
Long-Term Systemic Treatment of a Mouse Model Displaying Chronic FSHD-like Pathology with Antisense Therapeutics That Inhibit DUX4 Expression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Antisense Chemistries
2.2. Animals
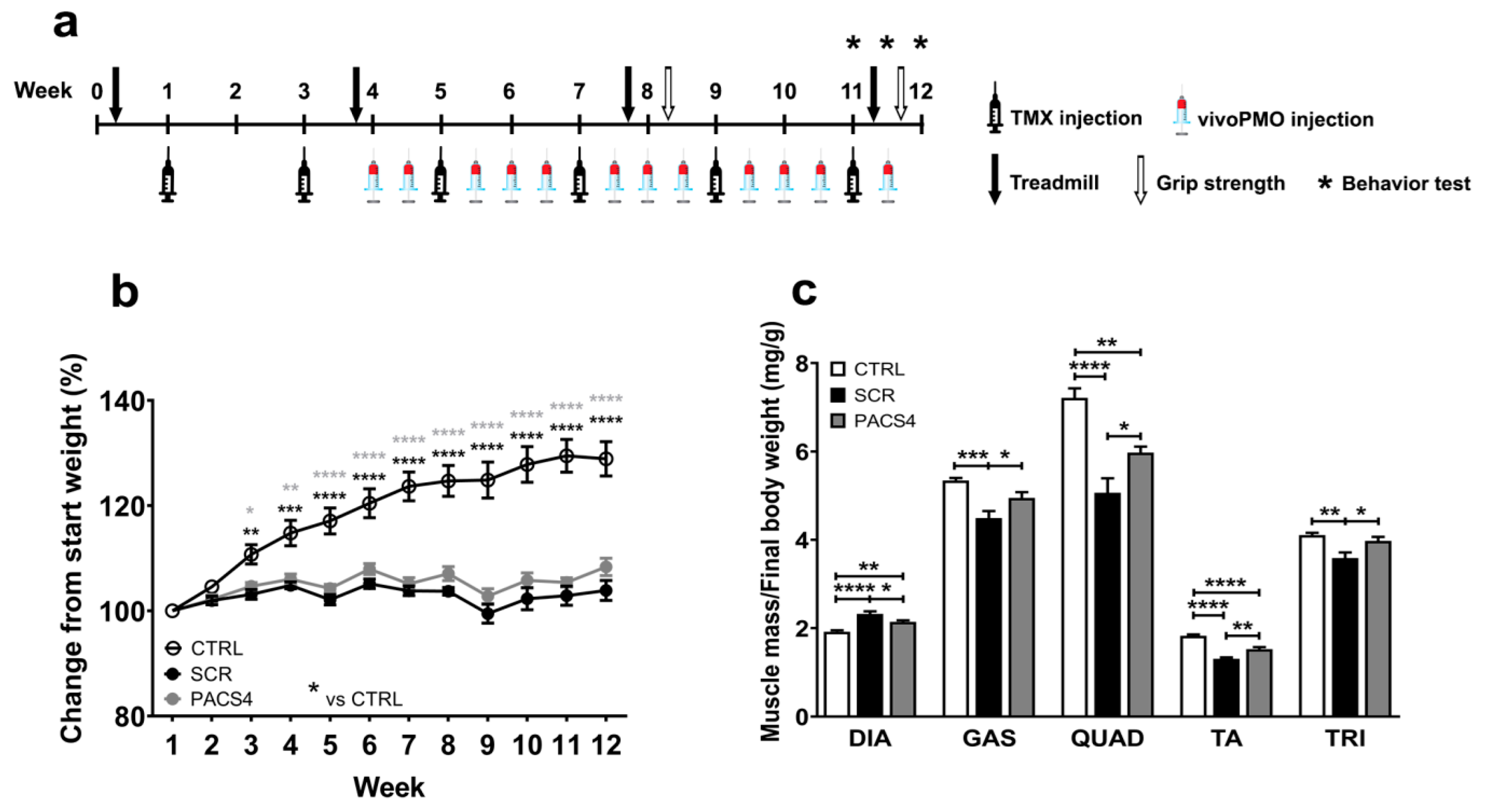
2.3. Study Design
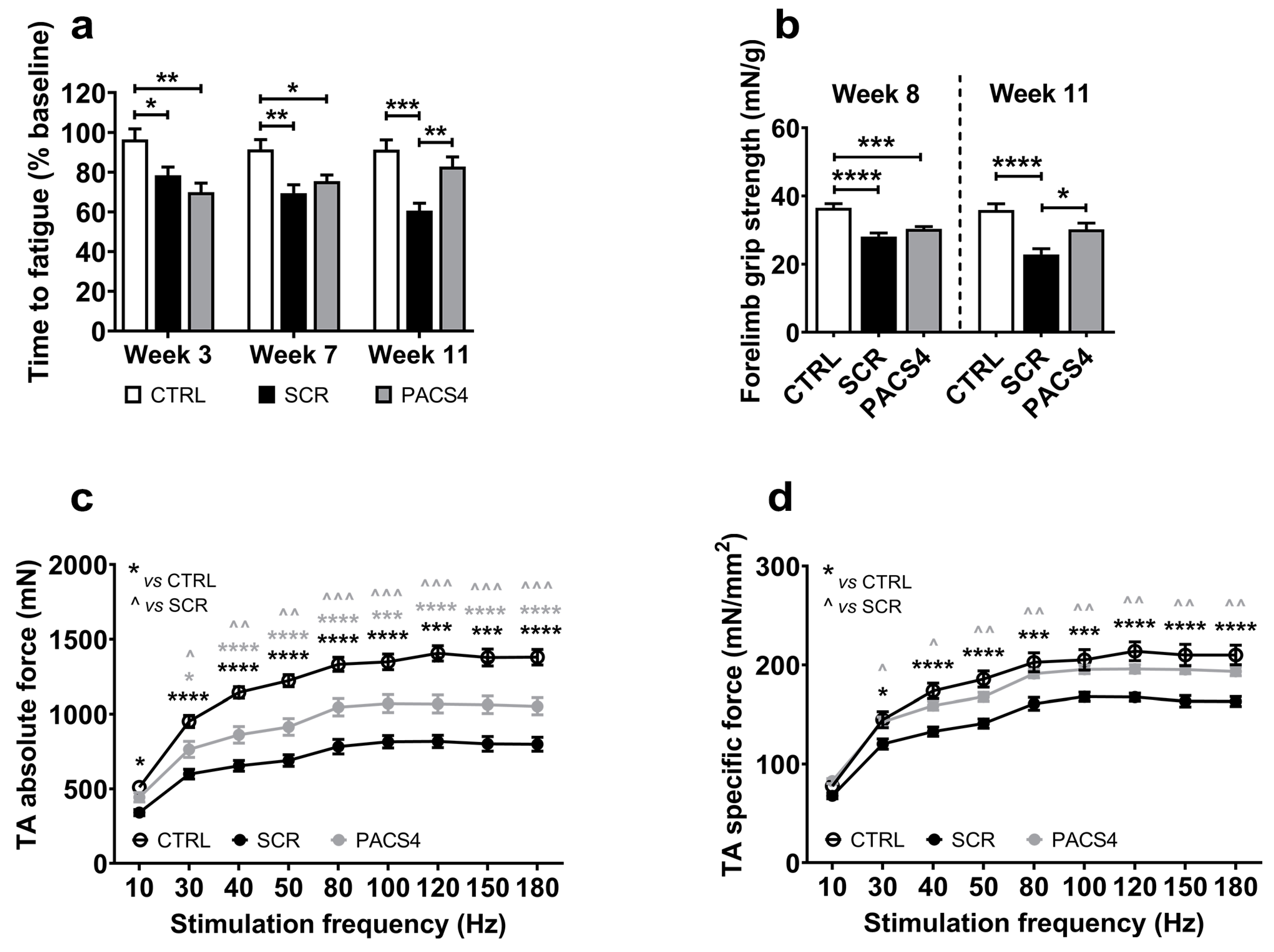
2.4. Behavioral and Functional Tests
2.5. Post-Mortem Tissue Processing
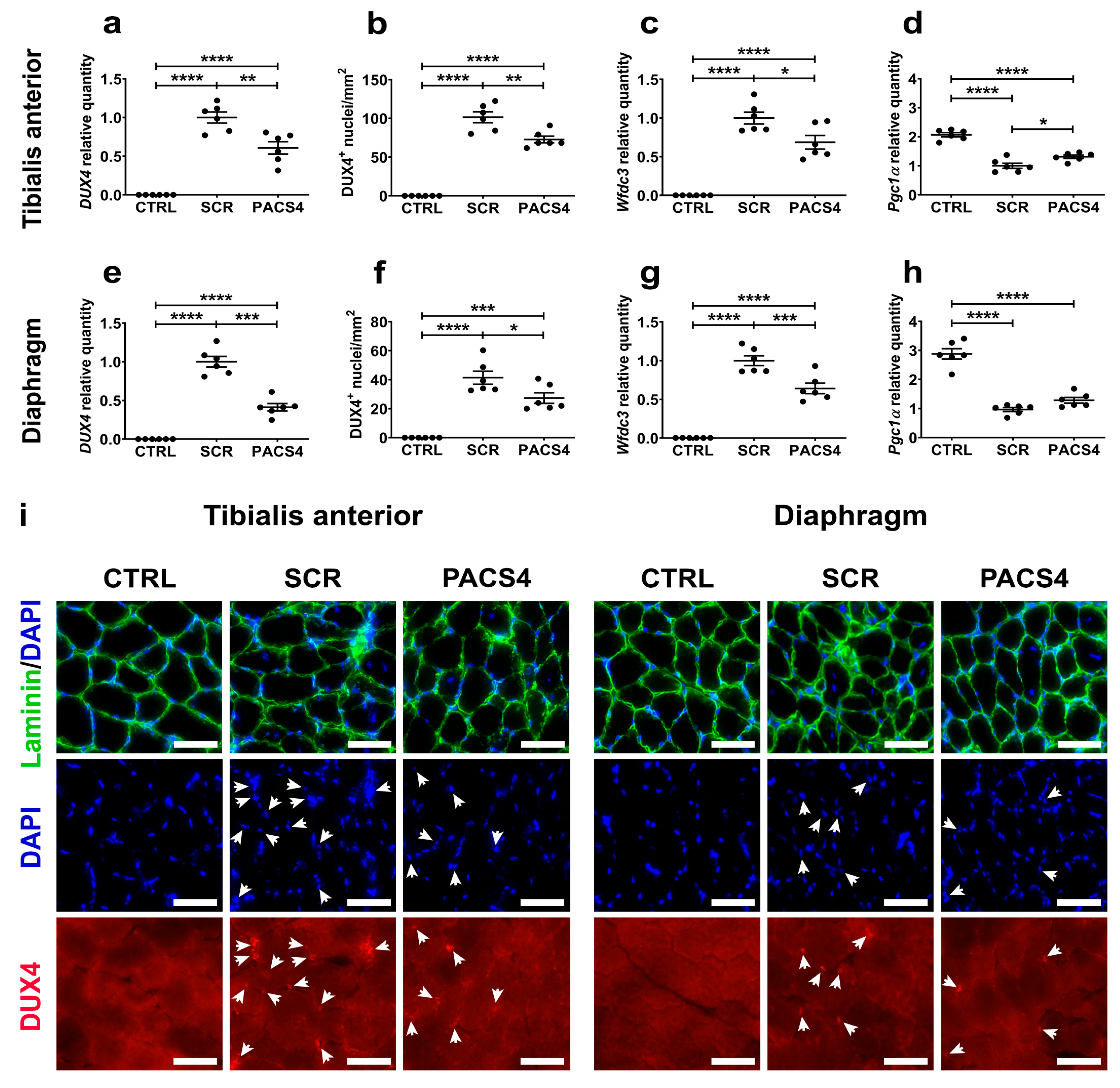
2.6. RT-qPCR Quantification for DUX4 and Relevant Genes
2.7. Immunohistochemistry
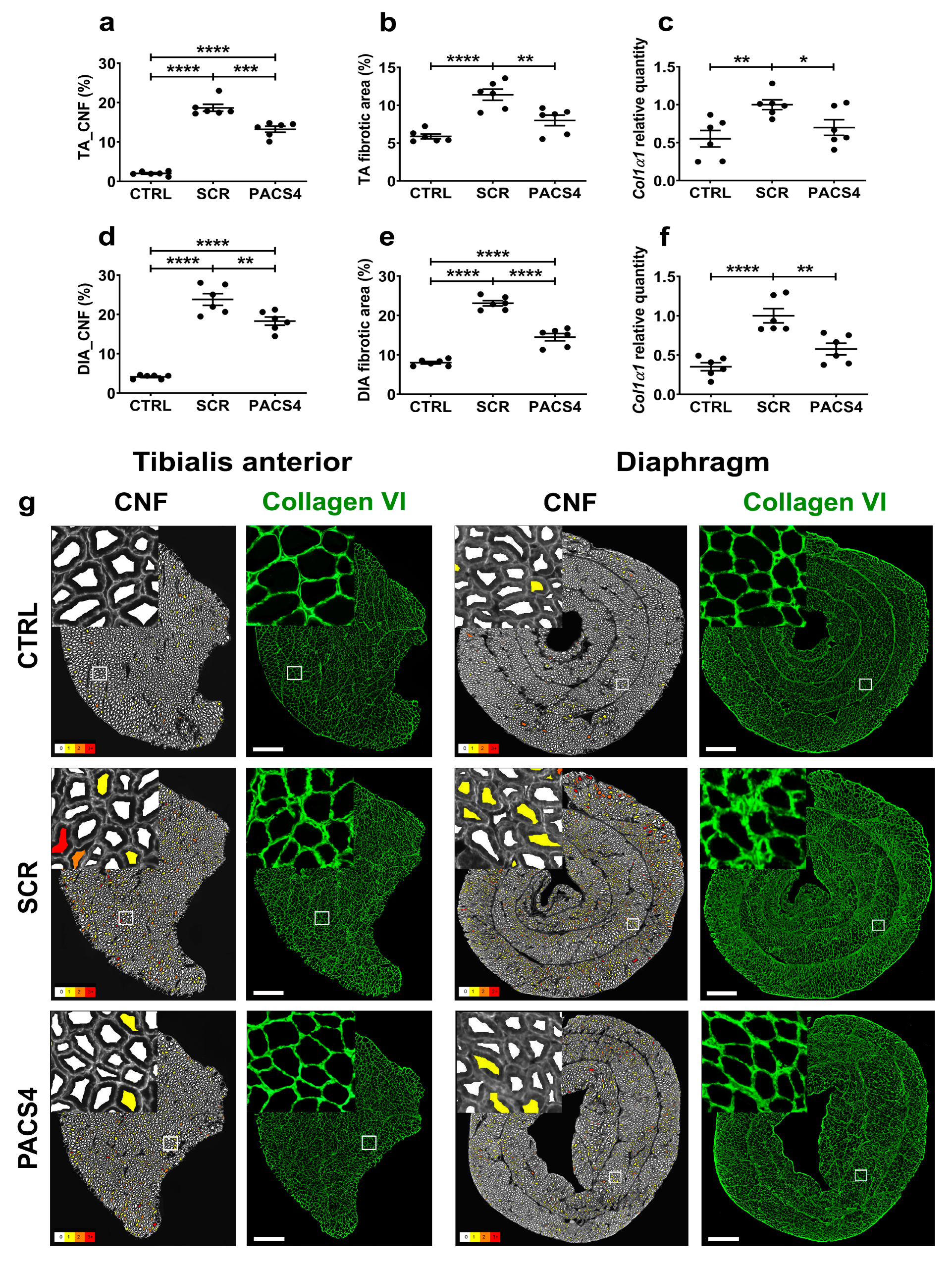
2.8. Histological Analyses
2.9. Statistical Analysis
3. Results
3.1. Systemic Treatment with vivoPMO-PACS4 Increases Muscle Mass of ACTA1-MCM/FLExDUX4 Mice
3.2. Long-Term vivoPMO-PACS4 Treatment Greatly Improves Muscle Function and Animal’s Behavior
3.3. vivoPMO-PACS4 Robustly Downregulates Expression of DUX4 and Murine DUX4-Related Targets
3.4. Long-Term vivoPMO-PACS4 Treatment Greatly Improves Muscle Histopathology
3.5. vivoPMO-PACS4 Treatment Significantly Reduces Muscle Turnover and Muscle Fibrosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mostacciuolo, M.L.; Pastorello, E.; Vazza, G.; Miorin, M.; Angelini, C.; Tomelleri, G.; Galluzzi, G.; Trevisan, C.P. Facioscapulohumeral muscular dystrophy: Epidemiological and molecular study in a north-east Italian population sample. Clin. Genet. 2009, 75, 550–555. [Google Scholar] [CrossRef]
- Deenen, J.C.; Arnts, H.; van der Maarel, S.M.; Padberg, G.W.; Verschuuren, J.J.G.M.; Bakker, E.; Weinreich, S.S.; Verbeek, A.L.; van Engelen, B.G. Population-based incidence and prevalence of facioscapulohumeral dystrophy. Neurology 2014, 83, 1056–1059. [Google Scholar] [CrossRef] [Green Version]
- Statland, J.; Tawil, R. Facioscapulohumeral Muscular Dystrophy. Neurol. Clin. 2014, 32, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Tonini, M.; Passos-Bueno, M.; Cerqueira, A.; Matioli, S.; Pavanello, R.; Zatz, M. Asymptomatic carriers and gender differences in facioscapulohumeral muscular dystrophy (FSHD). Neuromuscul. Disord. 2004, 14, 33–38. [Google Scholar] [CrossRef]
- Hamel, J.; Johnson, N.; Tawil, R.; Martens, W.B.; Dilek, N.; McDermott, M.P.; Heatwole, C. Patient-Reported Symptoms in Facioscapulohumeral Muscular Dystrophy (PRISM-FSHD). Neurology 2019, 93, e1180–e1192. [Google Scholar] [CrossRef]
- Moreira, S.; Wood, L.; Smith, D.; Marini-Bettolo, C.; Guglieri, M.; McMacken, G.; Bailey, G.; Mayhew, A.; Muni-Lofra, R.; Eglon, G.; et al. Respiratory involvement in ambulant and non-ambulant patients with facioscapulohumeral muscular dystrophy. J. Neurol. 2017, 264, 1271–1280. [Google Scholar] [CrossRef]
