
The Impact of Inflammatory Stimuli on Xylosyltransferase-I Regulation in Primary Human Dermal Fibroblasts

 , ,
, ,
Abstract
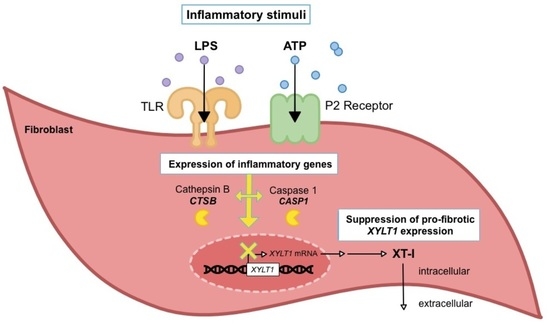
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials and Reagent Preparation
2.2. Primary Cell Culture
2.3. Cell Treatment and Sample Preparation
2.4. Cell Proliferation Assay
2.5. BCA Assay
2.6. XT-I Activity Determination by Mass Spectrometry
2.7. RNA Extraction and cDNA Synthesis
2.8. QRT-PCR Analysis
2.9. Statistical Analysis
3. Results
3.1. Time- and Concentration-Dependent Decrease of XYLT1 mRNA-Expression by LPS in Primary Skin Fibroblasts
3.2. Differences in ATP- and LPS-Induced Effects on the XYLT1 mRNA-Expression and XT-I Activity of Primary Skin Fibroblasts
3.3. CASP1 and CTSB Are Negative Regulators of XYLT1 mRNA Expression in Primary Skin Fibroblasts
3.4. PXE Fibroblasts Exhibit a Nonsignificant Reduction in XYLT1 mRNA Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Mack, M. Inflammation and fibrosis. Matrix Biol. 2018, 68–69, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Hernández, L.A.; Gómez-Olivares, J.L.; Buentello-Volante, B.; Bautista-de Lucio, V.M. Fibroblasts: The unknown sentinels eliciting immune responses against microorganisms. Eur. J. Microbiol. Immunol. 2017, 7, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.G.; Soares, D.G.; Pansani, T.N.; Turrioni, A.P.S.; Scheffel, D.L.; de Souza Costa, C.A.; Hebling, J. Effect of LPS treatment on the viability and chemokine synthesis by epithelial cells and gingival fibroblasts. Arch. Oral Biol. 2015, 60, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Gvirtz, R.; Ogen-Shtern, N.; Cohen, G. Kinetic cytokine secretion profile of LPS-induced inflammation in the human skin organ culture. Pharmaceutics 2020, 12, 299. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.-D.; Kleine, A.; Plümers, R.; Fischer, B.; Schmidt, V.; Hendig, D. Cytokine-mediated induction of human xylosyltransferase-I in systemic sclerosis skin fibroblasts. Biochem. Biophys. Res. Commun. 2021, 549, 34–39. [Google Scholar] [CrossRef]
- Sp, N.; Kang, D.Y.; Kim, H.D.; Rugamba, A.; Jo, E.S.; Park, J.-C.; Bae, S.W.; Lee, J.-M.; Jang, K.-J. Natural sulfurs inhibit LPS-induced inflammatory responses through NF-ΚB signaling in CCD-986Sk skin fibroblasts. Life 2021, 11, 427. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, P.; Ma, X.; Yin, X.; Li, J.; Wang, H.; Jiang, W.; Jia, Q.; Ni, L. Mechanisms that lead to the regulation of NLRP3 inflammasome expression and activation in human dental pulp fibroblasts. Mol. Immunol. 2015, 66, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.; Meade, K.G.; O’Farrelly, C. Non-canonical inflammasome-mediated IL-1β production by primary endometrial epithelial and stromal fibroblast cells is NLRP3 and caspase-4 dependent. Front. Immunol. 2019, 10, 102. [Google Scholar] [CrossRef]
- Shalhoub, J.; Falck-Hansen, M.A.; Davies, A.H.; Monaco, C. Innate immunity and monocyte-macrophage activation in atherosclerosis. J. Inflamm. 2011, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Roebuck, K.A. Regulation of interleukin-8 gene expression. J. Interferon Cytokine Res. 1999, 19, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Vidak, E.; Javoršek, U.; Vizovišek, M.; Turk, B. Cysteine cathepsins and their extracellular roles: Shaping the microenvironment. Cells 2019, 8, 264. [Google Scholar] [CrossRef]
