
A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment

 ,
,  , , , and
, , , and
Abstract
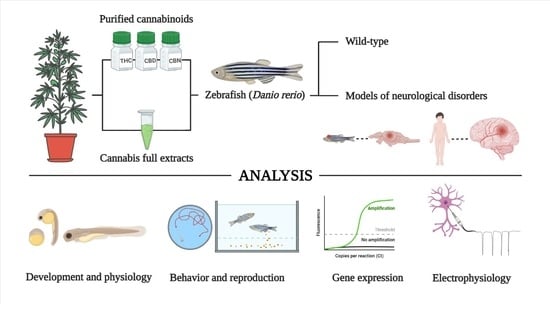
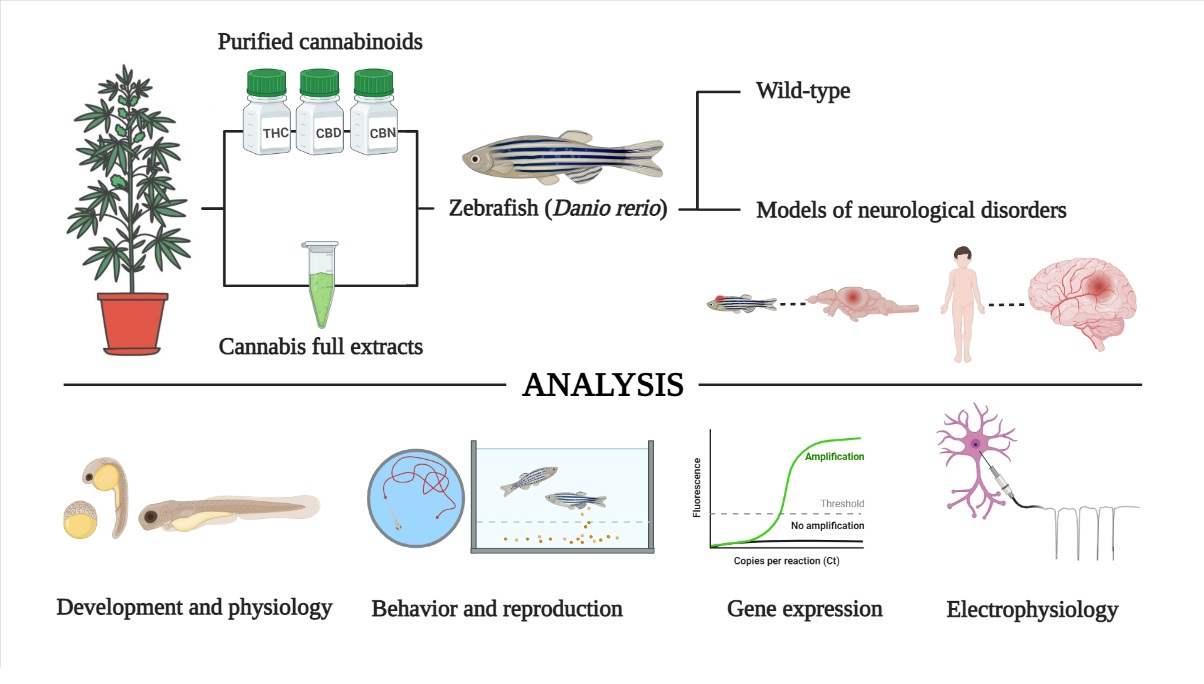
:
1. Introduction
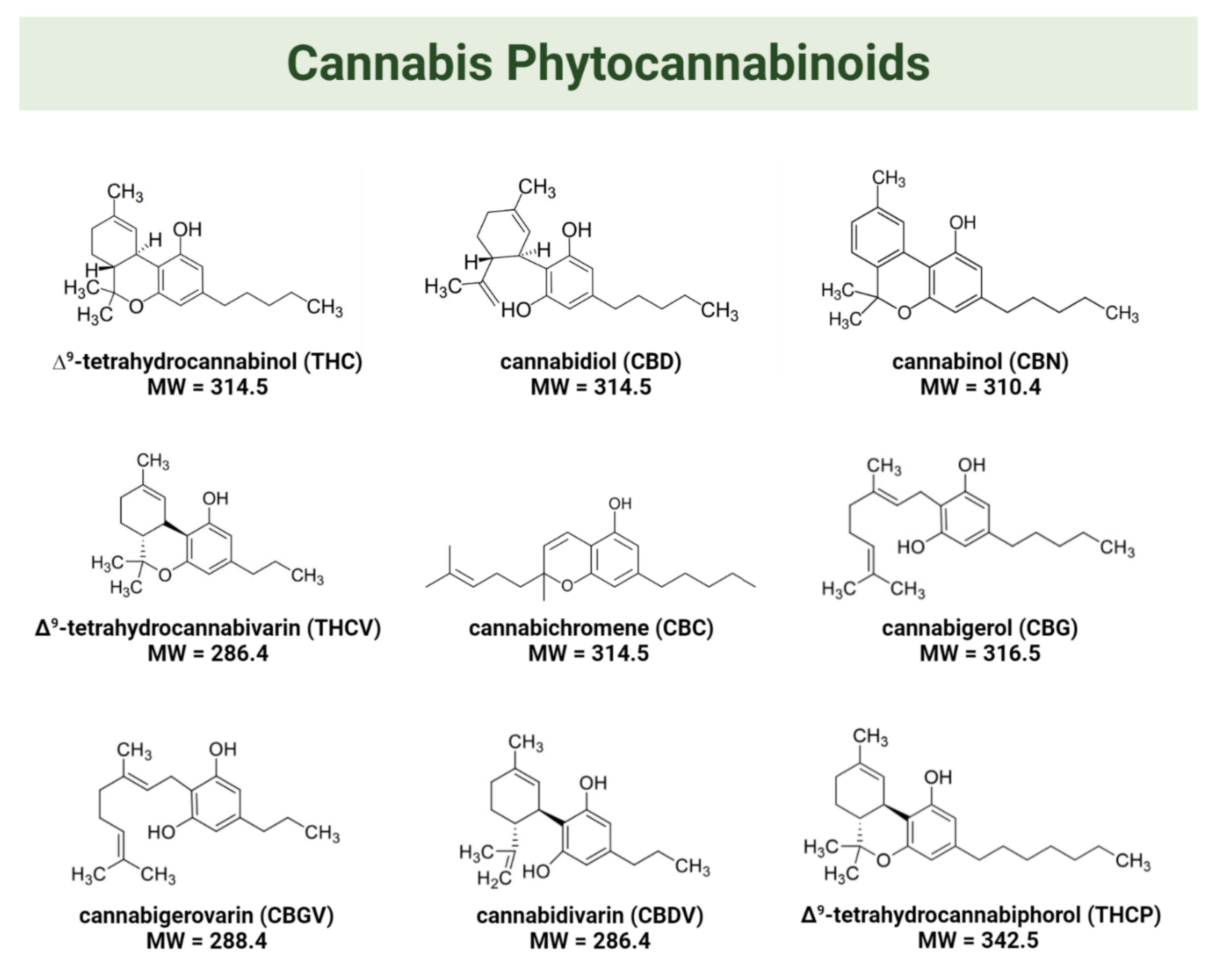
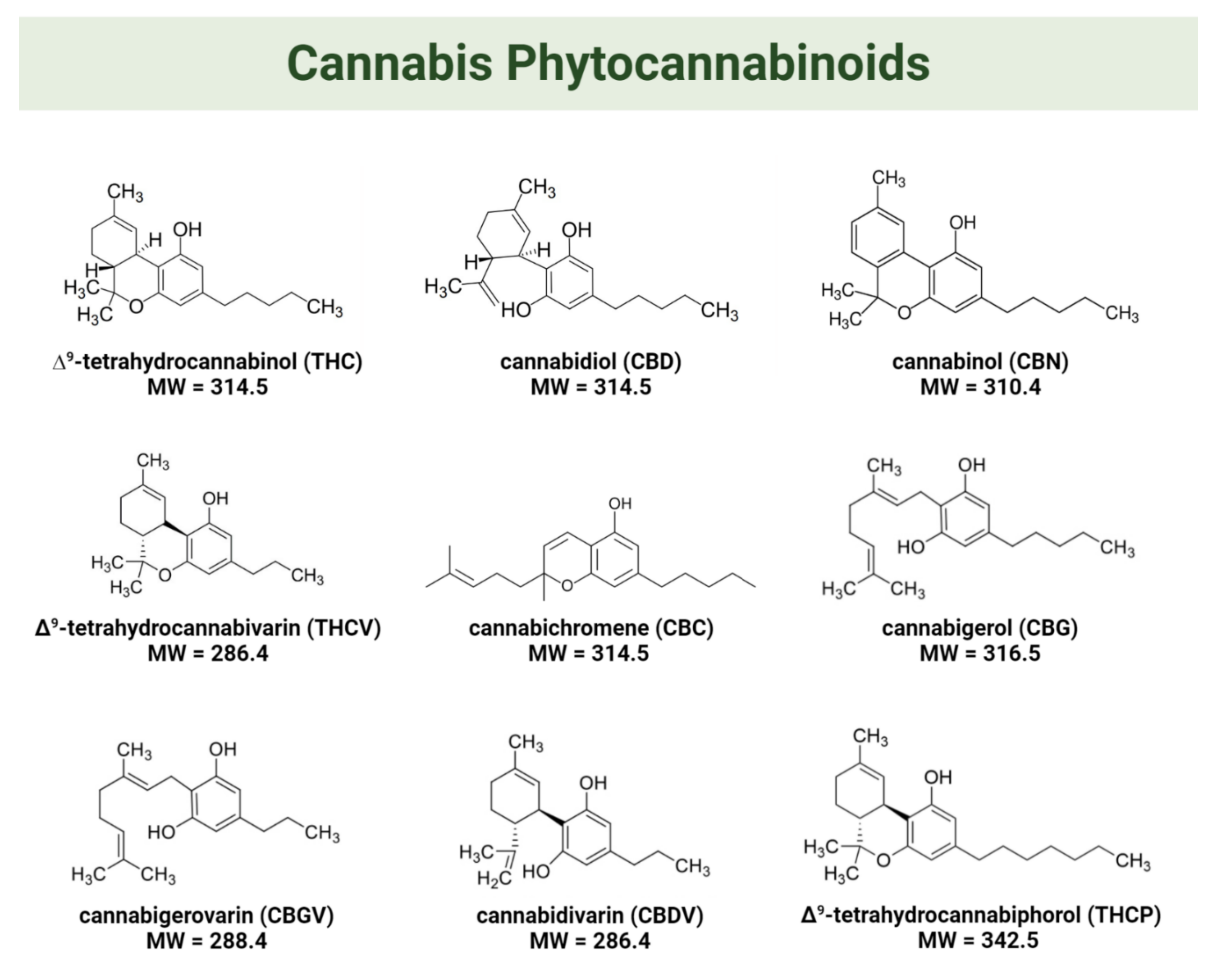
1.1. The Endocannabinoid System
1.2. Zebrafish as a Model System to Test the Bioactivity of Cannabinoids
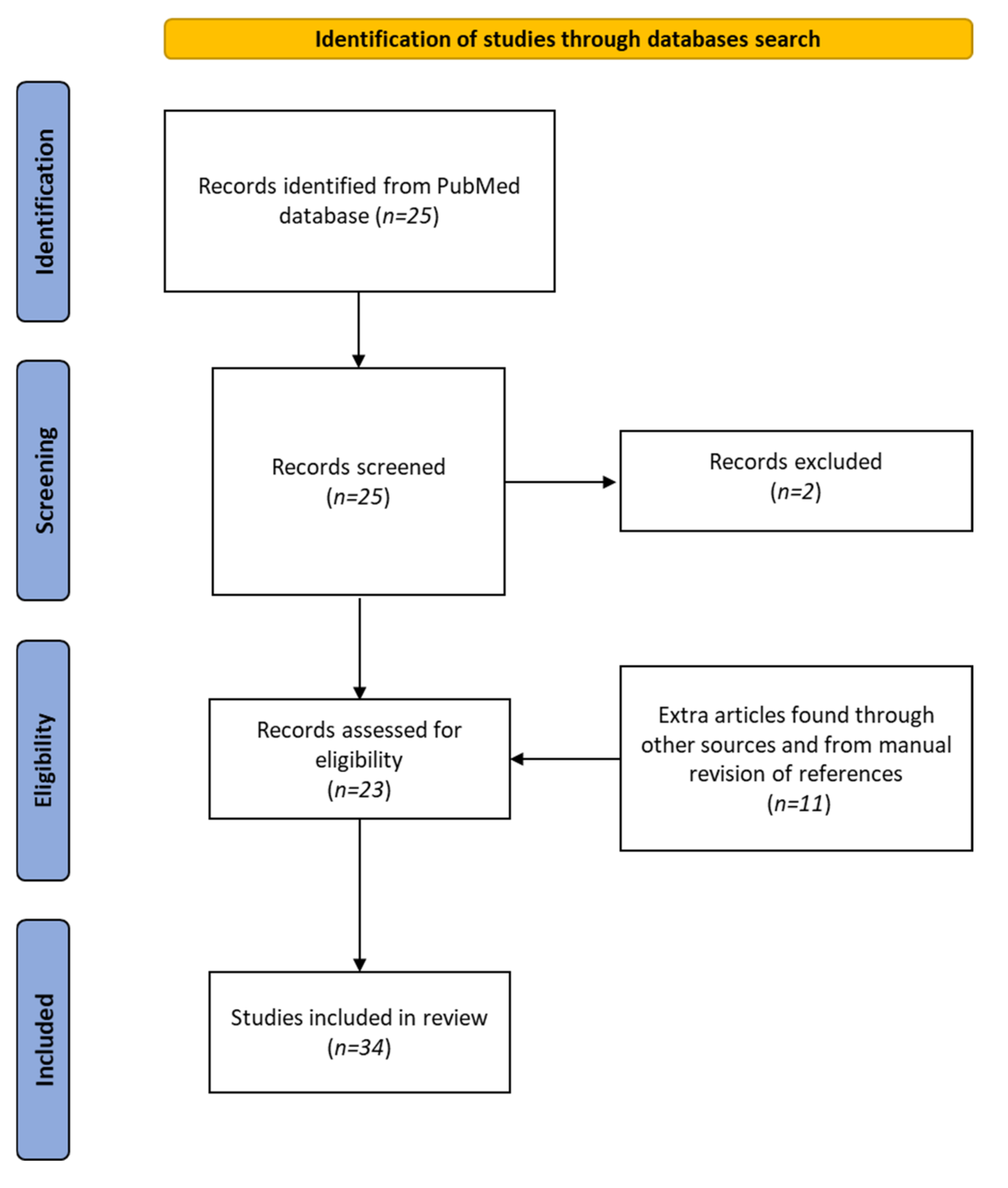
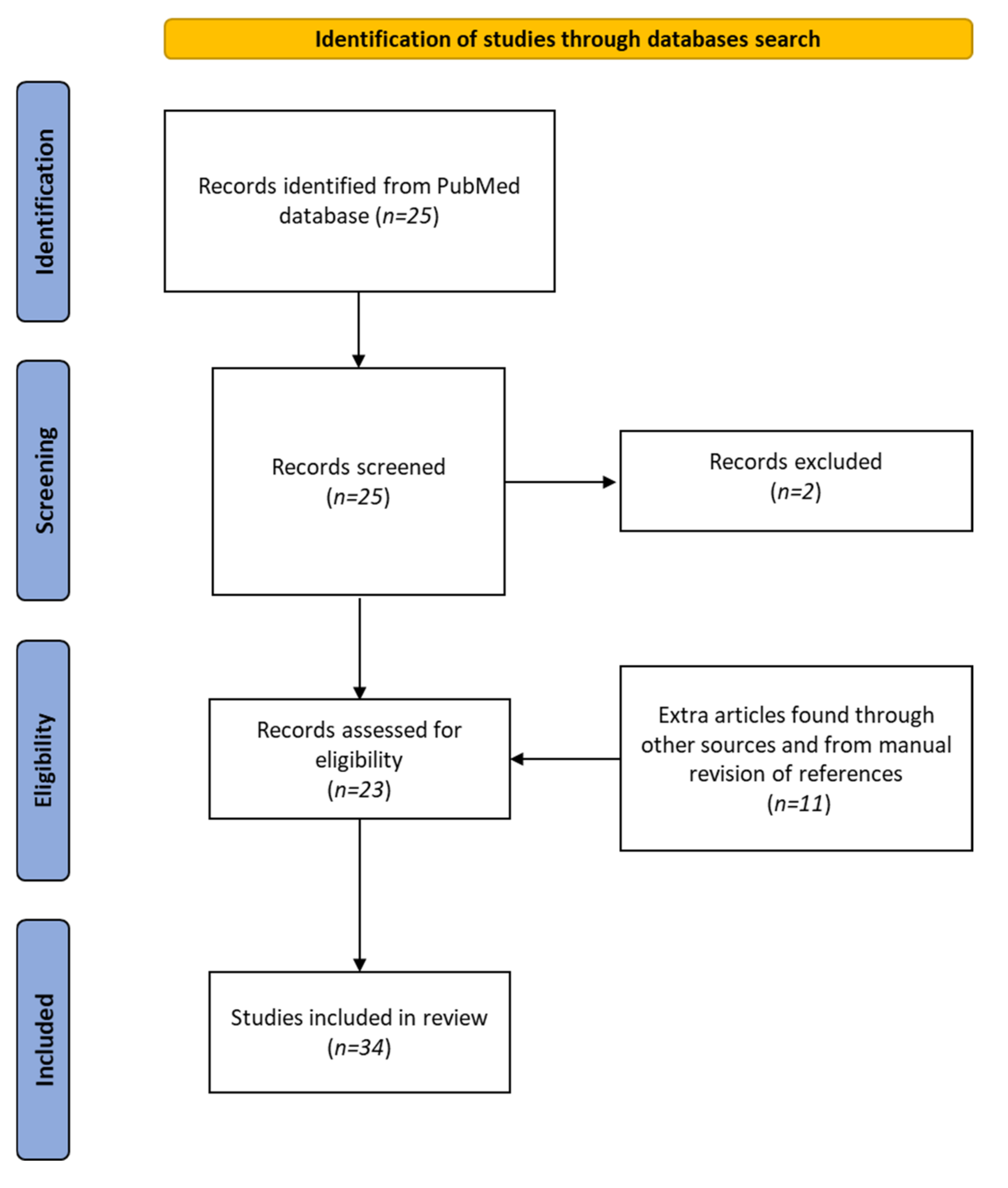
2. Materials and Methods
Data Sources and Searches
3. Discussion
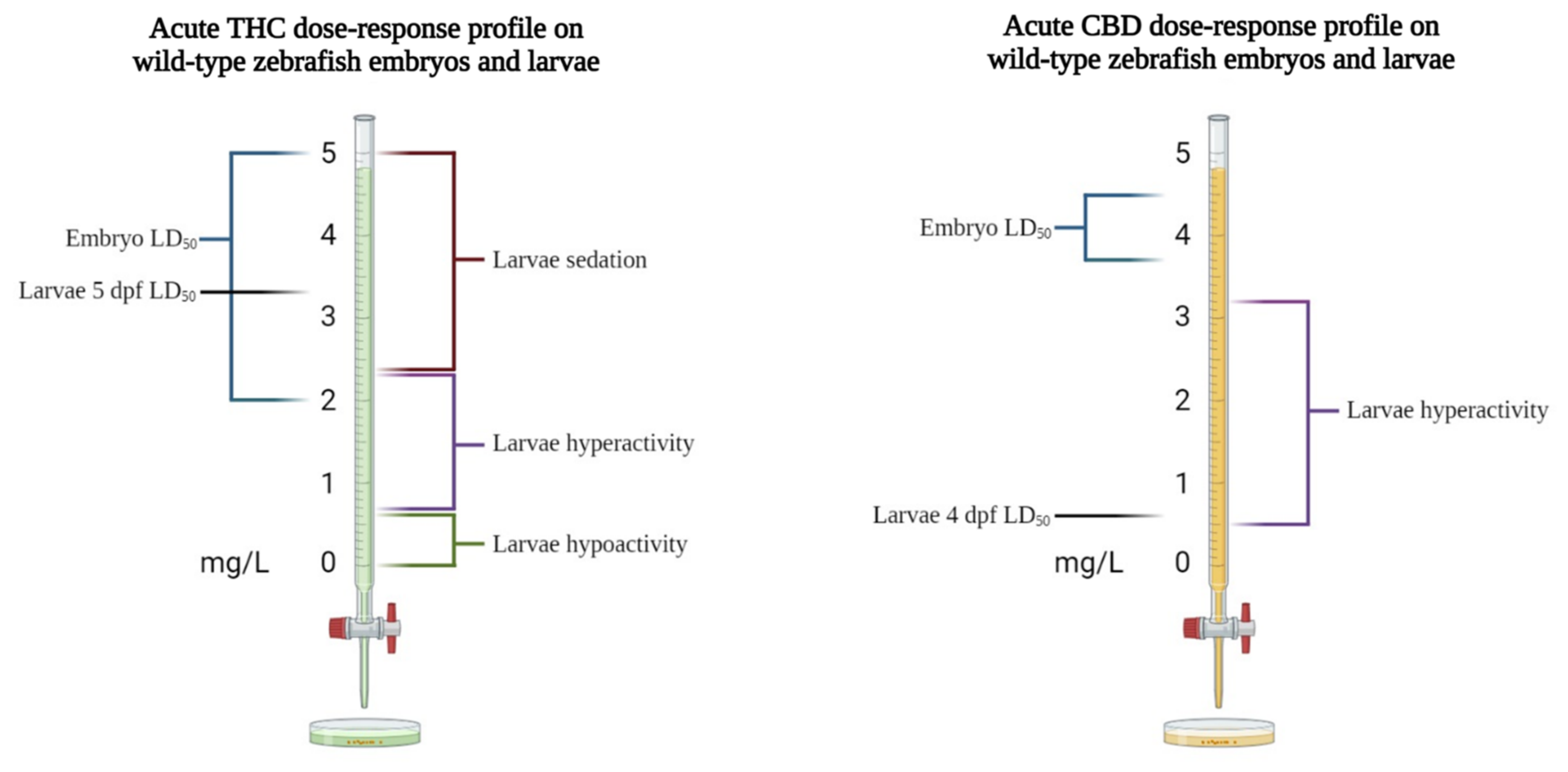
3.1. Effects of Phytocannabinoids in Wild-Type Zebrafish
3.2. Effects of Phytocannabinoids in Zebrafish Models of Neurological Disorders

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studies Carried Out on Embryos/Larvae | ||||||
|---|---|---|---|---|---|---|
| Compound Concentration and Exposure | Strain | Age | Nonbehavioral Analysis | Behavioral Analysis | Results | References |
| THC (0.016, 0.031, 0.156, 0.469, and 0.625 mg/L) and CBD (0.225, 0.3, 0.525, 0.75, and 1.125 mg/L). Acute exposure (3–4 min before analysis). | AB/TU 1 | 5 dpf | / | VMR test: 150 min of light followed by a 5 min dark–light cycle for 30 min. | Locomotion: THC: decreased locomotor activity at all concentrations tested; CBD: increased locomotor activity at concentrations above 0.525 mg/L. | Achenbach et al., 2018 [31] |
| THC (2, 4, 6, 8, and 10 mg/L) and CBD (1, 2, 3, and 4 mg/L). Acute exposure (5 h during gastrulation stage). | TL 2 | 5 dpf | Survival, hatching rate, morphology, basal heart rate, and synaptic activity at neuromuscular junctions | Auditory/mechanical escape response test | Survival: Embryos exposed to 8–10 mg/L THC and 3–4 mg/L CBD had reduced survival rates. Hatching rate: Reduced with both THC and CBD, at all concentrations tested. Morphology and basal heart rate: Dose-dependent reductions in both body length and heart rate. Synaptic activity: Reduced with 6 mg/L THC and 3 mg/L CBD. Escape response: No reduction in touch response but decreases in sound response with 6 mg/L of THC and 3 mg/L of CBD. | Ahmed et al., 2018 [10] |
| THC (0.3–3.4 mg/L), CP 55,940 (2.25–18 mg/L), and WIN 55,212-2 (0.3–1.8 mg/L). Acute exposure (1, 4, and 12 min before analysis) and chronic exposure (96 h: from 24 to 120 hpf). | / | 5 dpf | LD50 determination and morphology | VMR test: 4 min of light followed by 4 min of dark. | LD50: A total of 3.37 mg/L for THC, 1.8 mg/L for WIN 55,212-2, and 16.92 mg/L for CP 55,940. Morphology: THC caused malformations at all concentrations tested, while CP 55,940 and WIN 55,212-2 did not significantly increase the frequency of malformations. Locomotion: In acute exposure conditions, a biphasic response (stimulation at low concentrations and suppression at high concentrations) was observed; in chronic exposure, only 1.2 mg/L THC had a significant effect (increased distance traveled). | Akhtar et al., 2013 [15] |
| THC (6 mg/L). Acute exposure (5 h during gastrulation stage). | TL 2 | 2 and 5 dpf | Morphology of Mauthner cells and immunohistochemical analysis of the trunk muscles | Mechanical escape response test at 2 dpf and VMR test at 5 dpf (60 min). | Morphology: THC exposure reduced axonal diameter of Mauthner cells. Escape response: No reduction in C-bend response rate, but C-bend angle was increased in THC-treated embryos. Immunohistochemistry: White and red muscle fibers appeared thinner and slightly disorganized in THC-treated embryos. Locomotion: THC impaired locomotor performance. | Amin et al., 2020 [75] |
| Dihydrophenanthrene derivative (1–5 μM). Acute exposure (3–4 min before analysis). | AB/TU 1 | 5 dpf | / | VMR test: 150 min of light followed by 5-min dark–light cycles (for 30 min). | Locomotion: Locomotor activity was increased at concentrations from 2.5 to 5 μM during the first 50 min, but normally increased larval locomotor activity was reduced during the dark phases. | Banskota et al., 2021 [12] |
| THC (0.3125, 0.625, 1.25, 2.5, 5 mg/) and CBD (0.075, 0.15, 0.3, 0.6, 1.2 mg/L). Chronic exposure (94 h: from 2 to 96 hpf). | Tg(fli1:egfp) | 4 dpf | Toxicity and morphology | Touch response and VMR test: 10 min light–dark cycles (for 30 min). | Morphology: THC and CBD displayed concentration-dependent morphological toxicities. Locomotion: Larvae exposed to 0.3 mg/L THC, or 0.07 mg/L CBD, exhibited a significantly increased duration of movement during dark phases compared with control. By contrast, 1.25 mg/L THC and 0.1–0.3 mg/L CBD significantly reduced duration of movement compared with control. | Carty et al., 2018 [86] |
| THC (0.024, 0.12, and 0.6 mg/L) and CBD (0.006, 0.03, and 0.15 mg/L). Chronic exposure (90 h: from 6 to 96 hpf). | Tg(fli1:egfp) | 4 dpf | Survival and fertility rate | VMR test: 10 min dark–light cycles (for 30 min). | Survival and fertility: Not affected by treatments. Locomotion: Hypoactivity observed in larvae exposed to the lowest concentration of THC, and only during the dark phases. | Carty et al., 2019 [32] |
| CBN (0.25, 0.75, 1.0, 1.125, 1.2, 1.25, and 2 mg/L). Chronic exposure (96 h: from 24 to 120 hpf). | AB | 5–7 dpf | Survival, morphology, LD50, and basal heart rate | VMR test: 10 min dark–light cycles (for 30 min). Mechanical escape response test: 2 min in dark conditions. | LD50: 1.12 mg/L. Morphology: Concentrations higher than 0.75 mg/L led to malformations. Basal heart rate: At concentrations higher than 0.75 mg/l, heart rate decreased significantly, exhibiting characteristic bradycardia. Locomotion: Distance was significantly reduced as CBN concentration increased in both dark and light conditions; velocity increased with increasing CBN concentration under dark conditions and decreased under light conditions. Escape response: No differences. | Chousidis et al., 2020 [30] |