- Wohlgemuth, M.; Horlings, C.; van der Kooi, E.; Gilhuis, H.; Hendriks, J.; van der Maarel, S.; van Engelen, B.; Heijdra, Y.; Padberg, G. Respiratory function in facioscapulohumeral muscular dystrophy 1. Neuromuscul. Disord. 2017, 27, 526–530. [Google Scholar] [CrossRef]
- Kelly, C.R.; Saw, J.; Thapa, P.; Mandrekar, J.; Naddaf, E. Systemic manifestations and symptom burden of facioscapulohumeral muscular dystrophy in a referral cohort. Muscle Nerve 2022, 65, 415–421. [Google Scholar] [CrossRef]
- Della Marca, G.; Frusciante, R.; Dittoni, S.; Vollono, C.; Buccarella, C.; Iannaccone, E.; Rossi, M.; Scarano, E.; Pirronti, T.; Cianfoni, A.; et al. Sleep disordered breathing in facioscapulohumeral muscular dystrophy. J. Neurol. Sci. 2009, 285, 54–58. [Google Scholar] [CrossRef]
- Scully, M.A.; Eichinger, K.J.; Donlin-Smith, C.M.; Tawil, R.; Statland, J.M. Restrictive lung involvement in facioscapulohumeral muscular dystrophy. Muscle Nerve 2014, 50, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Henke, C.; Spiesshoefer, J.; Kabitz, H.-J.; Herkenrath, S.; Randerath, W.; Brix, T.; Görlich, D.; Young, P.; Boentert, M. Respiratory muscle weakness in facioscapulohumeral muscular dystrophy. Muscle Nerve 2019, 60, 679–686. [Google Scholar] [CrossRef]
- Dixit, M.; Ansseau, E.; Tassin, A.; Winokur, S.; Shi, R.; Qian, H.; Sauvage, S.; Mattéotti, C.; van Acker, A.M.; Leo, O.; et al. DUX4, a candidate gene of facioscapulohumeral muscular dystrophy, encodes a transcriptional activator of PITX1. Proc. Natl. Acad. Sci. USA 2007, 104, 18157–18162. [Google Scholar] [CrossRef] [Green Version]
- Ba, L.M.W.; Bs, S.E.G.; Mei, W.; Belayew, A.; Coppee, F.; Ladner, K.J.; Guttridge, D.; Yang, J.; Harper, S.Q. DUX4, a candidate gene for facioscapulohumeral muscular dystrophy, causes p53-dependent myopathy in vivo. Ann. Neurol. 2010, 69, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Vanderplanck, C.; Ansseau, E.; Charron, S.; Stricwant, N.; Tassin, A.; Laoudj-Chenivesse, D.; Wilton, S.; Coppée, F.; Belayew, A. The FSHD Atrophic Myotube Phenotype Is Caused by DUX4 Expression. PLoS ONE 2011, 6, e26820. [Google Scholar] [CrossRef]
