
Impact of Δ9-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Compounds
2.3. Synovial Fibroblast and Tissue Preparation
2.4. Intracellular Calcium and PoPo3 Uptake
2.5. Flow Cytometry
2.6. Isolation of PBMC from Peripheral Blood
2.7. RASF Co-Culture with PBMC
2.8. ELISA and Stimulation of SF
2.9. RASF Cell Viability
2.10. Statistical Analysis
3. Results
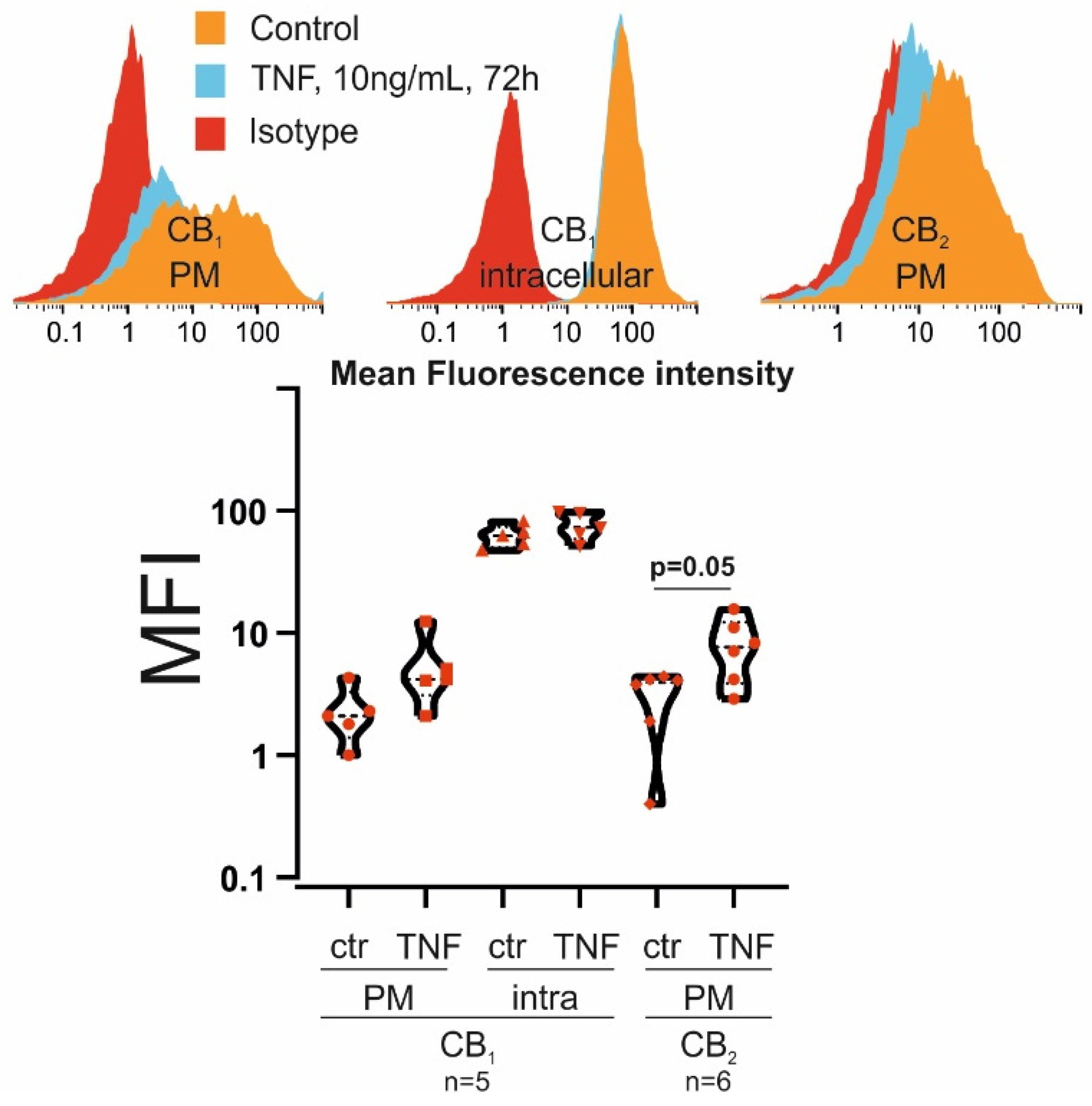
3.1. RASF Express CB1 and CB2
3.2. THC Increases Intracellular Calcium in RASF Primed with TNF
3.3. THC Enhances PoPo3 Uptake in RASF Primed with TNF
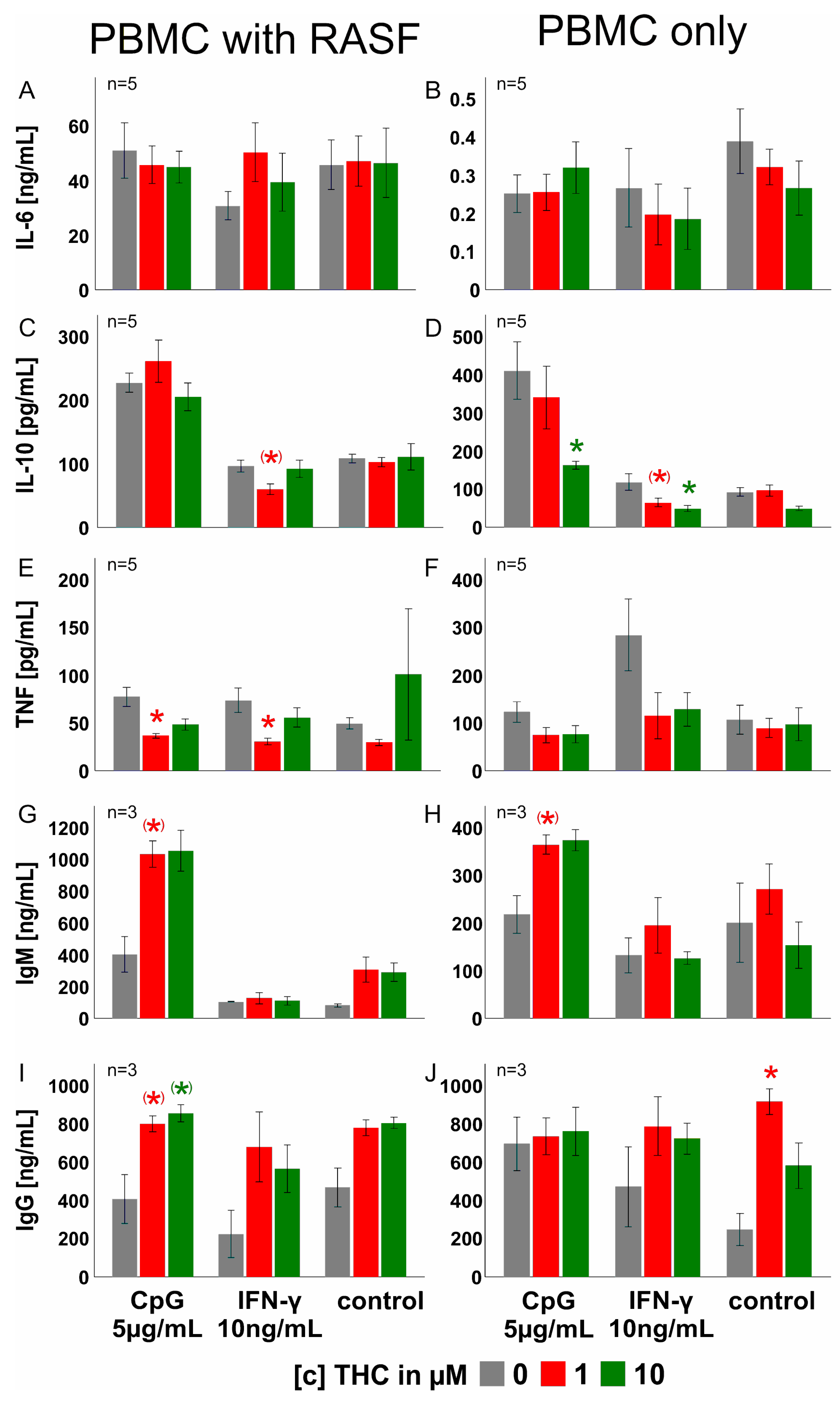
3.4. THC Reduces Cytokine Production Only at High Concentrations
3.5. THC Has Negligible Effects on PBMC Cytokine, IgM and IgG Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atakan, Z. Cannabis, a complex plant: Different compounds and different effects on individuals. Ther. Adv. Psychopharmacol. 2012, 2, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Grotenhermen, F. Pharmacology of cannabinoids. Neuro. Endocrinol. Lett. 2004, 25, 14–23. [Google Scholar] [PubMed]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2018, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, D.G. A Personal Retrospective: Elevating Anandamide (AEA) by Targeting Fatty Acid Amide Hydrolase (FAAH) and the Fatty Acid Binding Proteins (FABPs). Front. Pharmacol. 2016, 7, 370. [Google Scholar] [CrossRef] [PubMed]
- Busquets-Garcia, A.; Bains, J.; Marsicano, G. CB1 Receptor Signaling in the Brain: Extracting Specificity from Ubiquity. Neuropsychopharmacology 2018, 43, 4–20. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef]
- Kinsey, S.G.; Naidu, P.S.; Cravatt, B.F.; Dudley, D.T.; Lichtman, A.H. Fatty acid amide hydrolase blockade attenuates the development of collagen-induced arthritis and related thermal hyperalgesia in mice. Pharmacol. Biochem. Behav. 2011, 99, 718–725. [Google Scholar] [CrossRef]
- Mnich, S.J.; Hiebsch, R.R.; Huff, R.M.; Muthian, S. Anti-inflammatory properties of CB1-receptor antagonist involves beta2 adrenoceptors. J. Pharmacol. Exp. Ther. 2010, 333, 445–453. [Google Scholar] [CrossRef]
- De Laurentiis, A.; Fernandez Solari, J.; Mohn, C.; Zorrilla Zubilete, M.; Rettori, V. Endocannabinoid system participates in neuroendocrine control of homeostasis. Neuroimmunomodulation 2010, 17, 153–156. [Google Scholar] [CrossRef]
- Peters, K.Z.; Cheer, J.F.; Tonini, R. Modulating the Neuromodulators: Dopamine, Serotonin, and the Endocannabinoid System. Trends Neurosci. 2021, 44, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Watzl, B.; Scuderi, P.; Watson, R.R. Marijuana components stimulate human peripheral blood mononuclear cell secretion of interferon-gamma and suppress interleukin-1 alpha in vitro. Int. J. Immunopharmacol. 1991, 13, 1091–1097. [Google Scholar] [CrossRef]
