Bull. 2018, 44, 1195–1203. [Google Scholar] [PubMed]
- Baudonnat, M.; Huber, A.; David, V.; Walton, M.E. Heads for learning, tails for memory: Reward, reinforcement and a role of dopamine in determining behavioral relevance across multiple timescales. Front. Neurosci. 2013, 7, 175. [Google Scholar] [PubMed]
- Zarrindast, M.R.; Khakpai, F. The Modulatory Role of Dopamine in Anxiety-like Behavior. Arch. Iran. Med. 2015, 18, 591–603. [Google Scholar] [PubMed]
- Ferraro, L.; Tiozzo Fasiolo, L.; Beggiato, S.; Borelli, A.C.; Pomierny-Chamiolo, L.; Frankowska, M.; Antonelli, T.; Tomasini, M.C.; Fuxe, K.; Filip, M. Neurotensin: A role in substance use disorder? J. Psychopharmacol. 2016, 30, 112–127. [Google Scholar]
- Normandeau, C.P.; Ventura-Silva, A.P.; Hawken, E.R.; Angelis, S.; Sjaarda, C.; Liu, X.; Pego, J.M.; Dumont, E.C. A Key Role for Neurotensin in Chronic-Stress-Induced Anxiety-Like Behavior in Rats. Neuropsychopharmacology 2018, 43, 285–293. [Google Scholar]
- Torruella-Suárez, M.L.; McElligott, Z.A. Neurotensin in reward processes. Neuropharmacology 2020, 167, 108005. [Google Scholar]
- Lénárd, L.; László, K.; Kertes, E.; Ollmann, T.; Péczely, L.; Kovács, A.; Kállai, V.; Zagoracz, O.; Gálosi, R.; Karádi, Z. Substance P and neurotensin in the limbic system: Their roles in reinforcement and memory consolidation. Neurosci. Biobehav. Rev. 2018, 85, 1–20. [Google Scholar]
- Baik, J.H. Stress and the dopaminergic reward system. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar]
- Heimer, L.; Wilson, R.D. The subcortical projections of the allocortex: Similarities in the neural associations of the hippocampus, the piriform cortex, and the neocortex. In Golgi Centennial Symposium: Perspectives in Neurobiology; Santini, M., Ed.; Raven Press: New York, NY, USA, 1975; pp. 177–193. [Google Scholar]
- Klitenick, M.A.; Deutch, A.Y.; Churchill, L.; Kalivas, P.W. Topography and functional role of dopaminergic projections from the ventral mesencephalic tegmentum to the ventral pallidum. Neuroscience 1992, 50, 371–386. [Google Scholar]
- Panagis, G.; Spyraki, C. Neuropharmacological evidence for the role of dopamine in ventral pallidum self-stimulation. Psychopharmacology 1996, 123, 280–288. [Google Scholar]
- Gong, W.; Neill, D.; Justice, J.B., Jr. 6-Hydroxydopamine lesion of ventral pallidum blocks acquisition of place preference conditioning to cocaine. Brain Res. 1997, 754, 103–112. [Google Scholar] [CrossRef]
- Boyson, S.J.; McGonigle, P.; Molinoff, P.B. Quantitative autoradiographic localization of the D1 and D2 subtypes of dopamine receptors in rat brain. J. Neurosci. 1986, 6, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.S.; Jian, M. D1 and D2 dopamine receptors differentially increase Fos-like immunoreactivity in accumbal projections to the ventral pallidum and midbrain. Neuroscience 1995, 64, 1019–1034. [Google Scholar] [CrossRef]
- Mansour, A.; Meador-Woodruff, J.H.; Zhou, Q.; Civelli, O.; Akil, H.; Watson, S.J. A comparison of D1 receptor binding and mRNA in rat brain using receptor autoradiographic and in situ hybridization techniques. Neuroscience 1992, 46, 959–971. [Google Scholar] [CrossRef]
- Smith, Y.; Kieval, J.Z. Anatomy of the dopamine system in the basal ganglia. Trends Neurosci. 2000, 23, S28–S33. [Google Scholar] [CrossRef]
- Mengual, E.; Pickel, V.M. Ultrastructural immunocytochemical localization of the dopamine D2 receptor and tyrosine hydroxylase in the rat ventral pallidum. Synapse 2002, 43, 151–162. [Google Scholar] [CrossRef]