- Mizunoe, Y.; Kobayashi, M.; Hoshino, S.; Tagawa, R.; Itagawa, R.; Hoshino, A.; Okita, N.; Sudo, Y.; Nakagawa, Y.; Shimano, H.; et al. Cathepsin B overexpression induces degradation of perilipin 1 to cause lipid metabolism dysfunction in adipocytes. Sci. Rep. 2020, 10, 634. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; Ni, J.; Liu, Y.; Meng, J.; Yu, W.; Nakanishi, H.; Zhou, Y. Cathepsin B regulates collagen expression by fibroblasts via prolonging TLR2/NF- κ B activation. Oxid. Med. Cell. Longev. 2016, 2016, 7894247. [Google Scholar] [CrossRef] [PubMed]
- Mort, J.S.; Magny, M.-C.; Lee, E.R. Cathepsin B: An alternative protease for the generation of an aggrecan ‘metalloproteinase’ cleavage neoepitope. Biochem. J. 1998, 335, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Bien, S.; Ritter, C.A.; Gratz, M.; Sperker, B.; Sonnemann, J.; Beck, J.F.; Kroemer, H.K. Nuclear factor-ΚB mediates up-regulation of cathepsin B by doxorubicin in tumor cells. Mol. Pharmacol. 2004, 65, 1092–1102. [Google Scholar] [CrossRef]
- De Mingo, Á.; de Gregorio, E.; Moles, A.; Tarrats, N.; Tutusaus, A.; Colell, A.; Fernandez-Checa, J.C.; Morales, A.; Marí, M. Cysteine cathepsins control hepatic NF-ΚB-dependent inflammation via sirtuin-1 regulation. Cell Death Dis. 2016, 7, e2464. [Google Scholar] [CrossRef] [PubMed]
- Bruchard, M.; Mignot, G.; Derangère, V.; Chalmin, F.; Chevriaux, A.; Végran, F.; Boireau, W.; Simon, B.; Ryffel, B.; Connat, J.L.; et al. Chemotherapy-triggered cathepsin B release in myeloid-derived suppressor cells activates the Nlrp3 inflammasome and promotes tumor growth. Nat. Med. 2013, 19, 57–64. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Kakegawa, H.; Narita, Y.; Hachiya, Y.; Hayakawa, T.; Kos, J.; Turk, V.; Katunuma, N. Significance of cathepsin B accumulation in synovial fluid of rheumatoid arthritis. Biochem. Biophys. Res. Commun. 2001, 283, 334–339. [Google Scholar] [CrossRef]
- Götting, C.; Kuhn, J.; Kleesiek, K. Human xylosyltransferases in health and disease. Cell. Mol. Life Sci. 2007, 64, 1498–1517. [Google Scholar] [CrossRef]
- Koslowski, R.; Pfeil, U.; Fehrenbach, H.; Kasper, M.; Skutelsky, E.; Wenzel, K.-W. Changes in xylosyltransferase activity and in proteoglycan deposition in bleomycin-induced lung injury in rat. Eur. Respir. J. 2001, 18, 347–356. [Google Scholar] [CrossRef]
- Faust, I.; Roch, C.; Kuhn, J.; Prante, C.; Knabbe, C.; Hendig, D. Human xylosyltransferase-I—A new marker for myofibroblast differentiation in skin fibrosis. Biochem. Biophys. Res. Commun. 2013, 436, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Götting, C.; Kuhn, J.; Sollberg, S.; Huerkamp, C.; Brinkmann, T.; Krieg, T.; Kleesiek, K. Elevated serum xylosyltransferase activity correlates with a high level of hyaluronate in patients with systemic sclerosis. Acta Derm. Venereol. 2000, 80, 60–61. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hwang, H.S.; Lee, M.H.; Kim, H.A. Fibronectin fragment inhibits xylosyltransferase-1 expression by regulating Sp1/Sp3- dependent transcription in articular chondrocytes. Osteoarthr. Cartil. 2019, 27, 833–843. [Google Scholar] [CrossRef]
- Robert, S.; Gicquel, T.; Victoni, T.; Valença, S.; Barreto, E.; Bailly-Maître, B.; Boichot, E.; Lagente, V. Involvement of matrix metalloproteinases (MMPs) and inflammasome pathway in molecular mechanisms of fibrosis. Biosci. Rep. 2016, 36, e00360. [Google Scholar] [CrossRef] [PubMed]