- Chen, J.C.; King, O.D.; Zhang, Y.; Clayton, N.P.; Spencer, C.; Wentworth, B.M.; Emerson, C.P., Jr.; Wagner, K.R. Morpholino-mediated Knockdown of DUX4 Toward Facioscapulohumeral Muscular Dystrophy Therapeutics. Mol. Ther. 2016, 24, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.R.Q.; Maruyama, R.; Echigoya, Y.; Nguyen, Q.; Zhang, A.; Khawaja, H.; Chandra, S.S.; Jones, T.; Jones, P.; Chen, Y.-W.; et al. Inhibition of DUX4 expression with antisense LNA gapmers as a therapy for facioscapulohumeral muscular dystrophy. Proc. Natl. Acad. Sci. USA 2020, 117, 16509–16515. [Google Scholar] [CrossRef]
- Block, G.J.; Narayanan, D.; Amell, A.M.; Petek, L.M.; Davidson, K.C.; Bird, T.D.; Tawil, R.; Moon, R.T.; Miller, D.G. Wnt/β-catenin signaling suppresses DUX4 expression and prevents apoptosis of FSHD muscle cells. Hum. Mol. Genet. 2013, 22, 4661–4672. [Google Scholar] [CrossRef] [Green Version]
- Campbell, A.E.; Oliva, J.; Yates, M.P.; Zhong, J.W.; Shadle, S.C.; Snider, L.; Singh, N.; Tai, S.; Hiramuki, Y.; Tawil, R.; et al. BET bromodomain inhibitors and agonists of the beta-2 adrenergic receptor identified in screens for compounds that inhibit DUX4 expression in FSHD muscle cells. Skelet. Muscle 2017, 7, 1–18. [Google Scholar] [CrossRef]
- Cruz, J.M.; Hupper, N.; Wilson, L.S.; Concannon, J.B.; Wang, Y.; Oberhauser, B.; Patora-Komisarska, K.; Zhang, Y.; Glass, D.J.; Trendelenburg, A.-U.; et al. Protein kinase A activation inhibits DUX4 gene expression in myotubes from patients with facioscapulohumeral muscular dystrophy. J. Biol. Chem. 2018, 293, 11837–11849. [Google Scholar] [CrossRef] [Green Version]
- Wallace, L.M.; Liu, J.; Domire, J.; Garwick-Coppens, S.E.; Guckes, S.M.; Mendell, J.R.; Flanigan, K.; Harper, S.Q. RNA Interference Inhibits DUX4-induced Muscle Toxicity In Vivo: Implications for a Targeted FSHD Therapy. Mol. Ther. 2012, 20, 1417–1423. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, L.F.; Hamer, B.D.; Heuvel, A.V.D.; Franken, M.; Jackson, M.; Dwyer, C.A.; Tapscott, S.J.; Rigo, F.; van der Maarel, S.M.; de Greef, J.C. Systemic delivery of a DUX4-targeting antisense oligonucleotide to treat facioscapulohumeral muscular dystrophy. Mol. Ther. Nucleic Acids 2021, 26, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.; Galasinski, S.; Richey, A.; Campbell, A.E.; Meyers, M.; Modi, N.; Zhong, J.W.; Tawil, R.; Tapscott, S.J.; Sverdrup, F.M. Clinically Advanced p38 Inhibitors Suppress DUX4 Expression in Cellular and Animal Models of Facioscapulohumeral Muscular Dystrophy. J. Pharmacol. Exp. Ther. 2019, 370, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.Y.; Al-Kharsan, M.; Garwick-Coppens, S.E.; Chermahini, G.A.; Harper, M.A.; Palo, A.; Boudreau, R.L.; Harper, S.Q. Human miRNA miR-675 inhibits DUX4 expression and may be exploited as a potential treatment for Facioscapulohumeral muscular dystrophy. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Wallace, L.M.; Saad, N.Y.; Pyne, N.K.; Fowler, A.M.; Eidahl, J.O.; Domire, J.S.; Griffin, D.A.; Herman, A.C.; Sahenk, Z.; Rodino-Klapac, L.R.; et al. Pre-clinical Safety and Off-Target Studies to Support Translation of AAV-Mediated RNAi Therapy for FSHD. Mol. Ther. Methods Clin. Dev. 2018, 8, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himeda, C.L.; Jones, T.I.; Jones, P.L. Targeted epigenetic repression by CRISPR/dSaCas9 suppresses pathogenic DUX4-fl expression in FSHD. Mol. Ther. Methods Clin. Dev. 2021, 20, 298–311. [Google Scholar] [CrossRef]
- Ciszewski, L.; Lu-Nguyen, N.; Slater, A.; Brennan, A.; Williams, H.; Dickson, G.; Searle, M.S.; Popplewell, L. G-quadruplex ligands mediate downregulation of DUX4 expression. Nucleic Acids Res. 2020, 48, 4179–4194. [Google Scholar] [CrossRef] [Green Version]
- Rojas, L.A.; Valentine, E.; Accorsi, A.; Maglio, J.; Shen, N.; Robertson, A.; Kazmirski, S.; Rahl, P.; Tawil, R.; Cadavid, D.; et al. p38α Regulates Expression of DUX4 in a Model of Facioscapulohumeral Muscular Dystrophy. J. Pharmacol. Exp. Ther. 2020, 374, 489–498. [Google Scholar] [CrossRef]
- Mariot, V.; Joubert, R.; Marsollier, A.-C.; Hourdé, C.; Voit, T.; Dumonceaux, J. A Deoxyribonucleic Acid Decoy Trapping DUX4 for the Treatment of Facioscapulohumeral Muscular Dystrophy. Mol. Ther. Nucleic Acids 2020, 22, 1191–1199. [Google Scholar] [CrossRef]
- DeSimone, A.M.; Leszyk, J.; Wagner, K.R.; Emerson, C.P., Jr. Identification of the hyaluronic acid pathway as a therapeutic target for facioscapulohumeral muscular dystrophy. Sci. Adv. 2019, 5, eaaw7099. [Google Scholar] [CrossRef] [Green Version]
- Bosnakovski, D.; da Silva, M.T.; Sunny, S.T.; Ener, E.T.; Toso, E.A.; Yuan, C.; Cui, Z.; Walters, M.A.; Jadhav, A.; Kyba, M. A novel P300 inhibitor reverses DUX4-mediated global histone H3 hyperacetylation, target gene expression, and cell death. Sci. Adv. 2019, 5, eaaw7781. [Google Scholar] [CrossRef] [Green Version]
- Rashnonejad, A.; Amini-Chermahini, G.; Taylor, N.K.; Wein, N.; Harper, S.Q. Designed U7 snRNAs inhibit DUX4 expression and improve FSHD-associated outcomes in DUX4 overexpressing cells and FSHD patient myotubes. Mol. Ther. Nucleic Acids 2021, 23, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.R.Q.; Bittel, A.; Maruyama, R.; Echigoya, Y.; Nguyen, Q.; Huang, Y.; Dzierlega, K.; Zhang, A.; Chen, Y.