- Glimcher, P.W.; Margolin, D.H.; Giovino, A.A.; Hoebel, B.G. Neurotensin: A new ‘reward peptide’. Brain Res. 1984, 291, 119–124. [Google Scholar] [CrossRef]
- Bauco, P.; Rompré, P.P. Effects of neurotensin receptor activation on brain stimulation reward in Fischer 344 and Lewis rats. Eur. J. Pharm. 2001, 432, 57–61. [Google Scholar] [CrossRef]
- László, K.; Tóth, K.; Kertes, E.; Péczely, L.; Lénárd, L. The role of neurotensin in positive reinforcement in the rat central nucleus of amygdala. Behav. Brain Res. 2010, 208, 430–435. [Google Scholar] [CrossRef]
- Hou, I.C.; Suzuki, C.; Kanegawa, N.; Oda, A.; Yamada, A.; Yoshikawa, M.; Yamada, D.; Sekiguchi, M.; Wada, E.; Wada, K.; et al. beta-Lactotensin derived from bovine beta-lactoglobulin exhibits anxiolytic-like activity as an agonist for neurotensin NTS2 receptor via activation of dopamine D1 receptor in mice. J. Neurochem. 2011, 119, 785–790. [Google Scholar] [CrossRef]
- Steele, F.F., 3rd; Whitehouse, S.C.; Aday, J.S.; Prus, A.J. Neurotensin NTS1 and NTS2 receptor agonists produce anxiolytic-like effects in the 22-kHz ultrasonic vocalization model in rats. Brain Res. 2017, 1658, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Carraway, R.; Leeman, S.E. The isolation of a new hypotensive peptide, neurotensin, from bovine hypothalami. J. Biol. Chem. 1973, 248, 6854–6861. [Google Scholar] [CrossRef]
- Ferraro, L.; Tomasini, M.C.; Fuxe, K.; Agnati, L.F.; Mazza, R.; Tanganelli, S.; Antonelli, T. Mesolimbic dopamine and cortico-accumbens glutamate afferents as major targets for the regulation of the ventral striato-pallidal GABA pathways by neurotensin peptides. Brain Res. Rev. 2007, 55, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.B.; Kinkead, B.; Owens, M.J.; Nemeroff, C.B. Neurotensin and dopamine interactions. Pharm. Rev. 2001, 53, 453–486. [Google Scholar] [PubMed]
- Ferraro, L.; Beggiato, S.; Tomasini, M.C.; Antonelli, T.; Tanganelli, S.; Fuxe, K. Neurotensin NTS1-dopamine D2 receptor-receptor interactions in putative receptor heteromers: Relevance for schizophrenia. Curr. Protein Pept. Sci. 2014, 15, 681–690. [Google Scholar] [CrossRef] [PubMed]
- László, K.; Péczely, L.; Kovács, A.; Zagoracz, O.; Ollmann, T.; Kertes, E.; Kállai, V.; Csetényi, B.; Karádi, Z.; Lénárd, L. The role of intraamygdaloid neurotensin and dopamine interaction in conditioned place preference. Behav. Brain Res. 2018, 344, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Zahm, D.S.; Heimer, L. Ventral striatopallidal parts of the basal ganglia in the rat: I. Neurochemical compartmentation as reflected by the distributions of neurotensin and substance P immunoreactivity. J. Comp. Neurol. 1988, 272, 516–535. [Google Scholar] [CrossRef]
- Zahm, D.S. The ventral striatopallidal parts of the basal ganglia in the rat—II. Compartmentation of ventral pallidal efferents. Neuroscience 1989, 30, 33–50. [Google Scholar] [CrossRef]
- Root, D.H.; Melendez, R.I.; Záborszky, L.; Napier, T.C. The ventral pallidum: Subregion-specific functional anatomy and roles in motivated behaviors. Prog. Neurobiol. 2015, 130, 29–70. [Google Scholar] [CrossRef]
- Ollmann, T.; Péczely, L.; László, K.; Kovács, A.; Gálosi, R.; Berente, E.; Karádi, Z.; Lénárd, L. Positive reinforcing effect of neurotensin microinjection into the ventral pallidum in conditioned place preference test. Behav. Brain Res. 2015, 278, 470–475. [Google Scholar] [CrossRef]