- Artlett, C.M. Inflammasomes in wound healing and fibrosis: Inflammasomes in wound healing and fibrosis. J. Pathol. 2013, 229, 157–167. [Google Scholar] [CrossRef]
- Yang, H.-Z.; Wang, J.-P.; Mi, S.; Liu, H.-Z.; Cui, B.; Yan, H.-M.; Yan, J.; Li, Z.; Liu, H.; Hua, F.; et al. TLR4 activity is required in the resolution of pulmonary inflammation and fibrosis after acute and chronic lung injury. Am. J. Pathol. 2012, 180, 275–292. [Google Scholar] [CrossRef]
- Bitzer, M.; von Gersdorf, G.; Liang, D.; Dominguez-Rosales, A.; Beg, A.A.; Rojkind, M.; Böttinger, E.P. A mechanism of suppression of TGF–beta/SMAD signaling by NF-kappa B/RelA. Genes Dev. 2000, 14, 187–197. [Google Scholar] [CrossRef]
- Tiemann, J.; Lindenkamp, C.; Plümers, R.; Faust, I.; Knabbe, C.; Hendig, D. Statins as a therapeutic approach for the treatment of pseudoxanthoma elasticum patients: Evaluation of the spectrum efficacy of atorvastatin in vitro. Cells 2021, 10, 442. [Google Scholar] [CrossRef]
- De Vilder, E.Y.G.; Martin, L.; Lefthériotis, G.; Coucke, P.; Van Nieuwerburgh, F.; Vanakker, O.M. Rare modifier variants alter the severity of cardiovascular disease in pseudoxanthoma elasticum: Identification of novel candidate modifier genes and disease pathways through mixture of effects analysis. Front. Cell Dev. Biol. 2021, 9, 612581. [Google Scholar] [CrossRef]
- González-Dominguez, A.; Montañez, R.; Castejón-Vega, B.; Nuñez-Vasco, J.; Lendines-Cordero, D.; Wang, C.; Mbalaviele, G.; Navarro-Pando, J.M.; Alcocer-Gómez, E.; Cordero, M.D. Inhibition of the NLRP3 inflammasome improves lifespan in animal murine model of Hutchinson–Gilford progeria. EMBO Mol. Med. 2021, 13, e14012. [Google Scholar] [CrossRef]
- Sene, A.; Khan, A.A.; Cox, D.; Nakamura, R.E.I.; Santeford, A.; Kim, B.M.; Sidhu, R.; Onken, M.D.; Harbour, J.W.; Hagbi-Levi, S.; et al. Impaired cholesterol efflux in senescent macrophages promotes age-related macular degeneration. Cell Metab. 2013, 17, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Brampton, C.; Pomozi, V.; Chen, L.-H.; Apana, A.; McCurdy, S.; Zoll, J.; Boisvert, W.A.; Lambert, G.; Henrion, D.; Blanchard, S.; et al. ABCC6 deficiency promotes dyslipidemia and atherosclerosis. Sci. Rep. 2021, 11, 3881. [Google Scholar] [CrossRef] [PubMed]
- Hannaford, J.; Guo, H.; Chen, X. Involvement of cathepsins B and L in inflammation and cholesterol trafficking protein NPC2 secretion in macrophages: Cathepsin activity influences TNF-α and NPC2. Obesity 2013, 21, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.-D.; Plümers, R.; Fischer, B.; Schmidt, V.; Hendig, D.; Kuhn, J.; Knabbe, C.; Faust, I. Activin A-mediated regulation of XT-I in human skin fibroblasts. Biomolecules 2020, 10, 609. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Tardif, F.; Ross, G.; Rouabhia, M. Gingival and dermal fibroblasts produce interleukin-1b converting enzyme and interleukin-1b but not interleukin-18 even after stimulation with lipopolysaccharide. J. Cell. Physiol. 2004, 198, 125–132. [Google Scholar] [CrossRef]
- Eleftheriadis, T.; Liakopoulos, V.; Lawson, B.; Antoniadi, G.; Stefanidis, I.; Galaktidou, G. Lipopolysaccharide and hypoxia significantly alters interleukin-8 and macrophage chemoattractant protein-1 production by human fibroblasts but not fibrosis related factors. Hippokratia 2011, 15, 238–243. [Google Scholar]