| Acute exposure to acetic acid solution (0.1–0.5%) (nociception stimulus), followed by exposure to THC (0.15625 mg/L) or CBD (0.15 mg/L). Acute exposure (2 h before analysis). | AB/TU 1 | 5 dpf | / | VMR test: 2.5 h exposure to light followed by 5 min dark–light cycles (for 30 min). | Locomotion: THC-exposed larvae showed reduced activity compared with both acetic acid-exposed and control-group larvae, while CBD elevated the activity level of the larvae compared with acetic acid-exposed group. There was no significant reduction in the light–dark transition response in any of the test groups. | Ellis et al., 2018 [25] |
| 20 synthetic cannabinoids (1, 10, and 100 μM). Acute exposure (20 min before analysis). | scn1lab−/− | 5 dpf | Electrophysiology | VMR test: 10 min | Electrophysiology: Five synthetic cannabinoids decreased the frequency of spontaneous epileptiform events. Locomotion: Five synthetic cannabinoids decreased seizure-like swims in a concentration-dependent manner. | Griffin et al., 2020 [17] |
| CBD (0.5, 1, 5, and 10 mg/L) and WIN55,212–2 (0.5, 1, 5, and 10 mg/L). Acute exposure (30 min). | / | 4–6 dpf | / | VMR test: 15 min dark–light cycle for 180 min. | Locomotion: CBD reduced the movement velocity and total distance moved. Moreover, CBD at 10 mg/L attenuated the responses of larvae exposed to darkness. No differences were detected between the control and CBD-treated groups after 24 h in fresh water. Fish treated with WIN55,212–2 at 0.5 and 1 mg/L showed virtually no activity, even in darkness, whereas a concentration of 10 mg/L induced mortality. A 24 h period in fresh water had the effect of reversing most of the drug-induced immobilization, even in the WIN55,212-2-treated groups. Finally, treatment with CBD attenuated WIN55,212-2-induced abnormal immobilization, whereas equivalent doses of CBD and WIN55,212–2 produced a mixed response. | Hasumi et al., 2020 [111] |
| THCV (0.286 and 0.859 mg/L) and THCV−OH (0.859 mg/L). Chronic exposure (5 days: from 3 to 8 dpf). | / | 8 dpf | Number of ossified vertebral centers | / | Morphology: THCV reduced the number of ossified vertebral centers, whereas THCV−OH increased it. | Janssens et al., 2018 [37] |
| THC (2, 3, 4, and 6 μg/mL) and CBD (1.5, 2, and 3 mg/L). Acute exposure (9.5 h: from 0.5 to 10 hpf). | CaMPARI transgenic/Casper | 4–5 dpf | Neural activity | VMR test: 60 min | Neural activity: Reduced in embryos exposed to 2–3 mg/L of CBD and 4–6 mg/L of THC. Locomotion: Reduced in embryos exposed to 3 mg/L CBD and 6 mg/L THC. | Kanyo et al., 2021 [3] |
| Whole-plant cannabis extract. Chronic exposure (96 h of exposition starting at 24 hpf). | AB | 5 dpf | Gene expression | VMR test: 150 min of light followed by 5 min dark–light cycles (for 30 min). | Locomotion: During both the first 150 min of light and the remaining 30 min of light–dark cycles, larvae treated with cannabis at the highest dose (200 µL) showed increased locomotor activity. Gene expression: Both zebrafish cannabinoid receptors (cnr1 and cnr2) were overexpressed at the highest dose (200 µL). | Licitra et al., 2021 [91] |
| WIN55,212-2 (0.014–3.412 mg/L) and CP55,940 (0.188–3.013 mg/L), and specific cnr2 agonists HU-910 and JWH-133. Acute exposure (1 h before analysis). | AB/TU 1 and cnr1−/− | 5 dpf | / | VMR test: 4 min of light, 4 min of dark, and 30 min of light. | Locomotion: WIN55,212-2 and CP55,940 produced a dose-dependent reduction in locomotor activity in both the light and dark phases. HU-910 and JWH-133 have no effect on locomotion. In the cnr1−/− larvae, no inhibitory effect of WIN55,212-2 or CP55,940 on the average swimming velocity was found. The cnr1 antagonist AM251 did not affect locomotor activity, but blocked the effect of WIN55,212-2, which suggests that endocannabinoids are not active in regulating locomotor activity in zebrafish larvae at 5 dpf. | Luchtenburg et al., 2019 [87] |
| Whole-plant cannabis extracts. Acute exposure (2 h). | AB/TU 1 | 5 dpf | / | VMR test: 90 min of light followed by 5 min dark–light cycles (for 30 min). | Locomotion: During the first 30 min of light, exposure to high THC extracts led to reduced activity at 0.25 mg/L and higher activity at 1 and 2 mg/L. Instead, exposure to high CBD extracts led to hyperactivity at 0.5 and 1 mg/L. During the final 30 min of the light cycle, high THC extracts significantly decreased activity at all concentrations tested, while high CBD extracts led to a reduction in activity only at 2 mg/L. During light–dark transitions, the locomotor response was abolished at 2 mg/L (in the dark phase). | Nixon et al., 2021 [92] |
| THC (0.156–2.1875 mg/L), CBD (0.3–2.1 mg/L), and THC–CBD combination. Acute exposure (1 h before analysis). | AB/TU + PTZ and GABRA1 knock-out | 5 dpf | / | VMR test: 30 min | Locomotion: THC, CBD, and their combination reduced PTZ-induced neuro-hyperactivity and alleviated GABRA1−/− seizures. | Samarut et al., 2019 [119] |
| THC (1, 2, 5, and 10 mg/L). Acute exposure (19.5 h: from 4.5 to 24 hpf). | / | 1–9 dpf | Survival and morphology | Tail twitches | Survival: After 24 h of exposure, no effects of THC on survival were recorded, but after between 2 and 9 days of exposure, survival was greatly reduced. Morphology: At levels above 2 mg/L THC, larvae showed curved trunks and/or bulbous-tipped tails. Coiling: At 26–28 hpf, the number of twitches following exposure to 5 or 10 mg/L THC was significantly reduced. | Thomas, 1975 [104] |