- Deis, S.; Srivastava, R.K.; Ruiz de Azua, I.; Bindila, L.; Baraghithy, S.; Lutz, B.; Bab, I.; Tam, J. Age-related regulation of bone formation by the sympathetic cannabinoid CB1 receptor. Bone 2018, 108, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Sofia, R.D.; Nalepa, S.D.; Harakal, J.J.; Vassar, H.B. Anti-edema and analgesic properties of delta9-tetrahydrocannabinol (THC). J. Pharmacol. Exp. Ther. 1973, 186, 646–655. [Google Scholar]
- Ismail, M.; Hasan, H.; El-Orfali, Y.; Ismail, H.; Khawaja, G. Anti-Inflammatory, Antioxidative, and Hepatoprotective Effects of Trans Delta9-Tetrahydrocannabinol/Sesame Oil on Adjuvant-Induced Arthritis in Rats. Evid. Based Complement. Alternat. Med 2018, 2018, 9365464. [Google Scholar] [CrossRef]
- Cox, M.L.; Welch, S.P. The antinociceptive effect of Delta9-tetrahydrocannabinol in the arthritic rat. Eur. J. Pharmacol. 2004, 493, 65–74. [Google Scholar] [CrossRef]
- Cox, M.L.; Haller, V.L.; Welch, S.P. The antinociceptive effect of Delta9-tetrahydrocannabinol in the arthritic rat involves the CB(2) cannabinoid receptor. Eur. J. Pharmacol. 2007, 570, 50–56. [Google Scholar] [CrossRef]
- Smith, F.L.; Fujimori, K.; Lowe, J.; Welch, S.P. Characterization of delta9-tetrahydrocannabinol and anandamide antinociception in nonarthritic and arthritic rats. Pharmacol. Biochem. Behav. 1998, 60, 183–191. [Google Scholar] [CrossRef]
- Pezzato, S.; Bonetto, C.; Caimmi, C.; Tomassi, S.; Montanari, I.; Gnatta, M.G.; Fracassi, E.; Cristofalo, D.; Rossini, M.; Carletto, A.; et al. Depression is associated with increased disease activity and higher disability in a large Italian cohort of patients with rheumatoid arthritis. Adv. Rheumatol. 2021, 61, 57. [Google Scholar] [CrossRef]
- Dey, M.; Parodis, I.; Nikiphorou, E. Fatigue in Systemic Lupus Erythematosus and Rheumatoid Arthritis: A Comparison of Mechanisms, Measures and Management. J. Clin. Med. 2021, 10, 3566. [Google Scholar] [CrossRef]
- Ruscitti, P.; Cipriani, P.; Liakouli, V.; Iacono, D.; Pantano, I.; Margiotta, D.P.E.; Navarini, L.; Destro Castaniti, G.M.; Maruotti, N.; Di Scala, G.; et al. Occurrence and predictive factors of high blood pressure, type 2 diabetes, and metabolic syndrome in rheumatoid arthritis: Findings from a 3-year, multicentre, prospective, observational study. Clin. Exp. Rheumatol. 2021, 39, 995–1002. [Google Scholar] [PubMed]
- Kemble, S.; Croft, A.P. Critical Role of Synovial Tissue-Resident Macrophage and Fibroblast Subsets in the Persistence of Joint Inflammation. Front. Immunol. 2021, 12, 715894. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Straub, R.H. Cannabinoid-based drugs targeting CB1 and TRPV1, the sympathetic nervous system, and arthritis. Arthritis Res. Ther. 2015, 17, 226. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Schneider, M.; Pongratz, G. Joints for joints: Cannabinoids in the treatment of rheumatoid arthritis. Curr. Opin. Rheumatol. 2019, 31, 271–278. [Google Scholar] [CrossRef]
