
Association between Polymorphisms in CFH, ARMS2, CFI, and C3 Genes and Response to Anti-VEGF Treatment in Neovascular Age-Related Macular Degeneration

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. OCT Subgroup Prospective Study
2.3. Sample Collection
2.4. DNA Isolation and Genotyping
2.5. Measurements of Complement Factor I Level
2.6. In Silico Analysis of SNPs Regulatory Potential
2.7. Statistical Analysis
3. Results
3.1. Association with nAMD Development
3.2. Association between SNPs and OCT-Markers
3.3. Functional Annotation of the Studied Gene Polymorphisms
3.4. Effect of rs2285714 on Complement Factor I Plasma and Aqueous Humor Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spaide, R.F.; Jaffe, G.J.; Sarraf, D.; Freund, K.B.; Sadda, S.R.; Staurenghi, G.; Waheed, N.K.; Chakravarthy, U.; Rosenfeld, P.J.; Holz, F.G.; et al. Consensus Nomenclature for Reporting Neovascular Age-Related Macular Degeneration Data: Consensus on Neovascular Age-Related Macular Degeneration Nomenclature Study Group. Ophthalmology 2020, 127, 616–636. [Google Scholar] [CrossRef] [PubMed]
- Armento, A.; Ueffing, M.; Clark, S.J. The complement system in age-related macular degeneration. Cell. Mol. Life Sci. 2021, 78, 4487–4505. [Google Scholar] [CrossRef] [PubMed]
- Lorés-Motta, L.; de Jong, E.K.; den Hollander, A.I. Exploring the Use of Molecular Biomarkers for Precision Medicine in Age-Related Macular Degeneration. Mol. Diagn. Ther. 2018, 22, 315–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fursova, A.Z.; Chubar, N.V.; Tarasov, M.S.; Vasilyeva, M.A.; Gusarevich, O.G. Anti-VEGF therapy for age-related macular degeneration. Vestn. Oftalmol. 2018, 134, 59–67. [Google Scholar] [CrossRef]
- Faatz, H.; Rothaus, K.; Ziegler, M.; Book, M.; Heimes-Bussmann, B.; Pauleikhoff, D.; Lommatzsch, A. Vascular Analysis of Type 1, 2, and 3 Macular Neovascularization in Age-Related Macular Degeneration Using Swept-Source Optical Coherence Tomography Angiography Shows New Insights into Differences of Pathologic Vasculature and May Lead to a More Personalized Understanding. Biomedicines 2022, 10, 694. [Google Scholar] [CrossRef]
- Metrangolo, C.; Donati, S.; Mazzola, M.; Fontanel, L.; Messina, W.; D’alterio, G.; Rubino, M.; Radice, P.; Premi, E.; Azzolini, C. OCT Biomarkers in Neovascular Age-Related Macular Degeneration: A Narrative Review. J. Ophthalmol. 2021, 2021, 9994098. [Google Scholar] [CrossRef]
- Fursova, A.Z.; Derbeneva, A.S.; Tarasov, M.S.; Nikulich, I.F.; Devyatkin, V.A.; Telegina, D.V.; Kolosova, N.G.; Kozhevnikova, O.S. Leukocyte telomere length and response to antiangiogenic therapy in patients with neovascular age-related macular degeneration. Adv. Gerontol. Uspekhi Gerontol. 2021, 34, 823–830. [Google Scholar] [CrossRef]
- Clark, S.J.; Bishop, P.N. The eye as a complement dysregulation hotspot. Semin. Immunopathol. 2018, 40, 65–74. [Google Scholar] [CrossRef]