| CBD (0.075–0.3 mg/L), THC (0.3125–1.25 mg/L), CBDV (0.072, 0.172, 0.286, and 1.146 mg/L), CBN (0.078, 0.186, 0.310, and 1.242 mg/L), or LN (0.107, 0.256, 0.427, and 1.707 mg/L). Acute exposure (24 h: from 120 to 144 hpf). | scn1lab−/− and WT (scn1lab+/+ or scn1lab+/− + PTZ) | 6 dpf | Morphology | VMR test: 15 min | Morphology: CBN and CBDV led to a high incidence of deformities. Locomotion: THC (1.25 mg/L) significantly reduced total distance traveled. In wild-type specimens, PTZ-induced hyperlocomotion was significantly reduced following exposure to CBD or THC, but no changes were observed following CBDV, CBN, or LN exposure. In the scn1lab−/− mutants, the total distance traveled was significantly reduced following exposure to CBD (0.15 mg/L), THC (0.3125 mg/L), CBN, and LN. | Thornton et al., 2020 [14] |
| O-1602 and abnormal CBD (up to 3.14 mg/L). Chronic exposure (3 days: from 2 to 5 dpf). | AB/TU 1 | 5 dpf | Toxicity and antitumor effects | / | Toxicity: Concentrations greater than 2.5 μM led to higher levels of toxicity to the larvae. Antitumor: Both atypical cannabinoids significantly reduced the presence of injected cancer cells in the zebrafish larvae, by approximately 50%. | Tomko et al., 2019 [24] |
| CBD (5, 20, 70, 150, and 300 μg/L). Chronic exposure (4 days: from 0 to 4 dpf). | / | 3–4 dpf | Toxicity and morphological analysis | Motor activity calculated as number of active events for 3 min. | Toxicity and morphology: CBD did not show significant differences in the morphological parameters at any dose, but at the highest concentration, CBD significantly delayed the hatching time of embryos. Locomotion: Above 20 μg/L, CBD increases the motor activity at 24 hpf, but not at 48 hpf. | Valim Brigante et al., 2018 [52] |
| Studies carried out in adult fish | ||||||
| THC (0.024, 0.12, and 0.6 mg/L) and CBD (0.006, 0.03, and 0.15 mg/L). Chronic exposure (90 h: from 6 to 96 hpf). | Tg(fli1:egfp) | 12–18 months | Reproductive parameters | Open field test: 6 min | Reproduction: Reduced fecundity in adults exposed to CBD (0.15 mg/L) and THC (0.024 and 0.12 mg/L). Locomotion: No significant effects. | Carty et al., 2019 [32] |
| WIN55,212-2 (0.5, 5, or 50 mg/L). Acute aqueous exposure (10 min) and dietary exposure (1 μg/day/fish for 1 week). | / | / | / | Light–dark cross-maze test: 5 min | Locomotion: Altered behavioral anxiolytic responses and reduced locomotor activity at all tested doses. A 1-week dietary exposure promoted zebrafish exploration. | Connors et al., 2014. [125] |
| THC (0.0125, 0.3125, and 0.625 mg/L). Acute exposure (2 min prior to starting analysis). | EK | 9–12 months | / | Locomotion: 20 min | Locomotion: THC (0.3125 mg/L) reduced velocity and induced repetitive swimming patterns. | Dahlén et al., 2021 [2] |
| CBD (40 mg/L). Acute exposure (30 min). | / | 6 months | Gene expression | Locomotion: 3 min | Locomotion: CBD reduced distance traveled and velocity. Gene expression: CBD activated genes encoding proinflammatory cytokines (il-1b and il-17a/f2). | Jensen et al., 2018 [7] |
| CBD (0.1 0.5, 5.0, or 10 mg/kg) via intraperitoneal injection (1 h before analysis). | TU | 4 months | / | Locomotion, anxiety, aggressive behavior (1 min), and social interaction (10 min). Memory-assessment task. | Locomotion: Not affected. Anxiety: Inverted U-shaped dose–response curve with 0.5 mg/kg reducing the anxiety. Aggressive behavior and social interaction: Not affected by 5 mg/kg CBD. Memory: CBD (5 mg/kg) caused memory impairment. | Nazario et al., 2015 [48] |
| THC (0.024, 0.12, and 0.6 mg/L). Chronic exposure (90 h: from 6 to 96 hpf). | Tg(fli1:egfp) | 12–30 months | Survival, reproductive and growth parameters, and gene expression | Open field: 5 min | Survival: Increased at 0.024 mg/L THC in male fish. A significant reduction in survival of F1 THC-treated male fish by 30 months of age. Reproduction: THC exposition did not significantly alter sperm production, and exposure to 0.024 mg/L THC improved egg production in aged females; the resulting offspring at 96 hpf showed similar survival to both young and aged control fish. The F1 fish parentally exposed to 0.6 mg/L THC were completely unable to reproduce, unlike the aged controls. Growth: No difference in body length or mass was observed in male fish exposed to vehicle or THC, while exposure to the lowest concentration of THC (0.024 mg/L) resulted in significant reductions in mass in advanced aged females. Locomotion: With the exception of increased mobility in 0.12 mg/L-THC-exposed males, early-life treatment with THC did not affect locomotor abilities in 30-month-old male or female fish. Gene expression: Significant reductions in tnf-α and il-1β, and increases in il-6, pparα and pparγ. | Pandelides et al., 2020 [27] |
| THC (0.03125 mg/L). Acute exposure (1 h). | / | 12 months | / | Color-discrimination learning and spatial-cognition task. | Color-discrimination learning: THC administration did not lead to any observable effect on color-discrimination learning. Spatial cognition: Impaired. | Ruhl et al., 2014 [113] |
4. Pointers on Behavioral Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive eethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 5, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Dahlén, A.; Zarei, M.; Melgoza, A.; Wagle, M.; Guo, S. THC-induced behavioral stereotypy in zebrafish as a model of psychosis-like behavior. Sci. Rep. 2021, 11, 15693. [Google Scholar] [CrossRef]
- Kanyo, R.; Amin, M.R.; Locskai, L.F.; Bouvier, D.D.; Olthuis, A.M.; Allison, W.T.; Ali, D.W. Medium-throughput zebrafish optogenetic platform identifies deficits in subsequent neural activity following brief early exposure to cannabidiol and Δ9-tetrahydrocannabinol. Sci. Rep. 2021, 11, 11515. [Google Scholar] [CrossRef] [PubMed]
- Lazarjani, M.P.; Young, O.; Kebede, L.; Seyfoddin, A. Processing and extraction methods of medicinal cannabis: A narrative review. J. Cannabis Res. 2021, 3, 32. [Google Scholar] [CrossRef] [PubMed]