-W.; Yokota, T. DUX4 Transcript Knockdown with Antisense 2′-O-Methoxyethyl Gapmers for the Treatment of Facioscapulohumeral Muscular Dystrophy. Mol. Ther. 2020, 29, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Lu-Nguyen, N.; Malerba, A.; Herath, S.; Dickson, G.; Popplewell, L. Systemic antisense therapeutics inhibiting DUX4 expression ameliorates FSHD-like pathology in an FSHD mouse model. Hum. Mol. Genet. 2021, 30, 1398–1412. [Google Scholar] [CrossRef] [PubMed]
- Lu-Nguyen, N.; Malerba, A.; Pineda, M.M.A.; Dickson, G.; Popplewell, L.J. Improving Molecular and Histopathology in Diaphragm Muscle of the Double Transgenic ACTA1-MCM/FLExDUX4 Mouse Model of FSHD with Systemic Antisense Therapy. Hum. Gene Ther. 2022; (online ahead of print). [Google Scholar] [CrossRef]
- Jagannathan, S.; de Greef, J.C.; Hayward, L.J.; Yokomori, K.; Gabellini, D.; Mul, K.; Sacconi, S.; Arjomand, J.; Kinoshita, J.; Harper, S.Q. Meeting report: The 2021 FSHD International Research Congress. Skelet. Muscle 2022, 12, 1–10. [Google Scholar] [CrossRef]
- Jones, T.I.; Chew, G.-L.; Barraza-Flores, P.; Schreier, S.; Ramirez, M.; Wuebbles, R.D.; Burkin, D.J.; Bradley, R.K.; Jones, P.L. Transgenic mice expressing tunable levels of DUX4 develop characteristic facioscapulohumeral muscular dystrophy-like pathophysiology ranging in severity. Skelet. Muscle 2020, 10, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.; Jones, P.L. A cre-inducible DUX4 transgenic mouse model for investigating facioscapulohumeral muscular dystrophy. PLoS ONE 2018, 13, e0192657. [Google Scholar] [CrossRef] [Green Version]
- Lu-Nguyen, N.; Ferry, A.; Schnell, F.J.; Hanson, G.J.; Popplewell, L.; Dickson, G.; Malerba, A. Functional muscle recovery following dystrophin and myostatin exon splice modulation in aged mdx mice. Hum. Mol. Genet. 2019, 28, 3091–3100. [Google Scholar] [CrossRef]
- Terry, R.L.; Wells, D.J. Histopathological Evaluation of Skeletal Muscle with Specific Reference to Mouse Models of Muscular Dystrophy. Curr. Protoc. Mouse Biol. 2016, 6, 343–363. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Mayeuf-Louchart, A.; Hardy, D.; Thorel, Q.; Roux, P.; Gueniot, L.; Briand, D.; Mazeraud, A.; Bouglé, A.; Shorte, S.L.; Staels, B.; et al. MuscleJ: A high-content analysis method to study skeletal muscle with a new Fiji tool. Skelet. Muscle 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Razali, N.M.; Wah, Y.B. Power Comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling Tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]
