
Activation of Calcium-Activated Chloride Channels Suppresses Inherited Seizure Susceptibility in Genetically Epilepsy-Prone Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Acoustically Evoked Seizure Testing
2.3. Pharmacological Treatments
2.4. Data Analysis
3. Results
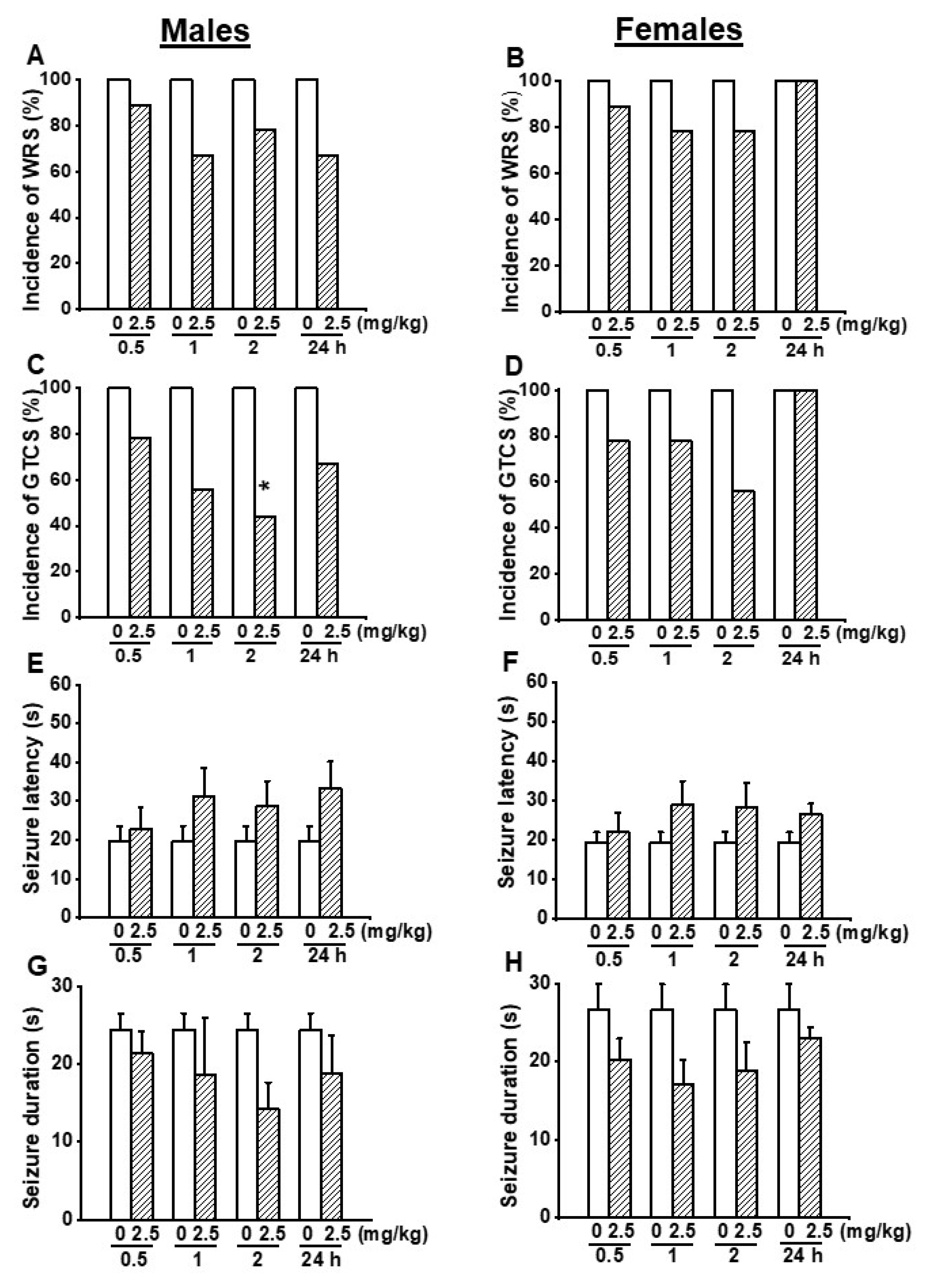
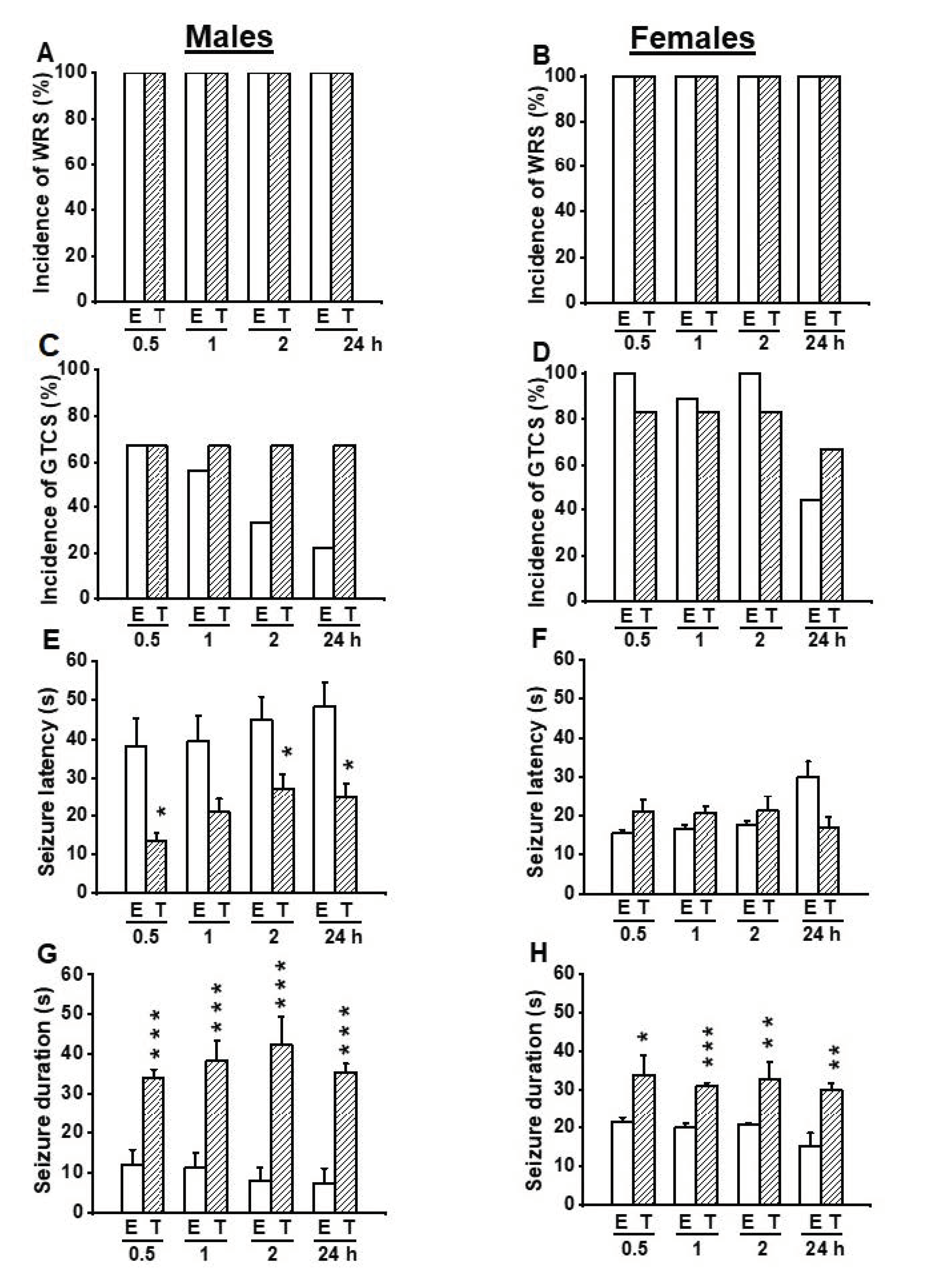
3.1. Effects on EACT at the Dose of 2.5 mg/kg on Acoustically Evoked Seizures in GEPR-3s
3.2. Effects on EACT at the Dose of 5 mg/kg on Acoustically Evoked Seizures in GEPR-3s
3.3. Effects on EACT at the Dose of 10 mg/kg on Acoustically Evoked Seizures in GEPR-3s
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Devinsky, O.; Bundock, E.; Hesdorffer, D.; Donner, E.; Moseley, B.; Cihan, E.; Hussain, F.; Friedman, D. Resolving ambiguities in SUDEP classification. Epilepsia 2018, 59, 1220–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, M.; Oliva, A.; Grassi, S.; Brigada, R.; Campuzano, O. Update on the genetic basis of sudden unexpected death in Epilepsy. Int. J. Mol. Sci. 2019, 20, 1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zack, M.M.; Kobau, R. National and State Estimates of the Numbers of Adults and Children with Active Epilepsy—United States, 2015. Morb. Mortal. Wkly. Rep. 2017, 66, 821–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. World Health Organization: Epilepsy: Epidemiology, Aetiology, and Prognosis; WHO Factsheet; WHO: Geneva, Switzerland, 2001.