- Chiarlone, A.; Börner, C.; Martín-Gómez, L.; Jiménez-González, A.; García-Concejo, A.; García-Bermejo, M.L.; Lorente, M.; Blázquez, C.; García-Taboada, E.; de Haro, A.; et al. MicroRNA let-7d is a target of cannabinoid CB1 receptor and controls cannabinoid signaling. Neuropharmacology 2016, 108, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, H.; Qureshi, I.A.; Jahan, N.; Went, T.R.; Sultan, W.; Sapkota, A.; Alfonso, M. A Systematic Review of Fibromyalgia and Recent Advancements in Treatment: Is Medicinal Cannabis a New Hope? Cureus 2021, 13, e17332. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.M.; Korbut, R.; Kania, P.W.; Buchmann, K. Cannabidiol effects on behaviour and immune gene expression in zebrafish (Danio rerio). PLoS ONE 2018, 31, e0200016. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.D.; Kim, J.H.; Cheslack-Postava, K.; Weinberger, A.H.; Wu, M.; Wyka, K.; Kattan, M. Trends in cannabis use among adults with children in the home in the United States, 2004–2017: Impact of state-level legalization for recreational and medical use. Addiction 2021, 116, 2770–2778. [Google Scholar] [CrossRef]
- Crippa, J.A.; Derenusson, G.N.; Ferrari, T.B.; Wichert-Ana, L.; Duran, F.L.; Martin-Santos, R.; Simões, M.V.; Bhattacharyya, S.; Fusar-Poli, P.; Atakan, Z.; et al. Neural basis of anxiolytic effects of cannabidiol (CBD) in generalized social anxiety disorder: A preliminary report. J. Psychopharmacol. 2011, 25, 121–130. [Google Scholar] [CrossRef]
- Ahmed, K.T.; Amin, M.R.; Shah, P.; Ali, D.W. Motor neuron development in zebrafish is altered by brief (5-hr) exposures to THC (∆9-tetrahydrocannabinol) or CBD (cannabidiol) during gastrulation. Sci. Rep. 2018, 8, 10518. [Google Scholar] [CrossRef] [Green Version]
- Omare, M.O.; Kibet, J.K.; Cherutoi, J.K.; Kengara, F.O. Current Trends in the Use of Cannabis sativa: Beyond Recreational and Medicinal Applications. Open Access Libr. 2021, 8, 6. [Google Scholar] [CrossRef]
- Banskota, A.H.; Stefanova, R.; Hui, J.P.M.; Berrué, F.; Achenbach, J.C.; Ellis, L. 9,10-Dihydro-5-hydroxy-2,3,6-trimethoxyphenanthrene-1,4-dione: A new dihydrophenanthrene from commercial cannabis and its effect on zebrafish larval behaviour. Nat. Prod. Res. 2021, 36, 3553–3558. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Gong, Z.; Kelly, B.C. Metabolomic profiling of zebrafish (Danio rerio) embryos exposed to the antibacterial agent triclosan. Environ. Toxicol. Chem. 2019, 38, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Thornton, C.; Dickson, K.E.; Carty, D.R.; Ashpole, N.M.; Willett, K.L. Cannabis constituents reduce seizure behavior in chemically-induced and scn1a-mutant zebrafish. Epilepsy Behav. 2020, 110, 107152. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.T.; Ali, S.; Rashidi, H.; van der Kooy, F.; Verpoorte, R.; Richardson, M.K. Developmental effects of cannabinoids on zebrafish larvae. Zebrafish 2013, 10, 283–293. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Trial of cannabidiol for drug-resistant seizures in the dravet syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef] [Green Version]
- Griffin, A.; Anvar, M.; Hamling, K.; Baraban, S.C. Phenotype-Based Screening of Synthetic Cannabinoids in a Dravet Syndrome Zebrafish Model. Front. Pharmacol. 2020, 11, 464. [Google Scholar] [CrossRef] [Green Version]
- Kerr, A.; Walston, V.; Wong, V.S.S.; Kellogg, M.; Ernst, L. Marijuana use among patients with epilepsy at a tertiary care center. Epilepsy Behav. 2019, 97, 144–148. [Google Scholar] [CrossRef]
- Sagdeo, A.; Askari, A.; Ball, P.; Morrissey, H. Exploring the efficacy and safety of cannabis in the management of fibromyalgia. Int. J. Curr. Pharm. Rev. Res. 2022, 14, 27–30. [Google Scholar] [CrossRef]
- Kosiba, J.D.; Maisto, S.A.; Ditre, J.W. Patient-reported use of medical cannabis for pain, anxiety, and depression symptoms: Systematic review and metaanalysis. Soc. Sci. Med. 2019, 233, 181–192. [Google Scholar] [CrossRef]
- Hamilton, I.; Monaghan, M. Cannabis and psychosis: Are we any closer to understanding the relationship. Curr. Psychiatr. Rep. 2019, 21, 21–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehnke, K.F.; Scott, J.R.; Litinas, E.; Sisley, S.; Williams, D.A.; Clauw, D.J. Pills to pot: Observational analyses of cannabis substitution among medical cannabis users with chronic pain. J. Pain 2019, 20, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Guzmàn, M. Cannabis for the management of cancer symptoms: THC version 2.0? Cannabis Cannabinoid. Res. 2018, 3, 117–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomko, A.; O’Leary, L.; Trask, H.; Achenbach, J.C.; Hall, S.R.; Goralski, K.B.; Ellis, L.D.; Dupré, D.J. Antitumor Activity of Abnormal Cannabidiol and Its Analog O-1602 in Taxol-Resistant Preclinical Models of Breast Cancer. Front. Pharmacol. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Ellis, L.D.; Berrue, F.; Morash, M.; Achenbach, J.C.; Hill, J.; McDougall, J.J. Comparison of cannabinoids with known analgesics using a novel high throughput zebrafish larval model of nociception. Behav. Brain Res. 2018, 337, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Schomig, E.; Leweke, F.M. Acute and chronic cannabinoid treatment differentially affects recognition memory and social behavior in pubertal and adult rats. Addict. Biol. 2008, 13, 345–357. [Google Scholar] [CrossRef]
- Pandelides, Z.; Thornton, C.; Lovitt, K.G.; Faruque, A.S.; Whitehead, A.P.; Willett, K.L.; Ashpole, N.M. Developmental exposure to Δ9-tetrahydrocannabinol (THC) causes biphasic effects on longevity, inflammation, and reproduction in aged zebrafish (Danio rerio). Geroscience 2020, 42, 923–926. [Google Scholar] [CrossRef]