- Ollmann, T.; Péczely, L.; László, K.; Kovács, A.; Gálosi, R.; Kertes, E.; Kállai, V.; Zagoracz, O.; Karádi, Z.; Lénárd, L. Anxiolytic effect of neurotensin microinjection into the ventral pallidum. Behav. Brain Res. 2015, 294, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Hasenöhrl, R.U.; Oitzl, M.S.; Huston, J.P. Conditioned place preference in the corral: A procedure for measuring reinforcing properties of drugs. J. Neurosci. Methods 1989, 30, 141–146. [Google Scholar] [CrossRef]
- Huston, J.P.; Silva, M.A.; Topic, B.; Müller, C.P. What’s conditioned in conditioned place preference? Trends Pharm. Sci. 2013, 34, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Privou, C.; Huston, J.P. Differential sensitivity of the caudal and rostral nucleus accumbens to the rewarding effects of a H1-histaminergic receptor blocker as measured with place-preference and self-stimulation behavior. Neuroscience 1999, 94, 93–103. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Hiroi, N.; White, N.M. The ventral pallidum area is involved in the acquisition but not expression of the amphetamine conditioned place preference. Neurosci. Lett. 1993, 156, 9–12. [Google Scholar] [CrossRef]
- McAlonan, G.M.; Robbins, T.W.; Everitt, B.J. Effects of medial dorsal thalamic and ventral pallidal lesions on the acquisition of a conditioned place preference: Further evidence for the involvement of the ventral striatopallidal system in reward-related processes. Neuroscience 1993, 52, 605–620. [Google Scholar] [CrossRef]
- Jay, T.M. Dopamine: A potential substrate for synaptic plasticity and memory mechanisms. Prog. Neurobiol. 2003, 69, 375–390. [Google Scholar] [CrossRef]
- Clark, M.; Bracci, E. Dichotomous dopaminergic control of ventral pallidum neurons. Front. Cell. Neurosci. 2018, 12, 260. [Google Scholar] [CrossRef]
- Stout, K.A.; Dunn, A.R.; Lohr, K.M.; Alter, S.P.; Cliburn, R.A.; Guillot, T.S.; Miller, G.W. Selective Enhancement of Dopamine Release in the Ventral Pallidum of Methamphetamine-Sensitized Mice. ACS Chem. Neurosci. 2016, 7, 1364–1373. [Google Scholar] [CrossRef]
- Gong, W.; Neill, D.; Justice, J.B., Jr. Conditioned place preference and locomotor activation produced by injection of psychostimulants into ventral pallidum. Brain Res. 1996, 707, 64–74. [Google Scholar] [CrossRef]
- Gong, W.; Neill, D.B.; Justice, J.B., Jr. GABAergic modulation of ventral pallidal dopamine release studied by in vivo microdialysis in the freely moving rat. Synapse 1998, 29, 406–412. [Google Scholar] [CrossRef]
- Skoubis, P.D.; Maidment, N.T. Blockade of ventral pallidal opioid receptors induces a conditioned place aversion and attenuates acquisition of cocaine place preference in the rat. Neuroscience 2003, 119, 241–249. [Google Scholar] [CrossRef]
- Péczely, L.; Ollmann, T.; László, K.; Kovács, A.; Gálosi, R.; Kertes, E.; Zagoracz, O.; Kállai, V.; Karádi, Z.; Lénárd, L. Role of ventral pallidal D2 dopamine receptors in the consolidation of spatial memory. Behav. Brain Res. 2016, 313, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, E.V.; Stavrovskaia, A.V.; Shugalev, N.P.; Lénárd, L.; Hartmann, G. Emotional state variations in rats during recall of passive avoidance reactions after neurotensin administration into nucleus accumbens of the brain. Zhurnal Vysshei Nervnoi Deiatelnosti Imeni IP Pavlova 2010, 60, 704–711. [Google Scholar]
- Shugalev, N.P.; Stavrovskaia, A.V.; Iamshchikova, N.G.; Ol’shanskiĭ, A.S.; Miroshnichenko, E.V. Reproduction of passive avoidance reactions after neurotensin microinjection into nucleus accumbens of rat brain against the background of reserpine action. Zhurnal Vysshei Nervnoi Deiatelnosti Imeni IP Pavlova 2012, 62, 357–363. [Google Scholar]