- Banerji, C.R.S.; Henderson, D.; Tawil, R.N.; Zammit, P.S. Skeletal muscle regeneration in facioscapulohumeral muscular dystrophy is correlated with pathological severity. Hum. Mol. Genet. 2020, 29, 2746–2760. [Google Scholar] [CrossRef] [PubMed]
- Lassche, S.; Küsters, B.; Heerschap, A.; Schyns, M.V.; Ottenheijm, C.A.; Voermans, N.C.; van Engelen, B.G. Correlation Between Quantitative MRI and Muscle Histopathology in Muscle Biopsies from Healthy Controls and Patients with IBM, FSHD and OPMD. J. Neuromuscul. Dis. 2020, 7, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Boldrin, L.; Ross, J.A.; Whitmore, C.; Doreste, B.; Beaver, C.; Eddaoudi, A.; Pearce, D.J.; Morgan, J.E. The effect of calorie restriction on mouse skeletal muscle is sex, strain and time-dependent. Sci. Rep. 2017, 7, 5160. [Google Scholar] [CrossRef] [Green Version]
- de Greef, J.; Hamlyn, R.; Jensen, B.S.; Landa, R.O.; Levy, J.R.; Kobuke, K.; Campbell, K.P. Collagen VI deficiency reduces muscle pathology, but does not improve muscle function, in the γ-sarcoglycan-null mouse. Hum. Mol. Genet. 2016, 25, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.M.; Ramirez, M.; Jones, T.I.; Jones, P.L. Identification of candidate miRNA biomarkers for facioscapulohumeral muscular dystrophy using DUX4-based mouse models. Dis. Model. Mech. 2021, 14, dmm049016. [Google Scholar] [CrossRef]
- Bosnakovski, D.; Chan, S.S.K.; Recht, O.O.; Hartweck, L.M.; Gustafson, C.J.; Athman, L.L.; Lowe, D.A.; Kyba, M. Muscle pathology from stochastic low level DUX4 expression in an FSHD mouse model. Nat. Commun. 2017, 8, 550. [Google Scholar] [CrossRef]
- Giesige, C.R.; Wallace, L.M.; Heller, K.N.; Eidahl, J.O.; Saad, N.Y.; Fowler, A.M.; Pyne, N.K.; Al-Kharsan, M.; Rashnonejad, A.; Chermahini, G.A.; et al. AAV-mediated follistatin gene therapy improves functional outcomes in the TIC-DUX4 mouse model of FSHD. JCI Insight 2018, 3, e123538. [Google Scholar] [CrossRef]
- Chen, T.-H.; Lai, Y.-H.; Lee, P.-L.; Hsu, J.-H.; Goto, K.; Hayashi, Y.K.; Nishino, I.; Lin, C.-W.; Shih, H.-H.; Huang, C.-C.; et al. Infantile facioscapulohumeral muscular dystrophy revisited: Expansion of clinical phenotypes in patients with a very short EcoRI fragment. Neuromuscul. Disord. 2013, 23, 298–305. [Google Scholar] [CrossRef]
- Tassin, A.; Laoudj-Chenivesse, D.; Vanderplanck, C.; Barro, M.; Charron, S.; Ansseau, E.; Chen, Y.-W.; Mercier, J.; Coppée, F.; Belayew, A. DUX4 expression in FSHD muscle cells: How could such a rare protein cause a myopathy? J. Cell. Mol. Med. 2012, 17, 76–89. [Google Scholar] [CrossRef]