- Carlini, B.H.; Garrett, S.B.; Carter, G.T. Medicinal Cannabis: A Survey Among Health Care Providers in Washington State. Am. J. Hosp. Palliat. Care 2017, 34, 85–91. [Google Scholar] [CrossRef]
- Lopez-Quintero, C.; Hasin, D.S.; de Los Cobos, J.P.; Pines, A.; Wang, S.; Grant, B.F.; Blanco, C. Probability and predictors of remission from life-time nicotine, alcohol, cannabis or cocaine dependence: Results from the National Epidemiologic Survey on Alcohol and Related Conditions. Addiction 2011, 106, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Chousidis, I.; Chatzimitakos, T.; Leonardos, D.; Filiou, M.D.; Stalikas, C.D.; Leonardos, I.D. Cannabinol in the spotlight: Toxicometabolomic study and behavioral analysis of zebrafish embryos exposed to the unknown cannabinoid. Chemosphere 2020, 252, 126417. [Google Scholar] [CrossRef]
- Achenbach, J.C.; Hill, J.; Hui, J.P.M.; Morash, M.G.; Berrue, F.; Ellis, L.D. Analysis of the Uptake, Metabolism, and Behavioral Effects of Cannabinoids on Zebrafish Larvae. Zebrafish 2018, 15, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Carty, D.R.; Miller, Z.S.; Thornton, C.; Pandelides, Z.; Kutchma, M.L.; Willett, K.L. Multigenerational consequences of early-life cannabinoid exposure in zebrafish. Toxicol. Appl. Pharmacol. 2019, 364, 133–143. [Google Scholar] [CrossRef]
- Borgelt, L.M.; Franson, K.L.; Nussbaum, A.M.; Wang, G.S. The pharmacologic and clinical effects of medical cannabis. Pharmacotherapy 2013, 33, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Pisanti, S.; Malfitano, A.M.; Ciaglia, E.; Lamberti, A.; Ranieri, R.; Cuomo, G.; Abate, M.; Faggiana, G.; Proto, M.C.; Fiore, D.; et al. Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacol. Ther. 2017, 175, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Trofin, I.G.; Dabija, G.; Vaireanu, D.I.; Filipescu, L. Long term storage and cannabis oil stability. Rev. Chim. 2012, 63, 293–297. [Google Scholar]
- Herring, A.C.; Faubert Kaplan, B.L.; Kaminski, N.E. Modulation of CREB and NFkB signal transduction by cannabinol in activated thymocytes. Cell. Signal. 2001, 13, 241–250. [Google Scholar] [CrossRef]
- Janssens, A.; Silvestri, C.; Martella, A.; Vanoevelen, J.M.; Di Marzo, V.; Voets, T. Δ9-tetrahydrocannabivarin impairs epithelial calcium transport through inhibition of TRPV5 and TRPV6. Pharmacol. Res. 2018, 136, 83–89. [Google Scholar] [CrossRef]
- Citti, C.; Linciano, P.; Russo, F.; Luongo, L.; Iannotta, M.; Maione, S.; Lagana, A.; Capriotti, A.L.; Forni, F.; Vandelli, M.A.; et al. A novel phytocannabinoid isolated from Cannabis sativa L. with an in vivo cannabimimetic activity higher than Δ9-tetrahydrocannabinol: Δ9-tetrahydrocannabiphorol. Sci. Rep. 2019, 9, 20335. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ruiz, J.; Gómez-Ruiz, M.; García, C.; Hernández, M.; Ramos, J.A. Modeling Neurodegenerative Disorders for Developing Cannabinoid-Based Neuroprotective Therapies. Meth. Enzymol. 2017, 593, 175–198. [Google Scholar] [CrossRef]
- Ramírez, B.G.; Blázquez, C.; Gómez del Pulgar, T.; Guzmán, M.; de Ceballos, M.L. Prevention of Alzheimer’s disease pathology by cannabinoids: Neuroprotection mediated by blockade of microglial activation. J Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef] [Green Version]
- Varvel, S.; Hamm, R.; Martin, B.; Lichtman, A. Differential effects of Δ9-THC on spatial reference and working memory in mice. Psychopharmacology 2001, 157, 142–150. [Google Scholar] [CrossRef]
- Hamley, I.W. The amyloid beta peptide: A chemist’s perspective. Role in Alzheimer’s and fibrillization. Chem. Rev. 2012, 112, 5147–5192. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Li, Y.; Liu, H.; Bai, G.; Mayl, J.; Lin, X.; Sutherland, K.; Nabar, N.; Cai, J. The potential therapeutic effects of THC on Alzheimer’s disease. J. Alzheimers Dis. 2014, 42, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilkei-Gorzo, A.; Albayram, O.; Draffehn, A.; Michel, K.; Piyanova, A.; Oppenheimer, H.; Dvir-Ginzberg, M.; Rácz, I.; Ulas, T.; Imbeault, S.; et al. A chronic low dose of Δ9-tetrahydrocannabinol (THC) restores cognitive function in old mice. Nat. Med. 2017, 23, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Kalueff, A.V. The behavioral effects of acute Δ⁹-tetrahydrocannabinol and heroin (diacetylmorphine) exposure in adult zebrafish. Brain Res. 2014, 1543, 109–119. [Google Scholar] [CrossRef]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The effect of light spectrum on the morphology and cannabinoid content of Cannabis sativa L. Med. Cannabis Cannabinoids 2018, 1, 19–27. [Google Scholar] [CrossRef]
- Desaulniers Brousseau, V.; Wu, B.S.; MacPherson, S.; Morello, V.; Lefsrud, M. Cannabinoids and Terpenes: How Production of Photo-Protectants Can Be Manipulated to Enhance Cannabis sativa L. Phytochemistry. Front. Plant. Sci. 2021, 12, 620021. [Google Scholar] [CrossRef]
- Nazario, L.R.; Antonioli, R.; Capiotti, K.M.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Bonan, C.D.; da Silva, R.S. Caffeine protects against memory loss induced by high and non-anxiolytic dose of cannabidiol in adult zebrafish (Danio rerio). Pharmacol. Biochem. Behav. 2015, 135, 210–216. [Google Scholar] [CrossRef]
- Barichello, T.; Ceretta, R.A.; Generoso, J.S.; Moreira, A.P.; Simões, L.R.; Comim, C.M.; Quevedo, J.; Vilela, M.C.; Zuardi, A.W.; Crippa, J.A.; et al. Cannabidiol reduces host immune response and prevents cognitive impairments in Wistar rats submitted to pneumococcal meningitis. Eur. J. Pharmacol. 2012, 697, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Merrick, J.; Lane, B.; Sebree, T.; Yaksh, T.; O’Neill, C.; Banks, S.L. Identification of Psychoactive Degradants of Cannabidiol in Simulated Gastric and Physiological Fluid. Cannabis Cannabinoid Res. 2016, 1, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valim Brigante, T.A.; Abe, F.R.; Zuardi, A.W.; Hallak, J.E.C.; Crippa, J.A.S.; de Oliveira, D.P. Cannabidiol did not induce teratogenicity or neurotoxicity in exposed zebrafish embryos. Chem. Biol. Interact. 2018, 291, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Dysarz, F.A.; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar] [PubMed]