- Danjo, T.; Yoshimi, K.; Funabiki, K.; Yawata, S.; Nakanishi, S. Aversive behavior induced by optogenetic inactivation of ventral tegmental area dopamine neurons is mediated by dopamine D2 receptors in the nucleus accumbens. Proc. Natl. Acad. Sci. USA 2014, 111, 6455–6460. [Google Scholar] [CrossRef]
- Mahler, S.V.; Vazey, E.M.; Beckley, J.T.; Keistler, C.R.; McGlinchey, E.M.; Kaufling, J.; Wilson, S.P.; Deisseroth, K.; Woodward, J.J.; Aston-Jones, G. Designer receptors show role for ventral pallidum input to ventral tegmental area in cocaine seeking. Nat. Neurosci. 2014, 17, 577–585. [Google Scholar] [CrossRef]
- Valenti, O.; Grace, A.A. Antipsychotic drug-induced increases in ventral tegmental area dopamine neuron population activity via activation of the nucleus accumbens-ventral pallidum pathway. Int. J. Neuropsychopharmacol. 2010, 13, 845–860. [Google Scholar] [CrossRef] [Green Version]
- Calder, A.J.; Beaver, J.D.; Davis, M.H.; van Ditzhuijzen, J.; Keane, J.; Lawrence, A.D. Disgust sensitivity predicts the insula and pallidal response to pictures of disgusting foods. Eur. J. Neurosci. 2007, 25, 3422–3428. [Google Scholar] [CrossRef]
- Floresco, S.B.; West, A.R.; Ash, B.; Moore, H.; Grace, A.A. Afferent modulation of dopamine neuron firing differentially regulates tonic and phasic dopamine transmission. Nat. Neurosci. 2003, 6, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Fibiger, H.C.; LePiane, F.G.; Jakubovic, A.; Phillips, A.G. The role of dopamine in intracranial self-stimulation of the ventral tegmental area. J. Neurosci. 1987, 7, 3888–3896. [Google Scholar] [CrossRef]
- Salamone, J.D.; Cousins, M.S.; Snyder, B.J. Behavioral functions of nucleus accumbens dopamine: Empirical and conceptual problems with the anhedonia hypothesis. Neurosci. Biobehav. Rev. 1997, 21, 341–359. [Google Scholar] [CrossRef]
- Tsai, H.C.; Zhang, F.; Adamantidis, A.; Stuber, G.D.; Bonci, A.; de Lecea, L.; Deisseroth, K. Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning. Science 2009, 324, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Torregrossa, M.M.; Kalivas, P.W. Neurotensin in the ventral pallidum increases extracellular gamma-aminobutyric acid and differentially affects cue- and cocaine-primed reinstatement. J. Pharm. Exp. 2008, 325, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Hjelmstad, G.O.; Xia, Y.; Margolis, E.B.; Fields, H.L. Opioid modulation of ventral pallidal afferents to ventral tegmental area neurons. J. Neurosci. 2013, 33, 6454–6459. [Google Scholar] [CrossRef]
- Forcelli, P.A.; West, E.A.; Murnen, A.T.; Malkova, L. Ventral pallidum mediates amygdala-evoked deficits in prepulse inhibition. Behav. Neurosci. 2012, 126, 290–300. [Google Scholar] [CrossRef]
- Chang, C.H.; Grace, A.A. Amygdala-ventral pallidum pathway decreases dopamine activity after chronic mild stress in rats. Biol. Psychiatry 2014, 76, 223–230. [Google Scholar] [CrossRef]
- Huston, J.P.; Hasenöhrl, R.U.; Boix, F.; Gerhardt, P.; Schwarting, R.K. Sequence-specific effects of neurokinin substance P on memory, reinforcement, and brain dopamine activity. Psychopharmacology 1993, 112, 147–162. [Google Scholar] [CrossRef]
- Boix, F.; Sandor, P.; Nogueira, P.J.; Huston, J.P.; Schwarting, R.K. Relationship between dopamine release in nucleus accumbens and place preference induced by substance P injected into the nucleus basalis magnocellularis region. Neuroscience 1995, 64, 1045–1055. [Google Scholar] [CrossRef]
- Studler, J.M.; Kitabgi, P.; Tramu, G.; Herve, D.; Glowinski, J.; Tassin, J.P. Extensive co-localization of neurotensin with dopamine in rat meso-cortico-frontal dopaminergic neurons. Neuropeptides 1988, 11, 95–100. [Google Scholar] [CrossRef]