- Seddon, J.M.; Yu, Y.; Miller, E.C.; Reynolds, R.; Tan, P.L.; Gowrisankar, S.; Goldstein, J.I.; Triebwasser, M.; Anderson, H.E.; Zerbib, J.; et al. Rare variants in CFI, C3 and C9 are associated with high risk of advanced age-related macular degeneration. Nat. Genet. 2013, 45, 1366–1370. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, D.; Yu, Y.; Schramm, E.C.; Triebwasser, M.; Wagner, E.K.; Raychaudhuri, S.; Daly, M.J.; Atkinson, J.P.; Seddon, J.M. Rare genetic variants in the CFI gene are associated with advanced age-related macular degeneration and commonly result in reduced serum factor I levels. Hum. Mol. Genet. 2015, 24, 3861–3870. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, L.G.; Igl, W.; Bailey, J.N.; Grassmann, F.; Sengupta, S.; Bragg-Gresham, J.L.; Burdon, K.P.; Hebbring, S.J.; Wen, C.; Gorski, M.; et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2016, 48, 134–143. [Google Scholar] [CrossRef] [Green Version]
- Micklisch, S.; Lin, Y.; Jacob, S.; Karlstetter, M.; Dannhausen, K.; Dasari, P.; von der Heide, M.; Dahse, H.M.; Schmölz, L.; Grassmann, F.; et al. Age-related macular degeneration associated polymorphism rs10490924 in ARMS2 results in deficiency of a complement activator. J. Neuroinflamm. 2017, 14, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.M.; Michels, M.; Kaiser, P.K.; Heier, J.S.; Sy, J.P.; Ianchulev, T.; ANCHOR Study Group. Ranibizumab versus verteporfin photodynamic therapy for neovascular age-related macular degeneration: Two-year results of the ANCHOR study. Ophthalmology 2009, 116, 57–65.e5. [Google Scholar] [CrossRef]
- Rosenfeld, P.J.; Brown, D.M.; Heier, J.S.; Boyer, D.S.; Kaiser, P.K.; Chung, C.Y.; Kim, R.Y.; MARINA Study Group. Ranibizumab for neovascular age-related macular degeneration. N. Eng. J. Med. 2006, 355, 1419–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldstein, S.M.; Simader, C.; Staurenghi, G.; Chong, N.V.; Mitchell, P.; Jaffe, G.J.; Lu, C.; Katz, T.A.; Schmidt-Erfurth, U. Morphology and Visual Acuity in Aflibercept and Ranibizumab Therapy for Neovascular Age-Related Macular Degeneration in the VIEW Trials. Ophthalmology 2016, 123, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Framme, C.; Eter, N.; Hamacher, T.; Hasanbasic, Z.; Jochmann, C.; Johnson, K.T.; Kahl, M.; Sachs, H.; Schilling, H.; Thelen, U.; et al. Aflibercept for Patients with Neovascular Age-Related Macular Degeneration in Routine Clinical Practice in Germany: Twelve-Month Outcomes of PERSEUS. Ophthalmol. Retin. 2018, 2, 539–549. [Google Scholar] [CrossRef]
- Bhisitkul, R.B.; Mendes, T.S.; Rofagha, S.; Enanoria, W.; Boyer, D.S.; Sadda, S.R.; Zhang, K. Macular atrophy progression and 7-year vision outcomes in subjects from the ANCHOR, MARINA, and HORIZON studies: The SEVEN-UP study. Am. J. Ophthalmol. 2015, 159, 915–924.e2. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, J.; Sun, X. Resistance to anti-VEGF therapy in neovascular age-related macular degeneration: A comprehensive review. Drug Des. Dev. Ther. 2016, 10, 1857–1867. [Google Scholar] [CrossRef] [Green Version]
- Dedania, V.S.; Grob, S.; Zhang, K.; Bakri, S.J. Pharmacogenomics of response to anti-VEGF therapy in exudative age-related macular degeneration. Retina 2015, 35, 381–391. [Google Scholar] [CrossRef]