- Straub, R.H. Interaction of the endocrine system with inflammation: A function of energy and volume regulation. Arthritis Res. Ther. 2014, 16, 203. [Google Scholar] [CrossRef]
- Pongratz, G.; Straub, R.H. The sympathetic nervous response in inflammation. Arthritis Res. Ther. 2014, 16, 504. [Google Scholar] [CrossRef]
- Nygaard, G.; Firestein, G.S. Restoring synovial homeostasis in rheumatoid arthritis by targeting fibroblast-like synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the cannabinoid receptor system in synovial tissue and fluid in patients with osteoarthritis and rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef]
- Lowin, T.; Zhu, W.; Dettmer-Wilde, K.; Straub, R.H. Cortisol-mediated adhesion of synovial fibroblasts is dependent on the degradation of anandamide and activation of the endocannabinoid system. Arthritis Rheum. 2012, 64, 3867–3876. [Google Scholar] [CrossRef]
- Gui, H.; Liu, X.; Wang, Z.W.; He, D.Y.; Su, D.F.; Dai, S.M. Expression of cannabinoid receptor 2 and its inhibitory effects on synovial fibroblasts in rheumatoid arthritis. Rheumatology 2014, 53, 802–809. [Google Scholar] [CrossRef]
- Lowin, T.; Pongratz, G.; Straub, R.H. The synthetic cannabinoid WIN55,212-2 mesylate decreases the production of inflammatory mediators in rheumatoid arthritis synovial fibroblasts by activating CB2, TRPV1, TRPA1 and yet unidentified receptor targets. J. Inflamm. 2016, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Fechtner, S.; Singh, A.K.; Ahmed, S. Role of cannabinoid receptor 2 in mediating interleukin-1beta-induced inflammation in rheumatoid arthritis synovial fibroblasts. Clin. Exp. Rheumatol. 2019, 37, 1026–1035. [Google Scholar] [PubMed]
- Selvi, E.; Lorenzini, S.; Garcia-Gonzalez, E.; Maggio, R.; Lazzerini, P.E.; Capecchi, P.L.; Balistreri, E.; Spreafico, A.; Niccolini, S.; Pompella, G.; et al. Inhibitory effect of synthetic cannabinoids on cytokine production in rheumatoid fibroblast-like synoviocytes. Clin. Exp. Rheumatol. 2008, 26, 574–581. [Google Scholar] [PubMed]
- Springs, A.E.; Karmaus, P.W.; Crawford, R.B.; Kaplan, B.L.; Kaminski, N.E. Effects of targeted deletion of cannabinoid receptors CB1 and CB2 on immune competence and sensitivity to immune modulation by Delta9-tetrahydrocannabinol. J. Leukoc. Biol. 2008, 84, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, F.; Burger, F.; Mach, F.; Steffens, S. CB2 cannabinoid receptor agonist JWH-015 modulates human monocyte migration through defined intracellular signaling pathways. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1145–H1155. [Google Scholar] [CrossRef]