- Piomelli, D. The molecular logic of endocannabinoid signalling. Nat. Rev. Neurosci. 2003, 4, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Katona, I.; Freund, T.F. Endocannabinoid signaling as a synaptic circuit breaker in neurological disease. Nat. Med. 2008, 14, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Lee, D.; Cox, C.D.; Karsten, C.A.; Peñagarikano, O.; Geschwind, D.H.; Gall, C.M.; Piomelli, D. Endocannabinoid signaling mediates oxytocin-driven social reward. Proc. Natl. Acad. Sci. USA 2015, 112, 14084–14089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogi, A.; Mariti, C.; Pirrone, F.; Baragli, P.; Gazzano, A. The Influence of Oxytocin on Maternal Care in Lactating Dogs. Animals 2021, 11, 1130. [Google Scholar] [CrossRef]
- Wei, D.; Dinh, D.; Lee, D.; Li, D.; Anguren, A.; Moreno-Sanz, G.; Gall, C.M.; Piomelli, D. Enhancement of Anandamide-Mediated Endocannabinoid Signaling Corrects Autism-Related Social Impairment. Cannabis Cannabinoid Res. 2016, 1, 81–89. [Google Scholar] [CrossRef]
- Carnovali, M.; Ottria, R.; Pasqualetti, S.; Banfi, G.; Ciuffreda, P.; Mariotti, M. Effects of bioactive fatty acid amide derivatives in zebrafish scale model of bone metabolism and disease. Pharmacol. Res. 2016, 104, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Migliarini, B.; Carnevali, O. Anandamide modulates growth and lipid metabolism in the zebrafish Danio rerio. Mol. Cell. Endocrinol. 2008, 286, S12–S16. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.; Pryce, G.; Visintin, C.; Sisay, S.; Bondarenko, A.I.; Vanessa Ho, W.S.; Jackson, S.J.; Williams, T.E.; Al-Izki, S.; Sevastou, I.; et al. Big conductance calcium-activated potassium channel openers control spasticity without sedation. Br. J. Pharmacol. 2017, 174, 2662–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, I.; Kalló, I.; Deli, L.; Vida, B.; Hrabovszky, E.; Fekete, C.; Moenter, S.M.; Watanabe, M.; Liposits, Z. Retrograde endocannabinoid signaling reduces GABAergic synaptic transmission to gonadotropin-releasing hormone neurons. Endocrinology 2010, 151, 5818–5829. [Google Scholar] [CrossRef] [Green Version]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB₁ receptors regulate neuronal energy metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Compagnucci, C.; Di Siena, S.; Bustamante, M.B.; Di Giacomo, D.; Di Tommaso, M.; Maccarrone, M.; Grimaldi, P.; Sette, C. Type-1 (CB1) cannabinoid receptor promotes neuronal differentiation and maturation of neural stem cells. PLoS ONE 2013, 8, e54271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo-Canabal, A.; Colón-Cruz, L.; Rodriguez-Morales, R.; Varshney, G.K.; Burgess, S.; González-Sepúlveda, L.; Yudowski, G.; Behra, M. Altered Swimming Behaviors in Zebrafish Larvae Lacking Cannabinoid Receptor 2. Cannabis Cannabinoid Res. 2019, 4, 88–101. [Google Scholar] [CrossRef]
- Ortega-Alvaro, A.; Aracil-Fernández, A.; García-Gutiérrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 2011, 36, 1489–1504. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.R.; Canseco-Alba, A.; Zhang, H.Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Escosteguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J. CB2 cannabinoid receptor knockout in mice impairs contextual long-term memory and enhances spatial working memory. Neural. Plast. 2016, 2016, 9817089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Z.X.; Peng, X.Q.; Li, X.; Song, R.; Zhang, H.Y.; Liu, Q.R.; Yang, H.J.; Bi, G.H.; Li, J.; Gardner, E.L. Brain cannabinoid CB₂ receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gutierrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Gutiérrez, M.S.; Pérez-Ortiz, J.M.; Gutiérrez-Adán, A.; Manzanares, J. Depression-resistant endophenotype in mice overexpressing cannabinoid CB(2) receptors. Br. J. Pharmacol. 2010, 160, 1773–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stempel, A.V.; Stumpf, A.; Zhang, H.Y.; Özdoğan, T.; Pannasch, U.; Theis, A.K.; Otte, D.M.; Wojtalla, A.; Rácz, I.; Ponomarenko, A.; et al. Cannabinoid Type 2 Receptors Mediate a Cell Type-Specific Plasticity in the Hippocampus. Neuron 2016, 90, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Benarroch, E. Endocannabinoids in basal ganglia circuits: Implications for Parkinson disease. Neurology 2007, 69, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.R.; Ahmed, K.T.; Ali, D.W. Early Exposure to THC Alters M-Cell Development in Zebrafish Embryos. Biomedicines 2020, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basavarajappa, B.S.; Shivakumar, M.; Joshi, V.; Subbanna, S. Endocannabinoid system in neurodegenerative disorders. J. Neurochem. 2017, 142, 624–648. [Google Scholar] [CrossRef] [PubMed]
- Van Wijk, R.C.; Krekels, E.H.J.; Kantae, V.; Harms, A.C.; Hankemeier, T.; van der Graaf, P.H.; Spaink, H.P. Impact of post-hatching maturation on the pharmacokinetics of paracetamol in zebrafish larvae. Sci. Rep. 2019, 9, 2149. [Google Scholar] [CrossRef] [PubMed]