- Enders, P.; Scholz, P.; Muether, P.S.; Fauser, S. Variability of disease activity in patients treated with ranibizumab for neovascular age-related macular degeneration. Eye 2016, 30, 1072–1076. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Zhou, Y.; Yi, X.; Fan, X.; Wang, J.; Yao, H.; Sham, P.C.; Hao, J.; Chen, K.; Li, M.J. VannoPortal: Multiscale functional annotation of human genetic variants for interrogating molecular mechanism of traits and diseases. Nucleic Acids Res. 2022, 50, D1408–D1416. [Google Scholar] [CrossRef] [PubMed]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oca, A.I.; Pérez-Sala, Á.; Pariente, A.; Ochoa, R.; Velilla, S.; Peláez, R.; Larráyoz, I.M. Predictive Biomarkers of Age-Related Macular Degeneration Response to Anti-VEGF Treatment. J. Pers. Med. 2021, 11, 1329. [Google Scholar] [CrossRef]
- Maroñas, O.; García-Quintanilla, L.; Luaces-Rodríguez, A.; Fernández-Ferreiro, A.; Latorre-Pellicer, A.; Abraldes, M.J.; Lamas, M.J.; Carracedo, A. Anti-VEGF Treatment and Response in Age-related Macular Degeneration: Disease’s Susceptibility, Pharmacogenetics and Pharmacokinetics. Curr. Med. Chem. 2020, 27, 549–569. [Google Scholar] [CrossRef]
- Hagstrom, S.A.; Ying, G.S.; Pauer, G.; Sturgill-Short, G.M.; Huang, J.; Callanan, D.G.; Kim, I.K.; Klein, M.L.; Maguire, M.G.; Martin, D.F.; et al. Pharmacogenetics for genes associated with age-related macular degeneration in the Comparison of AMD Treatments Trials (CATT). Ophthalmology 2013, 120, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagstrom, S.A.; Ying, G.S.; Maguire, M.G.; Martin, D.F.; CATT Research Group; Gibson, J.; Lotery, A.; Chakravarthy, U.; IVAN Study Investigators. VEGFR2 Gene Polymorphisms and Response to Anti-Vascular Endothelial Growth Factor Therapy in Age-Related Macular Degeneration. Ophthalmology 2015, 122, 1563–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloeckener-Gruissem, B.; Barthelmes, D.; Labs, S.; Schindler, C.; Kurz-Levin, M.; Michels, S.; Fleischhauer, J.; Berger, W.; Sutter, F.; Menghini, M. Genetic association with response to intravitreal ranibizumab in patients with neovascular AMD. Invest. Ophthalmol. Vis. Sci. 2011, 52, 4694–4702. [Google Scholar] [CrossRef]
- Nischler, C.; Oberkofler, H.; Ortner, C.; Paikl, D.; Riha, W.; Lang, N.; Patsch, W.; Egger, S.F. Complement factor H Y402H gene polymorphism and response to intravitreal bevacizumab in exudative age-related macular degeneration. Acta Ophthalmol. 2011, 89, e344–e349. [Google Scholar] [CrossRef]
- Lee, A.Y.; Raya, A.K.; Kymes, S.M.; Shiels, A.; Brantley, M.A., Jr. Pharmacogenetics of complement factor H (Y402H) and treatment of exudative age-related macular degeneration with ranibizumab. Br. J. Ophthalmol. 2009, 93, 610–613. [Google Scholar] [CrossRef] [Green Version]
- Hong, N.; Shen, Y.; Yu, C.Y.; Wang, S.Q.; Tong, J.P. Association of the polymorphism Y402H in the CFH gene with response to anti-VEGF treatment in age-related macular degeneration: A systematic review and meta-analysis. Acta Ophthalmol. 2016, 94, 334–345. [Google Scholar] [CrossRef] [Green Version]