- Lunn, C.A.; Fine, J.; Rojas-Triana, A.; Jackson, J.V.; Lavey, B.; Kozlowski, J.A.; Hipkin, R.W.; Lundell, D.J.; Bober, L. Cannabinoid CB(2)-selective inverse agonist protects against antigen-induced bone loss. Immunopharmacol. Immunotoxicol. 2007, 29, 387–401. [Google Scholar] [CrossRef]
- McDougall, J.J.; Yu, V.; Thomson, J. In vivo effects of CB2 receptor-selective cannabinoids on the vasculature of normal and arthritic rat knee joints. Br. J. Pharmacol. 2008, 153, 358–366. [Google Scholar] [CrossRef]
- Zhu, M.; Yu, B.; Bai, J.; Wang, X.; Guo, X.; Liu, Y.; Lin, J.; Hu, S.; Zhang, W.; Tao, Y.; et al. Cannabinoid Receptor 2 Agonist Prevents Local and Systemic Inflammatory Bone Destruction in Rheumatoid Arthritis. J. Bone Miner. Res. 2019, 34, 739–751. [Google Scholar] [CrossRef]
- Bai, J.; Ge, G.; Wang, Y.; Zhang, W.; Wang, Q.; Wang, W.; Guo, X.; Yu, B.; Xu, Y.; Yang, H.; et al. A selective CB2 agonist protects against the inflammatory response and joint destruction in collagen-induced arthritis mice. Biomed. Pharmacother. 2019, 116, 109025. [Google Scholar] [CrossRef]
- Arnett, F.C.; Edworthy, S.M.; Bloch, D.A.; McShane, D.J.; Fries, J.F.; Cooper, N.S.; Healey, L.A.; Kaplan, S.R.; Liang, M.H.; Luthra, H.S.; et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988, 31, 315–324. [Google Scholar] [CrossRef]
- Miller, L.E.; Justen, H.P.; Scholmerich, J.; Straub, R.H. The loss of sympathetic nerve fibers in the synovial tissue of patients with rheumatoid arthritis is accompanied by increased norepinephrine release from synovial macrophages. FASEB J. 2000, 14, 2097–2107. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.; Chen, S.; Shaw, A.C.; Allore, H.G. Statistical approaches for analyzing immunologic data of repeated observations: A practical guide. J. Immunol. Methods 2013, 398–399, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J. Statistics for Immunologists. Curr. Protoc. Immunol. 2018, 122, 54. [Google Scholar] [CrossRef] [PubMed]
- Lew, M. Good statistical practice in pharmacology. Problem 2. Br. J. Pharmacol. 2007, 152, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Apitz, M.; Anders, S.; Straub, R.H. Anti-inflammatory effects of N-acylethanolamines in rheumatoid arthritis synovial cells are mediated by TRPV1 and TRPA1 in a COX-2 dependent manner. Arthritis Res. Ther. 2015, 17, 321. [Google Scholar] [CrossRef]
- Kim, D.J.; Thayer, S.A. Activation of CB1 cannabinoid receptors inhibits neurotransmitter release from identified synaptic sites in rat hippocampal cultures. Brain Res. 2000, 852, 398–405. [Google Scholar] [CrossRef]