- Burton, C.E.; Zhou, Y.; Bai, Q.; Burton, E.A. Spectral properties of the zebrafish visual motor response. Neurosci. Let. 2017, 646, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Krug, R.G.; Clark, K.J. Elucidating cannabinoid biology in zebrafish (Danio rerio). Gene 2015, 570, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Oltrabella, F.; Melgoza, A.; Nguyen, B.; Guo, S. Role of the endocannabinoid system in vertebrates: Emphasis on the zebrafish model. Dev. Growth Differ. 2017, 59, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Vaz, R.; Hofmeister, W.; Lindstrand, A. Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques. Int. J. Mol. Sci. 2019, 20, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, O.; Sicca, F.; Paoli, E.; Trovato, F.; Santorelli, F.M.; Ratto, G.M.; Marchese, M. Evolution of Epileptiform Activity in Zebrafish by Statistical-Based Integration of Electrophysiology and 2-Photon Ca2+ Imaging. Cells 2020, 9, 769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogi, L.; Marchese, M.; Cellerino, A.; Licitra, R.; Naef, V.; Mero, S.; Bibbiani, C.; Fronte, B. β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy. Nutrients 2021, 13, 1619. [Google Scholar] [CrossRef] [PubMed]
- Naef, V.; Marchese, M.; Ogi, A.; Fichi, G.; Galatolo, D.; Licitra, R.; Doccini, S.; Verri, T.; Argenton, F.; Morani, F.; et al. Efficient Neuroprotective Rescue of Sacsin-Related Disease Phenotypes in Zebrafish. Int. J. Mol. Sci. 2021, 22, 8401. [Google Scholar] [CrossRef] [PubMed]
- Carty, D.R.; Thornton, C.; Gledhill, J.H.; Willett, K.L. Developmental Effects of Cannabidiol and Δ9-Tetrahydrocannabinol in Zebrafish. Toxicol. Sci. 2018, 162, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchtenburg, F.J.; Schaaf, M.J.M.; Richardson, M.K. Functional characterization of the cannabinoid receptors 1 and 2 in zebrafish larvae using behavioral analysis. Psychopharmacology 2019, 236, 2049–2058. [Google Scholar] [CrossRef] [Green Version]
- Migliarini, B.; Carnevali, O. A novel role for the endocannabinoid system during zebrafish development. Mol. Cell. Endocrinol. 2009, 299, 172–177. [Google Scholar] [CrossRef]
- Lam, C.S.; Rastegar, S.; Strahle, U. Distribution of cannabinoid receptor 1 in the CNS of zebrafish. Neuroscience 2006, 138, 83–95. [Google Scholar] [CrossRef]
- Watson, S.; Chambers, D.; Hobbs, C.; Doherty, P.; Graham, A. The endocannabinoid receptor, CB1, is required for normal axonal growth and fasciculation. Mol. Cell. Neurosci. 2008, 38, 89–97. [Google Scholar] [CrossRef]
- Licitra, R.; Martinelli, M.; Petrocchi, J.L.; Marchese, M.; Kiferle, C.; Fronte, B. In Vivo Evaluation of Cannabis sativa Full Extract on Zebrafish Larvae Development, Locomotion Behavior and Gene Expression. Pharmaceuticals 2021, 14, 1224. [Google Scholar] [CrossRef] [PubMed]
- Nixon, J.; Abramovici, H.; Cabecinha, A.; Martinez-Farina, C.; Hui, J.; Ellis, L. Assessing the bioactivity of cannabis extracts in larval zebrafish. J. Cannabis Res. 2021, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Using zebrafish to unravel the genetics of complex brain disorders. Curr. Top. Behav. Neurosci. 2012, 12, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Kirla, K.T.; Groh, K.J.; Steuer, A.E.; Poetzsch, M.; Banote, R.K.; Stadnicka-Michalak, J.; Eggen, R.I.; Schirmer, K.; Kraemer, T. From the Cover: Zebrafish Larvae Are Insensitive to Stimulation by Cocaine: Importance of Exposure Route and Toxicokinetics. Toxicol. Sci. 2016, 154, 183–193. [Google Scholar] [CrossRef]
- MacPhail, R.C.; Brooks, J.; Hunter, D.L.; Padnos, B.; Irons, T.D.; Padilla, S. Locomotion in larval zebrafish: Influence of time of day, lighting and ethanol. Neurotoxicology 2009, 30, 52–58. [Google Scholar] [CrossRef]
- Maximino, C.; De Brito, T.M.; de Mattos Dias, C.A.G.; Gouveia, A., Jr.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Kaluyeva, A.A.; Poudel, M.K.; Nguyen, M.; Song, C.; Kalueff, A.V. Building Zebrafish Neurobehavioral Phenomics: Effects of Common Environmental Factors on Anxiety and Locomotor Activity. Zebrafish 2015, 12, 339–348. [Google Scholar] [CrossRef]
- de Abreu, M.S.; Friend, A.J.; Demin, K.A.; Amstislavskaya, T.G.; Bao, W.; Kalueff, A.V. Zebrafish models: Do we have valid paradigms for depression? J. Pharmacol. Toxicol. Methods 2018, 94, 16–22. [Google Scholar] [CrossRef]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Versatile Setup for Measuring Multiple Behavior Endpoints in Zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Ogi, A.; Licitra, R.; Naef, V.; Marchese, M.; Fronte, B.; Gazzano, A.; Santorelli, F.M. Social Preference Tests in Zebrafish: A Systematic Review. Front. Vet. Sci. 2021, 7, 590057. [Google Scholar] [CrossRef] [PubMed]
- De Abreu, M.S.; Giacomini, A.; Genario, R.; Dos Santos, B.E.; da Rosa, L.G.; Demin, K.A.; Wappler-Guzzetta, E.A.; Kalueff, A.V. Neuropharmacology, pharmacogenetics and pharmacogenomics of aggression: The zebrafish model. Pharmacol. Res. 2019, 141, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.J. The toxicologic and teratologic effects of delta-9-tetrahydrocannabinol in the zebrafish embryo. Toxicol. Appl. Pharmacol. 1975, 32, 184–190. [Google Scholar] [CrossRef]
- Huestis, M.A. Human cannabinoid pharmacokinetics. Chem. Biodivers. 2007, 4, 1770–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifici, R.; Pichini, S.; Pellegrini, M.; Rotolo, M.C.; Giorgetti, R.; Tagliabracci, A.; Busardò, F.P.; Huestis, M.A. THC and CBD concentrations in blood, oral fluid and urine following a single and repeated administration of “light cannabis”. Clin. Chem. Lab. Med. 2020, 58, 682–689. [Google Scholar] [CrossRef]
- Clift, D.; Richendrfer, H.; Thorn, R.J.; Colwill, R.M.; Creton, R. High-throughput analysis of behavior in zebrafish larvae: Effects of feeding. Zebrafish 2014, 11, 455–461. [Google Scholar] [CrossRef]
- Grisham, M.G.; Ferraro, D.P. Biphasic effects of 9-tetrahydrocannabinol on variable interval schedule performance in rats. Psychopharmacology 1972, 27, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Breivogel, C.S.; Childers, S.R.; Deadwyler, S.A.; Hampson, R.E.; Porrino, L.J. Cannabinoid physiology and pharmacology: 30 years of progress. Neuropharmacology 2004, 47, 345–358. [Google Scholar] [CrossRef]