- Charleston, J.S.; Schnell, F.J.; Dworzak, J.; Donoghue, C.; Lewis, S.; Chen, L.; Young, G.D.; Milici, A.J.; Voss, J.; Dealwis, U.; et al. Eteplirsen treatment for Duchenne muscular dystrophy. Neurology 2018, 90, e2146–e2154. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Accession Number | Primer Sequence (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| DUX4 | Gene ID: 100288687 | Forward: CTCTGTGCCCTTGTTCTTC Reverse: TCCAGGAGATGTAACTCTAATCCA | 98 |
| Gapdh | NM_008084 | Forward: TCCATGACAACTTTGGCATTG Reverse: TCACGCCACAGCTTTCCA | 103 |
| Col1α1 | NM_007742 | Forward: GAAACTTTGCTTCCCAGATGTC Reverse: AGACCACGAGGACCAGAA | 94 |
| Pgc1α | NM_008904 | Forward: GGCACCTGAACAGAACGAAC Reverse: CAACAGGCATCAGCAGTGTC | 180 |
| Wfdc3 | NM_027961 | Forward: CTTCCATGTCAGGAGCTGTG Reverse: ACCAGGATTCTGGGACATTG | 134 |
| Open-Field Cage Activity | CTRL | SCR | PACS4 | SCR vs. WT | PACS4 vs. WT | PACS4 vs. SCR | |
|---|---|---|---|---|---|---|---|
| Parameter | Description | Mean ± SEM | Mean ± SEM | Mean ± SEM | p Value | p Value | p Value |
| Total activity | Total beam breaks | 840 ± 235 | 361 ± 51 | 565 ± 52 | 0.0208 * | 0.1976 | 0.0085 ** |
| Fast activity | Fast beam breaks | 87 ± 30 | 25 ± 4 | 62 ± 7 | 0.0209 * | 0.4994 | 0.1803 |
| Slow activity | Slow beam breaks | 753 ± 206 | 336 ± 53 | 503 ± 50 | 0.0303 * | 0.2209 | 0.5601 |
| Total static counts | Total beam breaks, movement lower than mobile threshold | 636 ± 160 | 324 ± 52 | 441 ± 47 | 0.0449 * | 0.2829 | 0.5705 |
| Fast static counts | Beam breaks, movement lower than mobile threshold and faster than fast threshold | 27 ± 8 | 12 ± 3 | 20 ± 4 | 0.1024 | 0.6527 | 0.3918 |
| Slow static count | Beam breaks, movement lower than mobile threshold and slower than fast threshold | 609 ± 152 | 312 ± 50 | 421 ± 43 | 0.0438 * | 0.2690 | 0.5849 |
| Total mobile counts | Total beam breaks, movement greater than mobile threshold | 204 ± 79 | 48 ± 6 | 113 ± 11 | 0.0169 * | 0.2257 | 0.4024 |
| Fast mobile counts | Beam breaks, movement greater than mobile threshold and faster than fast threshold | 60 ± 23 | 14 ± 2 | 41 ± 5 | 0.0187 * | 0.4972 | 0.1662 |
| Slow mobile counts | Beam breaks, movement greater than mobile threshold and slower than fast threshold | 144 ± 57 | 34 ± 4 | 72 ± 8 | 0.0191 * | 0.1607 | 0.5547 |
| Total rearing counts | Number of rearing beam breaks | 242 ± 61 | 86 ± 20 | 247 ± 29 | 0.0199 * | 0.9640 | 0.0050 ** |
| Fast rearing counts | Number of fast rearing beam breaks | 91 ± 19 | 49 ± 15 | 129 ± 15 | 0.1875 | 0.2508 | 0.0019 ** |