- Lowin, T.; Tingting, R.; Zurmahr, J.; Classen, T.; Schneider, M.; Pongratz, G. Cannabidiol (CBD): A killer for inflammatory rheumatoid arthritis synovial fibroblasts. Cell Death Dis. 2020, 11, 714. [Google Scholar] [CrossRef]
- Viele, D.; Betz, E. Effect of the calcium entry blocker, flunarizine, on ruthenium red uptake by endothelial cells following acute electrical stimulation of rabbit carotid arteries. Basic Res. Cardiol. 1985, 80, 59–65. [Google Scholar] [CrossRef]
- Sido, J.M.; Nagarkatti, P.S.; Nagarkatti, M. Production of endocannabinoids by activated T cells and B cells modulates inflammation associated with delayed-type hypersensitivity. Eur. J. Immunol. 2016, 46, 1472–1479. [Google Scholar] [CrossRef]
- Goncalves-Monteiro, S.; Ribeiro-Oliveira, R.; Vieira-Rocha, M.S.; Vojtek, M.; Sousa, J.B.; Diniz, C. Insights into Nuclear G-Protein-Coupled Receptors as Therapeutic Targets in Non-Communicable Diseases. Pharmaceuticals 2021, 14, 429. [Google Scholar] [CrossRef]
- DeNies, M.S.; Smrcka, A.V.; Schnell, S.; Liu, A.P. beta-arrestin mediates communication between plasma membrane and intracellular GPCRs to regulate signaling. Commun. Biol. 2020, 3, 789. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.J.; Yin, N.; Gao, F.; Miao, Y.; Li, Q.; Li, F.; Sun, X.H.; Yang, X.L.; Wang, Z. Cannabinoid CB1 and CB2 receptors differentially modulate L- and T-type Ca2+ channels in rat retinal ganglion cells. Neuropharmacology 2017, 124, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Laguerre, A.; Keutler, K.; Hauke, S.; Schultz, C. Regulation of Calcium Oscillations in beta-Cells by Co-activated Cannabinoid Receptors. Cell Chem. Biol. 2021, 28, 88–96.e3. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Bleck, J.; Schneider, M.; Pongratz, G. Selective killing of proinflammatory synovial fibroblasts via activation of transient receptor potential ankyrin (TRPA1). Biochem. Pharmacol. 2018, 154, 293–302. [Google Scholar] [CrossRef]
- Neumann-Raizel, H.; Shilo, A.; Lev, S.; Mogilevsky, M.; Katz, B.; Shneor, D.; Shaul, Y.D.; Leffler, A.; Gabizon, A.; Karni, R.; et al. 2-APB and CBD-Mediated Targeting of Charged Cytotoxic Compounds Into Tumor Cells Suggests the Involvement of TRPV2 Channels. Front. Pharmacol. 2019, 10, 1198. [Google Scholar] [CrossRef]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef]
- Storozhuk, M.V.; Zholos, A.V. TRP Channels as Novel Targets for Endogenous Ligands: Focus on Endocannabinoids and Nociceptive Signalling. Curr. Neuropharmacol. 2018, 16, 137–150. [Google Scholar] [CrossRef]
- Dall’Acqua, M.C.; Bonet, I.J.; Zampronio, A.R.; Tambeli, C.H.; Parada, C.A.; Fischer, L. The contribution of transient receptor potential ankyrin 1 (TRPA1) to the in vivo nociceptive effects of prostaglandin E(2). Life Sci. 2014, 105, 7–13. [Google Scholar] [CrossRef]