- French, J.A. Refractory epilepsy: Clinical overview. Epilepsia 2007, 48 (Suppl. S1), 3–7. [Google Scholar] [CrossRef]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Allen Hauser, W.; Mathern, G.; Moshé, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1107. [Google Scholar] [CrossRef]
- Lux, H.D.; Heinemann, U. Ionic changes during experimentally induced seizure activity. Electrencephalogr. Clin. Neurophysiol. 1978, 34, 289–297. [Google Scholar]
- Heinemann, U.; Konnerth, A.; Pumain, R.; Wadman, W.J. Extracellular calcium and potassium concentration changes in chronic epileptic brain tissue. Adv. Neurol. 1986, 44, 641–661. [Google Scholar]
- Albowitz, B.; König, P.; Kuhnt, U. Spatiotemporal distribution of intracellular calcium transients during epileptiform activity in guinea pig hippocampal slices. J. Neurophysiol. 1997, 77, 491–501. [Google Scholar] [CrossRef]
- Delorenzo, R.J.; Sun, D.A.; Deshpande, L.S. Cellular mechanisms underlying acquired epilepsy: The calcium hypothesis of the induction and maintenance of epilepsy. Pharmacol. Ther. 2005, 105, 229–266. [Google Scholar] [CrossRef] [Green Version]
- De Sarro, G.; De Sarro, A.; Federico, F.; Meldrum, B.S. Anticonvulsant properties of some calcium antagonists on sound-induced seizures in genetically epilepsy prone rats. Gen. Pharmacol. 1990, 21, 768–778. [Google Scholar] [CrossRef]
- De Sarro, G.; Ascioti, C.; Di Paola, E.D.; Vidal, M.J.; De Sarro, A. Effects of antiepileptic drugs, calcium channel blockers and other compounds on seizures induced by activation of voltage-dependent L calcium channel in DBA/2 mice. Gen. Pharmacol. 1992, 23, 1205–1216. [Google Scholar] [CrossRef]
- De Sarro, G.; Russo, E.; Citraro, R.; Meldrum, B.S. Genetically epilepsy-prone rats (GEPRs) and DBA/2 mice: Two animal models of audiogenic reflex epilepsy for the evaluation of new generation AEDs. Epilepsy Behav. 2017, 71, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Khandai, P.; Forcelli, P.A.; N’Gouemo, P. Activation of small conductance calcium-activated potassium channels suppresses seizure susceptibility in the genetically epilepsy-prone rats. Neuropharmacology 2020, 163, 107865. [Google Scholar] [CrossRef] [PubMed]
- Faingold, C.L. Neuronal networks in the genetically epilepsy-prone rat. Adv. Neurol. 1999, 79, 311–321. [Google Scholar]
- N’Gouemo, P.; Faingold, C.L.; Morad, M. Calcium channel dysfunction in inferior colliculus neurons of the genetically epilepsy-prone rat. Neuropharmacology 2009, 56, 665–675. [Google Scholar] [CrossRef] [Green Version]
- N’Gouemo, P.; Yasuda, R.P.; Faingold, C. Seizure susceptibility is associated with altered protein expression of voltage-gated calcium channel subunits in inferior colliculus neurons of the genetically epilepsy-prone rats. Brain Res. 2010, 1308, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, J.V.; Burman, R.J.; Katz, A.A.; Akerman, C.J. Ion dynamics during seizures. Front. Cell. Neurosci. 2015, 9, 419. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, J.V.; Joyce, B.; Kay, L.; Schlagheck, T.; Newey, S.E.; Srinivas, S.; Akerman, C.J. A genetically encoded chloride and pH sensor for dissociating ion dynamics in the nervous system. Front. Cell. Neurosci. 2013, 7, 202. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M. A calcium-activated chloride current generates the after-hyperpolarization of rat sensory neurons in culture. J. Physiol. Lond. 1985, 364, 217–239. [Google Scholar] [CrossRef]
- Hartzell, C.; Putzier, I.; Arreola, J. Calcium-activated chloride channels. Annu. Rev. Physiol. 2005, 67, 719–758. [Google Scholar] [CrossRef] [Green Version]