- Tang, X.; Liu, J.; Dong, W.; Li, P.; Li, L.; Hou, J.; Zheng, Y.; Lin, C.; Ren, J. Protective effect of kaempferol on LPS plus ATP-induced inflammatory response in cardiac fibroblasts. Inflammation 2015, 38, 94–101. [Google Scholar] [CrossRef]
- Yang, H.; Hu, C.; Li, F.; Liang, L.; Liu, L. Effect of lipopolysaccharide on the biological characteristics of human skin fibroblasts and hypertrophic scar tissue formation. IUBMB Life 2013, 65, 526–532. [Google Scholar] [CrossRef]
- Pönighaus, C.; Kuhn, J.; Kleesiek, K.; Götting, C. Involvement of a cysteine protease in the secretion process of Human xylosyltransferase I. Glycoconj. J. 2010, 27, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Solini, A.; Chiozzi, P.; Morelli, A.; Adinolfi, E.; Rizzo, R.; Baricordi, O.R.; Di Virgilio, F. Enhanced P2X7 activity in human fibroblasts from diabetic patients: A possible pathogenetic mechanism for vascular damage in diabetes. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Guo, Y.; Liu, Y.; Shi, B.; Guo, X.; Jin, L.; Yan, S. The in vitro effect of lipopolysaccharide on proliferation, inflammatory factors and antioxidant enzyme activity in bovine mammary epithelial cells. Anim. Nutr. 2016, 2, 99–104. [Google Scholar] [CrossRef]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakae, H.; Murakami, K.; Miyake, Y.; Matsuo, T. CXCL12 and CXCR4 expression by human gingival fibroblasts in periodontal disease. Clin. Exp. Immunol. 2005, 141, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, N.; Fukuda, K.; Fujitsu, Y.; Lu, Y.; Chikamoto, N.; Nishida, T. Lipopolysaccharide-induced expression of intercellular adhesion molecule-1 and chemokines in cultured human corneal fibroblasts. Investig. Opthalmol. Vis. Sci. 2005, 46, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.; Wan, B.; Wei, Z.; Wang, T.; Zhao, P.; Dou, Y.; Lv, Z.; Xia, Y.; Dai, Y. Role of cathepsin B in regulating migration and invasion of fibroblast-like synoviocytes into inflamed tissue from patients with rheumatoid arthritis. Clin. Exp. Immunol. 2014, 177, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Hentze, H.; Lin, X.Y.; Choi, M.S.K.; Porter, A.G. Critical role for cathepsin B in mediating caspase-1-dependent interleukin-18 maturation and caspase-1-independent necrosis triggered by the microbial toxin nigericin. Cell Death Differ. 2003, 10, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Prante, C.; Milting, H.; Kassner, A.; Farr, M.; Ambrosius, M.; Schön, S.; Seidler, D.G.; Banayosy, A.E.; Körfer, R.; Kuhn, J.; et al. Transforming growth factor Β1-regulated xylosyltransferase I activity in human cardiac fibroblasts and its impact for myocardial remodeling. J. Biol. Chem. 2007, 282, 26441–26449. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, A.; Reichert, S.N.A.; Südkamp, N.P.; Hernandez, S.L.; Nerlich, A.G.; Kühle, J.; Mayr, H.O. Expression of xylosyltransferases I and II and their role in the pathogenesis of arthrofibrosis. J. Orthop. Surg. 2020, 15, 27. [Google Scholar] [CrossRef]
- Buechler, M.B.; Pradhan, R.N.; Krishnamurty, A.T.; Cox, C.; Calviello, A.K.; Wang, A.W.; Yang, Y.A.; Tam, L.; Caothien, R.; Roose-Girma, M.; et al. Cross-tissue organization of the fibroblast lineage. Nature 2021, 593, 575–579. [Google Scholar] [CrossRef]