| Slow rearing counts | Number of slow rearing beam breaks | 151 ± 37 | 37 ± 7 | 118 ± 14 | 0.0007 *** | 0.4825 | 0.0091 ** |
| Total center rearing counts | Number of rearing beam breaks occurring away from the cage walls | 79 ± 23 | 19 ± 6 | 70 ± 17 | 0.0290 * | 0.9171 | 0.0457 * |
| Fast center rearing counts | Number of fast rearing beam breaks occurring away from the cage walls | 34 ± 12 | 12 ± 5 | 47 ± 16 | 0.4262 | 0.7782 | 0.0985 |
| Slow center rearing counts | Number of slow rearing beam breaks occurring away from the cage walls | 45 ± 14 | 7 ± 2 | 23 ± 5 | 0.0030 ** | 0.1321 | 0.2268 |
| Active time | Time of mobile or static activity (s) | 585 ± 143 | 282 ± 42 | 435 ± 45 | 0.0286 * | 0.3816 | 0.3216 |
| Static time | Time of static activity (s) | 489 ± 112 | 252 ± 40 | 369 ± 39 | 0.0397 * | 0.3984 | 0.3802 |
| Mobile time | Time of mobile activity (s) | 96 ± 34 | 30 ± 4 | 66 ± 7 | 0.0222 * | 0.4175 | 0.2468 |
| Rearing time | Time spent rearing (s) | 570 ± 170 | 148 ± 31 | 449 ± 53 | 0.0062 ** | 0.6193 | 0.0392 * |
| Front to back counts | Number of traverses from front to back | 50 ± 15 | 14 ± 2 | 33 ± 3 | 0.0054 ** | 0.2534 | 0.1697 |
| Inactive time | Time spent in inactivity (s) | 2992 ± 153 | 3290 ± 49 | 3166 ± 45 | 0.0382 * | 0.3090 | 0.4704 |
| Distance travelled | Total distance travelled (m) | 35 ± 8 | 17 ± 3 | 31 ± 3 | 0.0400 * | 0.8487 | 0.0904 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu-Nguyen, N.; Dickson, G.; Malerba, A.; Popplewell, L. Long-Term Systemic Treatment of a Mouse Model Displaying Chronic FSHD-like Pathology with Antisense Therapeutics That Inhibit DUX4 Expression. Biomedicines 2022, 10, 1623. https://doi.org/10.3390/biomedicines10071623
Lu-Nguyen N, Dickson G, Malerba A, Popplewell L. Long-Term Systemic Treatment of a Mouse Model Displaying Chronic FSHD-like Pathology with Antisense Therapeutics That Inhibit DUX4 Expression. Biomedicines. 2022; 10(7):1623. https://doi.org/10.3390/biomedicines10071623
Chicago/Turabian StyleLu-Nguyen, Ngoc, George Dickson, Alberto Malerba, and Linda Popplewell. 2022. "Long-Term Systemic Treatment of a Mouse Model Displaying Chronic FSHD-like Pathology with Antisense Therapeutics That Inhibit DUX4 Expression" Biomedicines 10, no. 7: 1623. https://doi.org/10.3390/biomedicines10071623
APA StyleLu-Nguyen, N., Dickson, G., Malerba, A., & Popplewell, L. (2022). Long-Term Systemic Treatment of a Mouse Model Displaying Chronic FSHD-like Pathology with Antisense Therapeutics That Inhibit DUX4 Expression. Biomedicines, 10(7), 1623. https://doi.org/10.3390/biomedicines10071623