- Morris, C.V.; DiNieri, J.A.; Szutorisz, H.; Hurd, Y.L. Molecular mechanisms of maternal cannabis and cigarette use on human neurodevelopment. Eur. J. Neurosci. 2011, 34, 1574–1583. [Google Scholar] [CrossRef] [Green Version]
- Hasumi, A.; Maeda, H.; Yoshida, K. Analyzing cannabinoid-induced abnormal behavior in a zebrafish model. PLoS ONE 2020, 15, e0236606. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, H.; Li, C.-Z.; Lai, P.-S.; Wang, G.; Chan, Y.S.; Cheng, S.H.; Chen, X. Cannabidiol promotes fin regeneration and reduces apoptosis in zebrafish embryos. J. Funct. Foods 2021, 86, 104694. [Google Scholar] [CrossRef]
- Ruhl, T.; Prinz, N.; Oellers, N.; Seidel, N.I.; Jonas, A.; Albayram, O.; Bilkei-Gorzo, A.; Von Der Emde, G. Acute administration of THC impairs spatial but not associative memory function in zebrafish. Psychopharmacology 2014, 231, 3829–3842. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.A.; Aguiar, D.C.; Guimaraes, F.S. Anxiolytic-like effect of cannabidiol in the rat Vogel conflict test. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 1466–1471. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.J.; Muller, C.H.; Murataeva, N.A.; Davis, B.J.; Mackie, K. Δ 9-tetrahydrocannabinol (Δ 9-THC) attenuates mouse sperm motility and male fecundity. Br. J. Pharmacol. 2012, 165, 2575–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.E.M.; Rapino, C.; Di Tommaso, M.; Pucci, M.; Battista, N.; Paro, R.; Simon, L.; Lutton, D.; Maccarrone, M. Differences in the endocannabinoid system of sperm from fertile and infertile men. PLoS ONE 2012, 7, e47704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bara, A.; Manduca, A.; Bernabeu, A.; Borsoi, M.; Serviado, M.; Lassalle, O.; Murphy, M.; Wager-Miller, J.; Mackie, K.; Pelissier-Alicot, A.-L.; et al. Sex-dependent effects of in utero cannabinoid exposure on cortical function. eLife 2018, 7, e36234. [Google Scholar] [CrossRef]
- Navarro, M.; Rubio, P.; de Fonseca, F.R. Behavioural consequences of maternal exposure to natural cannabinoids in rats. Psychopharmacology 1995, 122, 226. [Google Scholar] [CrossRef] [PubMed]
- Samarut, É.; Nixon, J.; Kundap, U.P.; Drapeau, P.; Ellis, L.D. Single and Synergistic Effects of Cannabidiol and Δ-9-Tetrahydrocannabinol on Zebrafish Models of Neuro-Hyperactivity. Front. Pharmacol. 2019, 10, 226. [Google Scholar] [CrossRef]
- Tang, F.; Hartz, A.M.S.; Bauer, B. Drug-Resistant Epilepsy: Multiple Hypotheses, Few Answers. Front. Neurol. 2017, 8, 301. [Google Scholar] [CrossRef]
- Zaheer, S.; Kumar, D.; Khan, M.T.; Giyanwani, P.R.; Kiran, F. Epilepsy and Cannabis: A Literature Review. Cureus 2018, 10, e3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Caro, C.; Leo, A.; Citraro, R.; De Sarro, C.; Russo, R.; Calignano, A.; Russo, E. The potential role of cannabinoids in epilepsy treatment. Expert Rev. Neurother. 2017, 17, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, S.; Mammana, S.; Cavalli, E.; Bramanti, P.; Mazzon, E. Use of Cannabidiol in the Treatment of Epilepsy: Efficacy and Security in Clinical Trials. Molecules 2019, 24, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Connors, K.A.; Valenti, T.W.; Lawless, K.; Sackerman, J.; Onaivi, E.S.; Brooks, B.W.; Gould, G.G. Similar anxiolytic effects of agonists targeting serotonin 5-HT1A or cannabinoid CB receptors on zebrafish behavior in novel environments. Aquat. Toxicol. 2014, 151, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Emran, F.; Rihel, J.; Dowling, J.E. A Behavioral Assay to Measure Responsiveness of Zebrafish to Changes in Light Intensities. J. Vis. Exp. 2008, 20, 923. [Google Scholar] [CrossRef]
- Ganzen, L.; Venkatraman, P.; Pang, C.P.; Leung, Y.F.; Zhang, M. Utilizing Zebrafish Visual Behaviors in Drug Screening for Retinal Degeneration. Int. J. Mol. Sci. 2017, 18, 1185. [Google Scholar] [CrossRef] [Green Version]
- Knafo, S.; Wyart, C. Active mechanosensory feedback during locomotion in the zebrafish spinal cord. Curr. Opin. Neurobiol. 2018, 52, 48–53. [Google Scholar] [CrossRef]
- El-Daher, F.; Becker, C.G. Neural circuit reorganisation after spinal cord injury in zebrafish. Curr. Opin. Genet. Dev. 2020, 64, 44–51. [Google Scholar] [CrossRef]
- Danos, N.; Lauder, G.V. Challenging zebrafish escape responses by increasing water viscosity. J. Exp. Biol. 2012, 215, 1854–1862. [Google Scholar] [CrossRef] [Green Version]
- Sillar, K.T. Mauthner cells. Curr. Biol. 2009, 19, 353–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Carmer, R.; Zhang, G.; Venkatraman, P.; Brown, S.A.; Pang, C.-P.; Zhang, M.; Ma, P.; Leung, Y.F. Statistical Analysis of Zebrafish Locomotor Response. PLoS ONE 2015, 10, e0139521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Licitra, R.; Marchese, M.; Naef, V.; Ogi, A.; Martinelli, M.; Kiferle, C.; Fronte, B.; Santorelli, F.M. A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment. Biomedicines 2022, 10, 1820. https://doi.org/10.3390/biomedicines10081820
Licitra R, Marchese M, Naef V, Ogi A, Martinelli M, Kiferle C, Fronte B, Santorelli FM. A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment. Biomedicines. 2022; 10(8):1820. https://doi.org/10.3390/biomedicines10081820
Chicago/Turabian StyleLicitra, Rosario, Maria Marchese, Valentina Naef, Asahi Ogi, Marco Martinelli, Claudia Kiferle, Baldassare Fronte, and Filippo Maria Santorelli. 2022. "A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment" Biomedicines 10, no. 8: 1820. https://doi.org/10.3390/biomedicines10081820
APA StyleLicitra, R., Marchese, M., Naef, V., Ogi, A., Martinelli, M., Kiferle, C., Fronte, B., & Santorelli, F. M. (2022). A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment. Biomedicines, 10(8), 1820. https://doi.org/10.3390/biomedicines10081820