- Gamage, T.F.; Farquhar, C.E.; Lefever, T.W.; Marusich, J.A.; Kevin, R.C.; McGregor, I.S.; Wiley, J.L.; Thomas, B.F. Molecular and Behavioral Pharmacological Characterization of Abused Synthetic Cannabinoids MMB- and MDMB-FUBINACA, MN-18, NNEI, CUMYL-PICA, and 5-Fluoro-CUMYL-PICA. J. Pharmacol. Exp. Ther. 2018, 365, 437–446. [Google Scholar] [CrossRef]
- Bottini, N.; Firestein, G.S. Duality of fibroblast-like synoviocytes in RA: Passive responders and imprinted aggressors. Nat. Rev. Rheumatol. 2013, 9, 24–33. [Google Scholar] [CrossRef]
- Storch, H.; Zimmermann, B.; Resch, B.; Tykocinski, L.O.; Moradi, B.; Horn, P.; Kaya, Z.; Blank, N.; Rehart, S.; Thomsen, M.; et al. Activated human B cells induce inflammatory fibroblasts with cartilage-destructive properties and become functionally suppressed in return. Ann. Rheum. Dis. 2016, 75, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.H.; Graham, J.; Lemberger, L. Marihuana, tetrahydrocannabinol and T-cell function. Life Sci. 1976, 19, 395–400. [Google Scholar] [CrossRef]
- Rachelefsky, G.S.; Opelz, G. Normal lymphocyte function in the presence of delta-9-tetrahydrocannabinol. Clin. Pharmacol. Ther. 1977, 21, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Karmaus, P.W.; Chen, W.; Kaplan, B.L.; Kaminski, N.E. Delta9-tetrahydrocannabinol suppresses cytotoxic T lymphocyte function independent of CB1 and CB 2, disrupting early activation events. J. Neuroimmune Pharmacol. 2012, 7, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.D.; Castaneda, J.T.; Kiertscher, S.M. Exposure to Delta9-Tetrahydrocannabinol Impairs the Differentiation of Human Monocyte-derived Dendritic Cells and their Capacity for T cell Activation. J. Neuroimmune Pharmacol. 2015, 10, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.S.; Majhi, R.K.; Tiwari, A.; Acharya, T.; Kumar, P.S.; Saha, S.; Kumar, A.; Goswami, C.; Chattopadhyay, S. Transient receptor potential ankyrin1 channel is endogenously expressed in T cells and is involved in immune functions. Biosci. Rep. 2019, 39, BSR20191437. [Google Scholar] [CrossRef]
- Gardner, B.; Zu, L.X.; Sharma, S.; Liu, Q.; Makriyannis, A.; Tashkin, D.P.; Dubinett, S.M. Autocrine and paracrine regulation of lymphocyte CB2 receptor expression by TGF-beta. Biochem. Biophys. Res. Commun. 2002, 290, 91–96. [Google Scholar] [CrossRef]
- Eisenstein, T.K.; Meissler, J.J.; Wilson, Q.; Gaughan, J.P.; Adler, M.W. Anandamide and Delta9-tetrahydrocannabinol directly inhibit cells of the immune system via CB2 receptors. J. Neuroimmunol. 2007, 189, 17–22. [Google Scholar] [CrossRef]
- Buckley, N.E.; McCoy, K.L.; Mezey, E.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar] [CrossRef]