- Korsunsky, I.; Wei, K.; Pohin, M.; Kim, E.Y.; Barone, F.; Kang, J.B.; Friedrich, M.; Turner, J.; Nayar, S.; Fisher, B.A.; et al. Cross-tissue, single-cell stromal atlas identifies shared pathological fibroblast phenotypes in four chronic inflammatory diseases. Med 2022, S2666–S6340. [Google Scholar] [CrossRef] [PubMed]
- Müller, B.; Prante, C.; Kleesiek, K.; Götting, C. Identification and characterization of the human xylosyltransferase I gene promoter region. J. Biol. Chem. 2009, 284, 30775–30782. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.C.; Kuhn, J.; Kleesiek, K.; Götting, C. Heterologous expression and biochemical characterization of soluble human xylosyltransferase II. Biochem. Biophys. Res. Commun. 2008, 365, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Terada, K.; Yamada, J.; Hayashi, Y.; Wu, Z.; Uchiyama, Y.; Peters, C.; Nakanishi, H. Involvement of cathepsin B in the processing and secretion of interleukin-1β in chromogranin A-atimulated microglia. Glia 2010, 58, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, Z.; Hayashi, Y.; Peters, C.; Tsuda, M.; Inoue, K.; Nakanishi, H. Microglial cathepsin B contributes to the initiation of peripheral inflammation-induced chronic pain. J. Neurosci. 2012, 32, 11330–11342. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sun, L.; Hashioka, S.; Yu, S.; Schwab, C.; Okada, R.; Hayashi, Y.; McGeer, P.L.; Nakanishi, H. Differential pathways for interleukin-1β production activated by chromogranin A and amyloid β in microglia. Neurobiol. Aging 2013, 34, 2715–2725. [Google Scholar] [CrossRef]
- Faust, I.; Donhauser, E.; Fischer, B.; Ibold, B.; Kuhn, J.; Knabbe, C.; Hendig, D. Characterization of dermal myofibroblast differentiation in pseudoxanthoma elasticum. Exp. Cell Res. 2017, 360, 153–162. [Google Scholar] [CrossRef]
- Wolf, B.; Gressner, A.M.; Nevo, Z.; Greiling, H. Age-related decrease in the activity of UDP-xylose: Core protein xylosyltransferase in rat costal cartilage. Mech. Ageing Dev. 1982, 19, 181–190. [Google Scholar] [CrossRef]
- Wyczałkowska-Tomasik, A.; Pączek, L. Cathepsin B and L activity in the serum during the human aging process. Arch. Gerontol. Geriatr. 2012, 55, 735–738. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ly, T.-D.; Lindenkamp, C.; Kara, E.; Schmidt, V.; Kleine, A.; Fischer, B.; Hendig, D.; Knabbe, C.; Faust-Hinse, I. The Impact of Inflammatory Stimuli on Xylosyltransferase-I Regulation in Primary Human Dermal Fibroblasts. Biomedicines 2022, 10, 1451. https://doi.org/10.3390/biomedicines10061451
Ly T-D, Lindenkamp C, Kara E, Schmidt V, Kleine A, Fischer B, Hendig D, Knabbe C, Faust-Hinse I. The Impact of Inflammatory Stimuli on Xylosyltransferase-I Regulation in Primary Human Dermal Fibroblasts. Biomedicines. 2022; 10(6):1451. https://doi.org/10.3390/biomedicines10061451
Chicago/Turabian StyleLy, Thanh-Diep, Christopher Lindenkamp, Eva Kara, Vanessa Schmidt, Anika Kleine, Bastian Fischer, Doris Hendig, Cornelius Knabbe, and Isabel Faust-Hinse. 2022. "The Impact of Inflammatory Stimuli on Xylosyltransferase-I Regulation in Primary Human Dermal Fibroblasts" Biomedicines 10, no. 6: 1451. https://doi.org/10.3390/biomedicines10061451
APA StyleLy, T.-D., Lindenkamp, C., Kara, E., Schmidt, V., Kleine, A., Fischer, B., Hendig, D., Knabbe, C., & Faust-Hinse, I. (2022). The Impact of Inflammatory Stimuli on Xylosyltransferase-I Regulation in Primary Human Dermal Fibroblasts. Biomedicines, 10(6), 1451. https://doi.org/10.3390/biomedicines10061451