- Abedi, F.; Wickremasinghe, S.; Richardson, A.J.; Islam, A.F.; Guymer, R.H.; Baird, P.N. Genetic influences on the outcome of anti-vascular endothelial growth factor treatment in neovascular age-related macular degeneration. Ophthalmology 2013, 120, 1641–1648. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Xie, P.; Ding, Y.; Yuan, D.; Liu, Q. Association between variants A69S in ARMS2 gene and response to treatment of exudative AMD: A meta-analysis. Br. J. Ophthalmol. 2015, 99, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xiong, H.; Xu, Y.; Xiong, X.; Zou, H.; Zheng, M.; Wang, X.; Zhou, X. Association between VEGF-A and VEGFR-2 polymorphisms and response to treatment of neovascular AMD with anti-VEGF agents: A meta-analysis. Br. J. Ophthalmol. 2017, 101, 976–984. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, M.; Takahashi, A.; Momozawa, Y.; Arakawa, S.; Miya, F.; Tsunoda, T.; Ashikawa, K.; Oshima, Y.; Yasuda, M.; Yoshida, S.; et al. Genome-wide association study suggests four variants influencing outcomes with ranibizumab therapy in exudative age-related macular degeneration. J. Hum. Genet. 2018, 63, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.M.; Rosen, R.B.; Meyerle, C.B.; Kurup, S.K.; Ardeljan, D.; Agron, E.; Tai, K.; Pomykala, M.; Chew, E.Y.; Chan, C.C.; et al. Suggestive association between PLA2G12A single nucleotide polymorphism rs2285714 and response to anti-vascular endothelial growth factor therapy in patients with exudative age-related macular degeneration. Mol. Vis. 2012, 18, 2578–2585. [Google Scholar] [PubMed]
- Aygun, F.B.; Kadayıfcılar, S.; Ozgul, R.K.; Eldem, B. Complement Factor I Gene Polymorphism in a Turkish Age-Related Macular Degeneration Population. Ophthalmologica 2020, 243, 187–194. [Google Scholar] [CrossRef]
- Balikova, I.; Postelmans, L.; Pasteels, B.; Coquelet, P.; Catherine, J.; Efendic, A.; Hosoda, Y.; Miyake, M.; Yamashiro, K.; Thienpont, B.; et al. Genetic biomarkers in the VEGF pathway predicting response to anti-VEGF therapy in age-related macular degeneration. BMJ Open Ophthalmol. 2019, 4, e000273. [Google Scholar] [CrossRef] [Green Version]
- Park, U.C.; Shin, J.Y.; McCarthy, L.C.; Kim, S.J.; Park, J.H.; Chung, H.; Yu, H.G. Pharmacogenetic associations with long-term response to anti-vascular endothelial growth factor treatment in neovascular AMD patients. Mol. Vis. 2014, 20, 1680–1694. [Google Scholar]
- Garrido-Martín, D.; Borsari, B.; Calvo, M.; Reverter, F.; Guigó, R. Identification and analysis of splicing quantitative trait loci across multiple tissues in the human genome. Nat. Commun. 2021, 12, 727. [Google Scholar] [CrossRef]
- Schick, T.; Steinhauer, M.; Aslanidis, A.; Altay, L.; Karlstetter, M.; Langmann, T.; Kirschfink, M.; Fauser, S. Local complement activation in aqueous humor in patients with age-related macular degeneration. Eye 2017, 31, 810–813. [Google Scholar] [CrossRef] [Green Version]
- Hallam, T.M.; Marchbank, K.J.; Harris, C.L.; Osmond, C.; Shuttleworth, V.G.; Griffiths, H.; Cree, A.J.; Kavanagh, D.; Lotery, A.J. Rare Genetic Variants in Complement Factor I Lead to Low FI Plasma Levels Resulting in Increased Risk of Age-Related Macular Degeneration. Invest. Ophthalmol. Vis. Sci. 2020, 61, 18. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, T.J.; Lechanteur, Y.; Lorés-Motta, L.; Schick, T.; Daha, M.R.; Altay, L.; Liakopoulos, S.; Smailhodzic, D.; den Hollander, A.I.; Hoyng, C.B.; et al. Complement Activation Levels Are Related to Disease Stage in AMD. Invest. Ophthalmol. Vis. Sci. 2020, 61, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, T.D.; Toso, M.; Pappas, C.; Nichols, L.; Bishop, P.N.; Hageman, G.S. Assessment of Proteins Associated With Complement Activation and Inflammation in Maculae of Human Donors Homozygous Risk at Chromosome 1 CFH-to-F13B. Invest. Ophthalmol. Vis. Sci. 2015, 56, 4870–4879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Oguchi, Y.; Omori, T.; Ishida, Y.; Shintake, H.; Tomita, R.; Kasai, A.; Ogasawara, M.; Sugano, Y.; Itagaki, K.; et al. Changes in complement activation products after anti-VEGF injection for choroidal neovascularization in age-related macular degeneration and pachychoroid disease. Sci. Rep. 2021, 11, 8464. [Google Scholar] [CrossRef] [PubMed]
- Medina, F.; Motta, A.; Takahashi, W.Y.; Carricondo, P.C.; Motta, M.; Melo, M.B.; Vasconcellos, J. Association of the CFH Y402H Polymorphism with the 1-Year Response of Exudative AMD to Intravitreal Anti-VEGF Treatment in the Brazilian Population. Ophthalmic Res. 2019, 61, 168–173. [Google Scholar] [CrossRef]
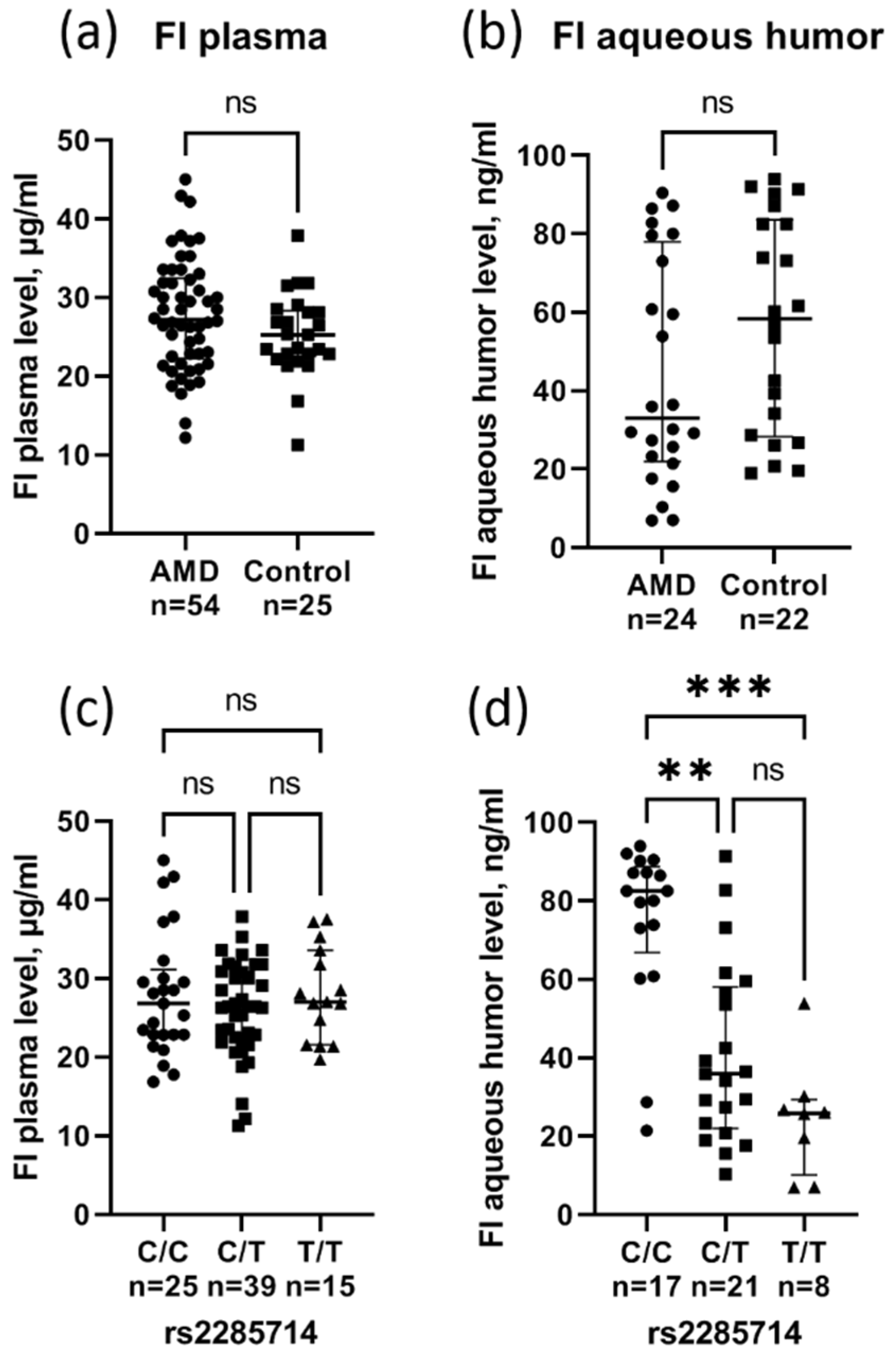

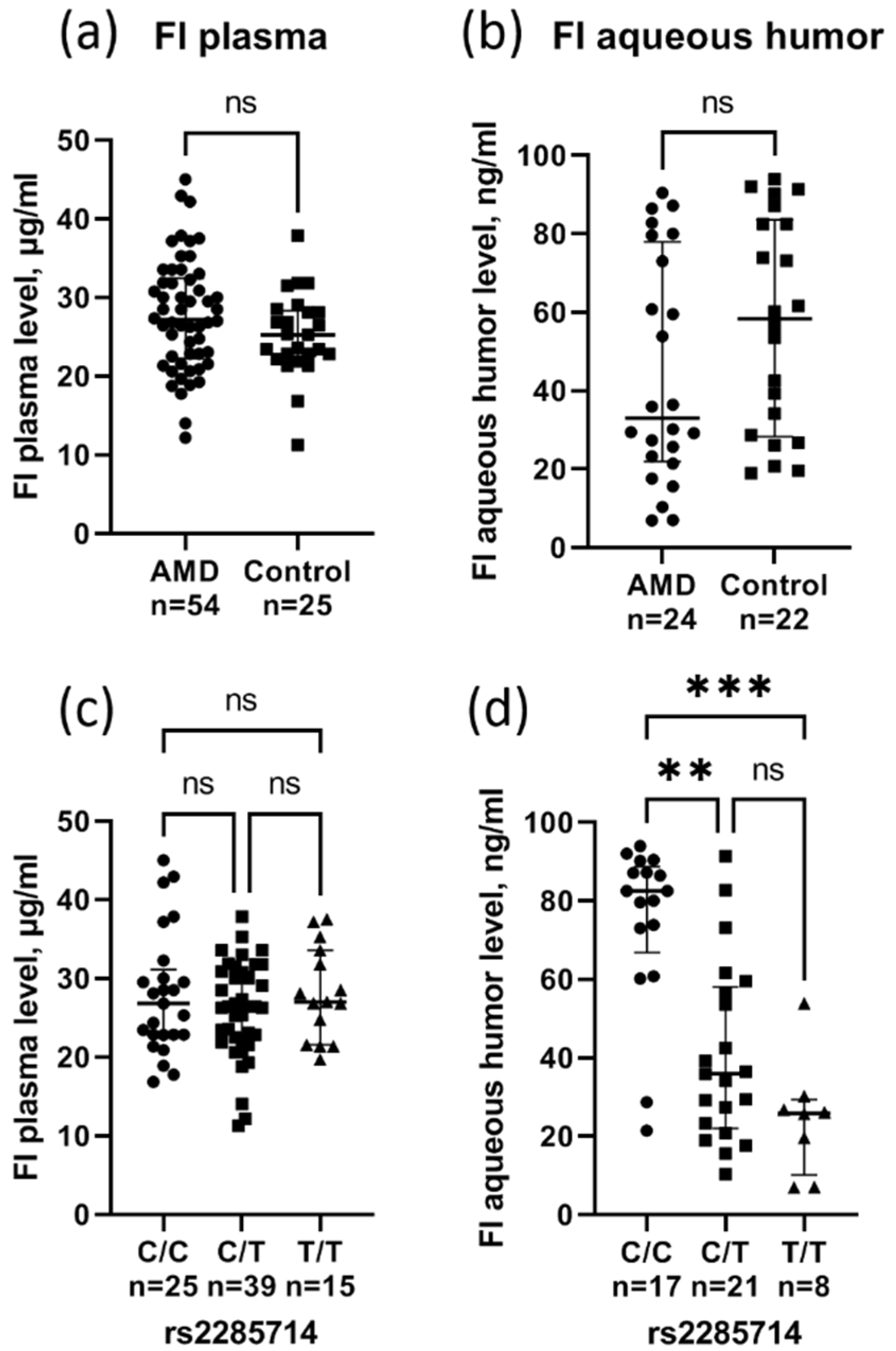{kind=link}
| SNP | Location/Consequence | Chromosome Position/Orientation | Nucleotide Substitution | MAF |
|---|---|---|---|---|
| rs800292 | Missense variant of the CFH: Val62Ile | 1:196673103, plus | G>A | 0.26 |
| rs6677604 | Intron variant of the CFH | 1:196717788, plus | G>A | 0.19 |
| rs2285714 | 3′ untranslated region of CFI; synonymous variant of PLA2G12A | 4:109717654, minus | C>T | 0.39 |
| rs10490924 | Missense variant of ARMS2: Ala69Ser; 5′ untranslated region of HTRA1 | 10:122454932, plus | G>T | 0.19 |
| rs2230199 | Missense variant of C3: Arg102Gly | 19:6718376, minus | G>C | 0.22 |
| rsID | Gene Name | Sequence |
|---|---|---|
| rs800292 | CFH | Forward 5′-AAGGCACCCAGGCTATCTAT-3′ |
| Reverse 5′-TAATGGATTAAGAGCAACCCATTCT-3′ | ||
| 5′-FAM-ATACCATTATT[+A][+T][+A]TTTCCAA-BHQ1-3′ | ||
| 5′-VIC-CATACCATTATT[+A][+C][+A]TTTCCAA-BHQ1-3′ | ||
| rs6677604 | CFH | Forward 5′-ACACCAGAGCAGATACAGCA-3′ |
| Reverse 5′-TGCCACCAAAGCACAATACC-3′ | ||
| 5′-FAM-CCTTTC[+C]T[+C]TCGCATTTTCTC-BHQ1-3′ | ||
| 5′-VIC-CCTTTC[+C]C[+C]TCGCATTTTCTC-BHQ1-3′ | ||
| rs2285714 | CFI/PLA2G12A | Forward 5′-GTGTTTTCTGTACATCTCGGCA-3′ |
| Reverse 5′-TGCCTTTTGCAGCTTAACATTG-3′ | ||
| 5′-FAM-TGCCACAGGTCTCATAGCACCTGT-BHQ1-3′ | ||
| 5′-VIC-TGCCACAGGTTTCATAGCACCTGTC-BHQ2-3′ | ||
| rs10490924 | ARMS2/HTRA1 | Forward 5′-AGTGACAAGCAGAGGAGCAA-3′ |
| Reverse 5′-CAGCAGGAGAGAAGAAGGCT-3′ | ||
| 5′-FAM-CATGATCCCAGCTGCTAAAATCCA-BHQ1-3′ | ||
| 5′-VIC-CCATGATCCCAGCTTCTAAAATCCAC-BHQ1-3′ | ||
| rs2230199 | C3 | Forward 5′-TGGTCTTGTCTGTCTGGATGAA-3′ |
| Reverse 5′-CAAGATCCGGAAGCTGGAC-3′ | ||
| 5′-FAM-CGAACTTGTTGCCCCCCTTTTC-BHQ1-3′ | ||
| 5′-VIC-CGAACTTGTTGCGCCCCTTTT-BHQ1-3′ |
| SNP | Genotype/Allele | Case | Control | HWE a (Case) | HWE a (Control) |
|---|---|---|---|---|---|
| rs800292 G>A | G/G | 138 (72%) | 77 (52%) | 0.79 | 0.84 |
| A/G | 50 (26%) | 60 (41%) | |||
| A/A | 5 (3%) | 10 (7%) | |||
| A | 60 (16%) | 80 (27%) | |||
| rs6677604 G>A | G/G | 152 (79%) | 82 (56%) | 0.3 | 0.67 |
| A/G | 37 (19%) | 54 (37%) | |||
| A/A | 4 (2%) | 11 (7%) | |||
| A | 45 (12%) | 76 (26%) | |||
| rs2285714 C>T | C/C | 61 (32%) | 56 (38%) | 1 | 0.39 |
| C/T | 95 (49%) | 65 (44%) | |||
| T/T | 37 (19%) | 26 (18%) | |||
| T | 169 (44%) | 117 (40%) | |||
| rs10490924 G>T | G/G | 40 (21%) | 70 (48%) | 0.31 | 0.26 |