- Basu, S.; Ray, A.; Dittel, B.N. Cannabinoid Receptor 2 (CB2) Plays a Role in the Generation of Germinal Center and Memory B Cells, but Not in the Production of Antigen-Specific IgG and IgM, in Response to T-dependent Antigens. PLoS ONE 2013, 8, e67587. [Google Scholar] [CrossRef]
- Newton, C.A.; Klein, T.W. Cannabinoid 2 (CB2) receptor involvement in the down-regulation but not up-regulation of serum IgE levels in immunized mice. J. Neuroimmune Pharmacol. 2012, 7, 591–598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Agudelo, M.; Newton, C.; Widen, R.; Sherwood, T.; Nong, L.; Friedman, H.; Klein, T.W. Cannabinoid receptor 2 (CB2) mediates immunoglobulin class switching from IgM to IgE in cultures of murine-purified B lymphocytes. J. Neuroimmune Pharmacol. 2008, 3, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sido, J.M.; Jackson, A.R.; Nagarkatti, P.S.; Nagarkatti, M. Marijuana-derived Delta-9-tetrahydrocannabinol suppresses Th1/Th17 cell-mediated delayed-type hypersensitivity through microRNA regulation. J. Mol. Med. 2016, 94, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Ngaotepprutaram, T.; Kaplan, B.L.; Carney, S.; Crawford, R.; Kaminski, N.E. Suppression by Delta(9)-tetrahydrocannabinol of the primary immunoglobulin M response by human peripheral blood B cells is associated with impaired STAT3 activation. Toxicology 2013, 310, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ranganathan, M.; Shu, C.; Liang, X.; Ganesh, S.; Osafo-Addo, A.; Yan, C.; Zhang, X.; Aouizerat, B.E.; Krystal, J.H.; et al. Single-cell Transcriptome Mapping Identifies Common and Cell-type Specific Genes Affected by Acute Delta9-tetrahydrocannabinol in Humans. Sci. Rep. 2020, 10, 3450. [Google Scholar] [CrossRef]
- Ghosh, S.; Kinsey, S.G.; Liu, Q.S.; Hruba, L.; McMahon, L.R.; Grim, T.W.; Merritt, C.R.; Wise, L.E.; Abdullah, R.A.; Selley, D.E.; et al. Full Fatty Acid Amide Hydrolase Inhibition Combined with Partial Monoacylglycerol Lipase Inhibition: Augmented and Sustained Antinociceptive Effects with Reduced Cannabimimetic Side Effects in Mice. J. Pharmacol. Exp. Ther. 2015, 354, 111–120. [Google Scholar] [CrossRef]
- Habib, G.; Khazin, F.; Artul, S. The Effect of Medical Cannabis on Pain Level and Quality of Sleep among Rheumatology Clinic Outpatients. Pain. Res. Manag. 2021, 2021, 1756588. [Google Scholar] [CrossRef]
- Graczyk, M.; Lukowicz, M.; Dzierzanowski, T. Prospects for the Use of Cannabinoids in Psychiatric Disorders. Front. Psychiatry 2021, 12, 620073. [Google Scholar] [CrossRef]
- Mendiguren, A.; Aostri, E.; Pineda, J. Regulation of noradrenergic and serotonergic systems by cannabinoids: Relevance to cannabinoid-induced effects. Life Sci. 2018, 192, 115–127. [Google Scholar] [CrossRef]