- Kitabgi, P. Neurotensin modulates dopamine neurotransmission at several levels along brain dopaminergic pathways. Neurochem. Int. 1989, 14, 111–119. [Google Scholar] [CrossRef]
- Bayer, V.E.; Towle, A.C.; Pickel, V.M. Ultrastructural localization of neurotensin-like immunoreactivity within dense core vesicles in perikarya, but not terminals, colocalizing tyrosine hydroxylase in the rat ventral tegmental area. J. Comp. Neurol. 1991, 311, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Sarret, P.; Beaudet, A. Chapter VI Neurotensin receptors in the central nervous system. In Handbook of Chemical Neuroanatomy; Björklund, A., Hökfelt, T., Björklund, A., Hökfelt, T., Quinton, P., Eds.; Peptide Receptors Part II; Elsevier: Amsterdam, The Netherlands, 2002; Volume 20, pp. 323–400. [Google Scholar]
- Richfield, E.K.; Young, A.B.; Penney, J.B. Comparative distribution of dopamine D-1 and D-2 receptors in the basal ganglia of turtles, pigeons, rats, cats, and monkeys. J. Comp. Neurol. 1987, 262, 446–463. [Google Scholar] [CrossRef]
- Fassio, A.; Evans, G.; Grisshammer, R.; Bolam, J.P.; Mimmack, M.; Emson, P.C. Distribution of the neurotensin receptor NTS1 in the rat CNS studied using an amino-terminal directed antibody. Neuropharmacology 2000, 39, 1430–1442. [Google Scholar] [CrossRef]
- Reichard, R.A.; Parsley, K.P.; Subramanian, S.; Stevenson, H.S.; Schwartz, Z.M.; Sura, T.; Zahm, D.S. The lateral preoptic area and ventral pallidum embolden behavior. Brain Struct. Funct. 2019, 224, 1245–1265. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment and Dose | Number of Animals in CPP Test (n) | Number of Animals in EPM Test (n) | ||
|---|---|---|---|---|
| 1st Injection | 2nd Injection | |||
| D1-like antagonist cohorts | vehicle | vehicle | 7 | 11 |
| vehicle | 100 ng NT | 7 | 10 | |
| 1 μg SCH23390 | vehicle | 8 | 12 | |
| 1 μg SCH23390 | 100 ng NT | 9 | 12 | |
| D2-like antagonist cohorts | vehicle | vehicle | 8 | 11 |
| vehicle | 100 ng NT | 7 | 9 | |
| 4 μg sulpiride | vehicle | 7 | 9 | |
| 4 μg sulpiride | 100 ng NT | 8 | 10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ollmann, T.; Lénárd, L.; Péczely, L.; Berta, B.; Kertes, E.; Zagorácz, O.; Hormay, E.; László, K.; Szabó, Á.; Gálosi, R.; et al. Effect of D1- and D2-like Dopamine Receptor Antagonists on the Rewarding and Anxiolytic Effects of Neurotensin in the Ventral Pallidum. Biomedicines 2022, 10, 2104. https://doi.org/10.3390/biomedicines10092104
Ollmann T, Lénárd L, Péczely L, Berta B, Kertes E, Zagorácz O, Hormay E, László K, Szabó Á, Gálosi R, et al. Effect of D1- and D2-like Dopamine Receptor Antagonists on the Rewarding and Anxiolytic Effects of Neurotensin in the Ventral Pallidum. Biomedicines. 2022; 10(9):2104. https://doi.org/10.3390/biomedicines10092104
Chicago/Turabian StyleOllmann, Tamás, László Lénárd, László Péczely, Beáta Berta, Erika Kertes, Olga Zagorácz, Edina Hormay, Kristóf László, Ádám Szabó, Rita Gálosi, and et al. 2022. "Effect of D1- and D2-like Dopamine Receptor Antagonists on the Rewarding and Anxiolytic Effects of Neurotensin in the Ventral Pallidum" Biomedicines 10, no. 9: 2104. https://doi.org/10.3390/biomedicines10092104
APA StyleOllmann, T., Lénárd, L., Péczely, L., Berta, B., Kertes, E., Zagorácz, O., Hormay, E., László, K., Szabó, Á., Gálosi, R., Karádi, Z., & Kállai, V. (2022). Effect of D1- and D2-like Dopamine Receptor Antagonists on the Rewarding and Anxiolytic Effects of Neurotensin in the Ventral Pallidum. Biomedicines, 10(9), 2104. https://doi.org/10.3390/biomedicines10092104