| G/T | 88 (46%) | 58 (39%) | |||
| T/T | 65 (34%) | 19 (13%) | |||
| T | 218 (56%) | 168 (33%) | |||
| rs2230199 G>C | G/G | 121 (63%) | 100 (68%) | 0.51 | 0.57 |
| G/C | 66 (34%) | 41 (28%) | |||
| C/C | 6 (3%) | 6 (4%) | |||
| C | 78 (20%) | 53 (18%) |
| SNP | Model of Inheritance | OR (95% CI) Adjusted for Sex and Age | p-Value | AIC |
|---|---|---|---|---|
| rs800292 G>A | Dominant: A/A+A/G vs. G/G | 0.44 (0.28–0.69) | 0.0004 | 453.7 |
| Recessive: A/A vs. G/G+A/G | 0.36 (0.12–1.09 | 0.062 | 463 | |
| Additive | 0.49 (0.33–0.72) | 0.0002 | 453 | |
| rs6677604 G>A | Dominant: A/A+A/G vs. G/G | 0.36 (0.22–0.58) | <0.0001 | 448.5 |
| Recessive: A/A vs. G/G+A/G | 0.27 (0.08–0.89) | 0.022 | 461.2 | |
| Additive | 0.41 (0.27–0.63) | <0.0001 | 447.6 | |
| rs2285714 C>T | Dominant: C/T+T/T vs. C/C | 1.36 (0.86–2.14) | 0.19 | 464.8 |
| Recessive: T/T vs. C/C+C/T | 1.12 (0.64–1.95) | 0.7 | 466.4 | |
| Additive | 1.19 (0.87–1.61) | 0.28 | 465.3 | |
| rs10490924 G>T | Dominant: G/T+T/T vs. G/G | 3.35 (2.06–5.45) | <0.0001 | 441.7 |
| Recessive: T/T vs. G/T+G/G | 3.42 (1.93–6.05) | <0.0001 | 446.6 | |
| Additive | 2.43 (1.76–3.36) | <0.0001 | 434.1 | |
| rs2230199 G>C | Dominant: C/G+G/G vs. G/G | 1.33 (0.84–2.11) | 0.22 | 465 |
| Recessive: C/C vs. G/G+C/G | 0.82 (0.25–2.61) | 0.73 | 466.4 | |
| Additive | 1.21 (0.81–1.79) | 0.35 | 465.6 |
| Baseline | 3-IVI | 5-IVI | Final | p-Value | |
|---|---|---|---|---|---|
| BCVA, letters | 46 ± 22 | 54 ± 21 | 69 ± 14 | 70 ± 14 | <0.001 a |
| CRT, μm | 341 ± 69 | 265 (234–306) | 234 (200–284) | 211 (190–262) | <0.001 b |
| PED, abs. % | 115 (100) | 95 (82.6) | 77 (66.9) | 72 (62.6) | |
| PED height, μm | 123 (89–167) | 45 (23–78) | 44 (23–63) | 32 (21–61) | <0.001 b |
| SRF, abs. % | 107 (93) | 80 (69.6) | 33 (28.7) | 8 (7) | |
| SRF height, μm | 67 (34–102) | 34 (23–56) | 22 (12–29) | 19 ± 8 | <0.001 b |
| IRF, abs. (%) | 89 (77.4) | 59 (51.3) | 21 (18.3) | 4 (3.5) | <0.001 c |
| IRC, abs. (%) | 52 (45.2) | 20 (17.7) | - | - | <0.001 d |
| Parameter | rs2285714 | p-Value | |||
|---|---|---|---|---|---|
| C/C | C/T | T/T | |||
| PED baseline, μm | 111 (74–127) | 126 (106–182) | 145 (100–167) | 0.018 a pC/T-C/C = 0.025 | |
| PED 3-IVI, μm | 30 (2–45) | 42 (23–78) | 45 (23–67) | 0.093 a | |
| PED 5-IVI, μm | 23 (0–35) | 28 (0–56) | 32 (0–45) | 0.265 a | |
| PED final, μm | 16 (0–33) | 21 (0–48) | 11 (0–32) | 0.549 a | |
| SRF baseline, μm | 46 (23–75) | 79 (41–118) | 67 (47–90) | 0.015 a pC/T-C/C = 0.014 | |
| SRF 3-IVI, μm | 11 (0–26) | 32 (10–56) | 34 (11–56) | 0.007 a pC/T-C/C = 0.007 | |
| SRF 5-IVI | Absence (%) | 34 (85.0) | 36 (64.3) | 10 (58.8) | 0.044 b |
| Presence (%) | 6 (15.0) | 20 (35.7) | 7 (41.2) | ||
| SRF final | Absence (%) | 39 (97.5) | 52 (92.9) | 14 (82.4) | 0.125 b |
| Presence (%) | 1 (2.5) | 4 (7.1) | 3 (17.6) | ||
| CRT baseline, μm | 340 (288–381) | 335 (292–382) | 340 (288–394) | 0.688 a | |
| CRT 3-IVI, μm | 241 (220–294) | 270 (246–310) | 270 (259–335) | 0.051 a | |
| CRT 5-IVI, μm | 228 (200–278) | 244 (221–282) | 245 (210–300) | 0.152 a | |
| CRT final, μm | 201 (190–259) | 212 (200–258) | 230 (198–279) | 0.180 a | |
| IRC initial | Absence (%) | 22 (55.0) | 31 (55.4) | 9 (52.9) | 0.985 b |
| Presence (%) | 18 (45.0) | 25 (44.6) | 8 (47.1) | ||