- Luo, Y.; Sun, W.; Feng, X.; Ba, X.; Liu, T.; Guo, J.; Xiao, L.; Jiang, J.; Hao, Y.; Xiong, D.; et al. (-)-menthol increases excitatory transmission by activating both TRPM8 and TRPA1 channels in mouse spinal lamina II layer. Biochem. Biophys. Res. Commun. 2019, 516, 825–830. [Google Scholar] [CrossRef]
- Capellino, S.; Weber, K.; Gelder, M.; Harle, P.; Straub, R.H. First appearance and location of catecholaminergic cells during experimental arthritis and elimination by chemical sympathectomy. Arthritis Rheum. 2012, 64, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Jenei-Lanzl, Z.; Capellino, S.; Kees, F.; Fleck, M.; Lowin, T.; Straub, R.H. Anti-inflammatory effects of cell-based therapy with tyrosine hydroxylase-positive catecholaminergic cells in experimental arthritis. Ann Rheum. Dis. 2015, 74, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, J.; Wang, C.; Liu, L.; Wu, Y.; Zhang, Y.; Zhou, A.; Zhang, L.; Wei, W. beta2-adrenoceptor signaling reduction is involved in the inflammatory response of fibroblast-like synoviocytes from adjuvant-induced arthritic rats. Inflammopharmacology 2019, 27, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Makda, A.A.; Elmore, M.A.; Hill, M.E.; Stamps, A.; Tejura, S.; Finnen, M.J. Differential effects of CB1 and CB2 agonists on cAMP levels and MAP kinase activation in human peripheral blood mononuclear cells. Biochem. Soc. Trans. 1997, 25, 217S. [Google Scholar] [CrossRef]
- Kraemer, M.; Madea, B.; Hess, C. Detectability of various cannabinoids in plasma samples of cannabis users: Indicators of recent cannabis use? Drug Test. Anal. 2019, 11, 1498–1506. [Google Scholar] [CrossRef]
- Maia, J.; Midao, L.; Cunha, S.C.; Almada, M.; Fonseca, B.M.; Braga, J.; Goncalves, D.; Teixeira, N.; Correia-da-Silva, G. Effects of cannabis tetrahydrocannabinol on endocannabinoid homeostasis in human placenta. Arch. Toxicol. 2019, 93, 649–658. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lowin, T.; Kok, C.; Smutny, S.; Pongratz, G. Impact of Δ9-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells. Biomedicines 2022, 10, 1118. https://doi.org/10.3390/biomedicines10051118
Lowin T, Kok C, Smutny S, Pongratz G. Impact of Δ9-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells. Biomedicines. 2022; 10(5):1118. https://doi.org/10.3390/biomedicines10051118
Chicago/Turabian StyleLowin, Torsten, Christina Kok, Sophie Smutny, and Georg Pongratz. 2022. "Impact of Δ9-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells" Biomedicines 10, no. 5: 1118. https://doi.org/10.3390/biomedicines10051118
APA StyleLowin, T., Kok, C., Smutny, S., & Pongratz, G. (2022). Impact of Δ9-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells. Biomedicines, 10(5), 1118. https://doi.org/10.3390/biomedicines10051118