| IRC 3-IVI | Absence (%) | 32 (84.2) | 45 (80.4) | 14 (82.4) | 0.892 b |
| Presence (%) | 6 (15.8) | 11 (19.6) | 3 (17.6) | ||
| Anastomoses and loops final | Absence (%) | 32 (86.5) | 43 (82.7) | 10 (62.5) | 0.112 b |
| Presence (%) | 5 (13.5) | 9 (17.3) | 6 (37.5) | ||
| Active MNV final | Absence (%) | 35 (87.5) | 51 (91.1) | 12 (70.6) | 0.091 b |
| Presence (%) | 5 (12.5) | 5 (8.9) | 5 (29.4) | ||
| Parameter | rs2285714 (Dominant Model) | p-Value | ||
|---|---|---|---|---|
| C/C | C/T+T/T | |||
| PED baseline, μm | 111 (74–127) | 134 (100–179) | 0.005 a | |
| PED 3-IVI, μm | 30 (2–45) | 44 (23–78) | 0.031 a | |
| PED 5-IVI, μm | 23 (0–35) | 32 (0–56) | 0.144 a | |
| PED final, μm | 16 (0–33) | 21 (0–46) | 0.427 a | |
| SRF baseline, μm | 46 (23–75) | 78 (41–114) | 0.004 a | |
| SRF 3-IVI, μm | 11 (0–26) | 34 (11–56) | 0.002 a | |
| SRF 5-IVI | Absence (%) | 34 (85.0) | 46 (63.0) | 0.014 b |
| Presence (%) | 6 (15.0) | 27 (37.0) | ||
| SRF final | Absence (%) | 39 (97.5) | 66 (90.4) | 0.256 b |
| Presence (%) | 1 (2.5) | 7 (9.6) | ||
| CRT baseline, μm | 339 ± 77 | 343 ± 66 | 0.770 c | |
| CRT 3-IVI, μm | 241 (220–294) | 270 (247–312) | 0.022 a | |
| CRT 5-IVI, μm | 228 (200–278) | 245 (220–290) | 0.053 a | |
| CRT final, μm | 201 (190–259) | 220 (200–263) | 0.087 a | |
| IRC initial | Absence (%) | 22 (55.0) | 40 (54.8) | 0.983 b |
| Presence (%) | 18 (45.0) | 33 (45.2) | ||
| IRC 3-IVI | Absence (%) | 32 (84.2) | 59 (80.8) | 0.659 b |
| Presence (%) | 6 (15.8) | 14 (19.2) | ||
| Anastomoses and loops final | Absence (%) | 32 (86.5) | 53 (77.9) | 0.287 b |
| Presence (%) | 5 (13.5) | 15 (22.1) | ||
| Active MNV final | Absence (%) | 35 (87.5) | 63 (86.3) | 1.000 b |
| Presence (%) | 5 (12.5) | 10 (13.7) | ||
| rs2285714 (recessive model) | ||||
| C/C+C/T | T/T | |||
| PED baseline, μm | 123 (89–168) | 145 (100–167) | 0.311 a | |
| PED 3-IVI, μm | 34 (19–78) | 45 (23–67) | 0.690 a | |
| PED 5-IVI, μm | 23 (0–55) | 32 (0–45) | 0.816 a | |
| PED final, μm | 20 (0–44) | 11 (0–32) | 0.639 a | |
| SRF baseline, μm | 63 (33–96) | 67 (47–90) | 0.531 a | |
| SRF 3-IVI, μm | 23 (0–45) | 34 (11–56) | 0.395 a | |
| SRF 5-IVI | Absence (%) | 70 (72.9) | 10 (58.8) | 0.239 b |
| Presence (%) | 26 (27.1) | 7 (41.2) | ||
| SRF final | Absence (%) | 91 (94.8) | 14 (82.4) | 0.099 b |
| Presence (%) | 5 (5.2) | 3 (17.6) | ||
| CRT baseline, μm | 337 (289–381) | 340 (288–394) | 0.388 a | |
| CRT 3-IVI, μm | 264 (233–302) | 270 (259–335) | 0.133 a | |
| CRT 5-IVI, μm | 234 (201–280) | 245 (210–300) | 0.502 a | |
| CRT final, μm | 211 (190–258) | 230 (198–279) | 0.229 a | |
| IRC initial | Absence (%) | 53 (55.2) | 9 (52.9) | 1.000 b |
| Presence (%) | 43 (44.8) | 8 (47.1) | ||
| IRC 3-IVI | Absence (%) | 77 (81.9) | 14 (82.4) | 0.966 b |
| Presence (%) | 17 (18.1) | 3 (17.6) | ||
| Anastomoses and loops final | Absence (%) | 75 (84.3) | 10 (62.5) | 0.041 b |
| Presence (%) | 14 (15.7) | 6 (37.5) | ||
| Active MNV final | Absence (%) | 86 (89.6) | 12 (70.6) | 0.033 b |
| Presence (%) | 10 (10.4) | 5 (29.4) | ||
| Parameter | rs2230199 (Dominant Model) | p | ||
|---|---|---|---|---|
| C/C+C/G | G/G | |||
| PED baseline, μm | 123 (89–171) | 123 (99–167) | 0.705 a | |
| PED 3-IVI, μm | 34 (11–62) | 45 (22–78) | 0.373 a | |
| PED 5-IVI, μm | 23 (0–45) | 30 (0–56) | 0.793 a | |
| PED final, μm | 18 (0–33) | 20 (0–45) | 0.875 a | |
| SRF baseline, μm | 67 (40–90) | 63 (29–109) | 0.616 a | |
| SRF 3-IVI, μm | 23 (0–42) | 23 (0–55) | 0.510 a | |
| SRF 5-IVI | Absence (%) | 28 (70.0) | 52 (71.2) | 0.890 b |
| Presence (%) | 12 (30.0) | 21 (28.8) | ||
| SRF final | Absence (%) | 36 (90.0) | 69 (94.5) | 0.451 b |
| Presence (%) | 4 (10.0) | 4 (5.5) | ||
| CRT baseline, μm | 336 (273–405) | 283 (227–345) | 0.007 a | |
| CRT 3-IVI, μm | 282 ± 51 | 269 ± 53 | 0.220 c | |
| CRT 5-IVI, μm | 234 (208–288) | 234 (201–280) | 0.683 a | |
| CRT final, μm | 211 (200–270) | 211 (190–256) | 0.791 a | |
| IRC initial | Absence (%) | 14 (35.0%) | 48 (65.8) | 0.002 b |
| Presence (%) | 26 (65.0) | 25 (34.2) | ||
| IRC 3-IVI | Absence (%) | 28 (70.0) | 63 (88.7) | 0.014 b |
| Presence (%) | 12 (30.0) | 8 (11.3) | ||
| Anastomoses and loops final | Absence (%) | 31 (77.5) | 54 (83.1) | 0.480 b |
| Presence (%) | 9 (22.5) | 11 (16.9) | ||
| MNV final | Absence (%) | 33 (82.5) | 65 (89.0) | 0.389 b |
| Presence (%) | 7 (17.5) | 8 (11.0) | ||
| B Coefficient | Standard Error | p-Value | |
|---|---|---|---|
| CRT baseline | 0.533 | 0.049 | <0.001 |
| rs2285714: C/T | 20.867 | 7.516 | 0.007 |
| rs2285714: T/T | 24.001 | 10.652 | 0.026 |
| rs2285714: C/T+T/T (vs. C/C) | 21.570 | 7.131 | 0.003 |
| B Coefficient | Standard Error | p-Value | |
|---|---|---|---|
| MNV: type 2 | 10.480 | 4.635 | 0.026 |
| SRF height baseline | 0.412 | 0.046 | <0.001 |
| rs2285714: C/T+T/T (vs. C/C) | 9.440 | 4.658 | 0.045 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozhevnikova, O.S.; Fursova, A.Z.; Derbeneva, A.S.; Nikulich, I.F.; Tarasov, M.S.; Devyatkin, V.A.; Rumyantseva, Y.V.; Telegina, D.V.; Kolosova, N.G. Association between Polymorphisms in CFH, ARMS2, CFI, and C3 Genes and Response to Anti-VEGF Treatment in Neovascular Age-Related Macular Degeneration. Biomedicines 2022, 10, 1658. https://doi.org/10.3390/biomedicines10071658
Kozhevnikova OS, Fursova AZ, Derbeneva AS, Nikulich IF, Tarasov MS, Devyatkin VA, Rumyantseva YV, Telegina DV, Kolosova NG. Association between Polymorphisms in CFH, ARMS2, CFI, and C3 Genes and Response to Anti-VEGF Treatment in Neovascular Age-Related Macular Degeneration. Biomedicines. 2022; 10(7):1658. https://doi.org/10.3390/biomedicines10071658
Chicago/Turabian StyleKozhevnikova, Oyuna S., Anzhella Zh. Fursova, Anna S. Derbeneva, Ida F. Nikulich, Mikhail S. Tarasov, Vasiliy A. Devyatkin, Yulia V. Rumyantseva, Darya V. Telegina, and Nataliya G. Kolosova. 2022. "Association between Polymorphisms in CFH, ARMS2, CFI, and C3 Genes and Response to Anti-VEGF Treatment in Neovascular Age-Related Macular Degeneration" Biomedicines 10, no. 7: 1658. https://doi.org/10.3390/biomedicines10071658
APA StyleKozhevnikova, O. S., Fursova, A. Z., Derbeneva, A. S., Nikulich, I. F., Tarasov, M. S., Devyatkin, V. A., Rumyantseva, Y. V., Telegina, D. V., & Kolosova, N. G. (2022). Association between Polymorphisms in CFH, ARMS2, CFI, and C3 Genes and Response to Anti-VEGF Treatment in Neovascular Age-Related Macular Degeneration. Biomedicines, 10(7), 1658. https://doi.org/10.3390/biomedicines10071658