- Shroeder, B.C.; Cheng, T.; Jan, Y.N.; Jan, L.Y. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 2008, 134, 1019–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputo, A.; Caci, E.; Ferrera, L.; Pedemonte, N.; Sondo, E.; Pfeffer, U.; Ravazzolo, R.; Zegarra-Moran, O.; Galietta, L.J. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008, 322, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.M.; et al. TMEM16A confers receptor-activated calcium dependent chloride conductance. Nature 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Kunzelmann, K.; Kongsuphol, P.; Aldehni, F.; Tian, Y.; Ousingsawat, J.; Warth, R.; Schreiber, R. Bestrophin and TMEM16-Ca2+ activated Cl- channels with different functions. Cell Calcium 2009, 46, 233–241. [Google Scholar] [CrossRef]
- Kunzelmann, K.; Kongsuphol, P.; Chootip, K.; Toledo, C.; Martins, J.R.; Almaca, J.; Tian, Y.; Witzgall, R.; Ousingsawat, J.; Schreiber, R. Role of Ca2+-activated Cl− channels bestrophin and anoctamin in epithelial cells. Biol. Chem. 2009, 392, 125–134. [Google Scholar] [CrossRef]
- Cho, S.J.; Jeon, J.H.; Chun, D.I.; Yeo, S.W.; Kim, I.-B. Anoctamin 1 expression in the mouse auditory brainstem. Cell Tissue Res. 2014, 357, 563–569. [Google Scholar] [CrossRef]
- National Research Council (U.S.); Institute for Laboratory Animal Research (U.S.); National Academies Press (U.S.). Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Mishra, P.K.; Dailey, J.W.; Reigel, C.E.; Jobe, P.C. Audiogenic convulsions in moderate seizure genetically epilepsy-prone rats (GEPR-3s). Epilepsy Res. 1989, 3, 191–198. [Google Scholar] [CrossRef]
- Wang, T.A.; Chen, C.; Huang, F.; Feng, S.; Tien, J.; Braz, J.M.; Basbaum, A.I.; Jan, Y.N.; Jan, L.Y. TMEM16C is involved in thermoregulation and protects rodent pups from febrile seizures. Proc. Natl. Acad. Sci. USA 2021, 118, e2023342118. [Google Scholar] [CrossRef]
- Ha, G.E.; Cheon, G. Calcium-activated chloride channels: New target to control the spiking pattern of neurons. BMR Rep. 2017, 50, 109–110. [Google Scholar] [CrossRef] [Green Version]
- Verma-Ahuja, S.; Pencek, T.L. Hippocampal CA1 neurons properties in the genetically epilepsy-prone rats. Epilepsy Res. 1994, 18, 205–215. [Google Scholar] [CrossRef]
- Verma-Ahuja, S.; Evans, M.S.; Pencek, T.L. Evidence for decreased calcium dependent potassium conductance in hippocampal CA3 neurons of the genetically epilepsy-prone rats. Epilepsy Res. 1995, 22, 137–144. [Google Scholar] [CrossRef]
- N’Gouemo, P.; Yasuda, R.P.; Faingold, C.L. Protein expression of small conductance calcium-activated potassium channels is altered in inferior colliculus neurons of the genetically epilepsy-prone rat. Brain Res. 2009, 1270, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Xiao, S.; Huang, F.; Harfe, B.D.; Jan, Y.N.; Jan, L.Y. Calcium-activated chloride channels (CaCCs) regulate action potential and synaptic response in hippocampal neurons. Neuron 2012, 74, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, W.; Schmelzeisen, S.; Parthier, D.; Möhrlen, F. Anoctamin and calcium-activated chloride channels may modulate inhibitory transmission in the cerebellar cortex. PLoS ONE 2015, 10, e0142160. [Google Scholar]
- Ha, G.E.; Lee, J.; Kwak, H.; Song, K.; Kwon, J.; Jung, S.-Y.; Hong, J.; Chang, G.-E.; Hwang, E.M.; Shin, H.-S.; et al. The Ca2+-activated chloride channel anoctamin-2 mediates spike-frequency adaptation and regulates sensory transmission in thalamocortical neurons. Nat. Commun. 2016, 7, 13791–13803. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Simms, J.; Peters, C.J.; Tynan-La Fontaine, M.; Li, K.; Gill, M.; Jan, Y.N.; Jan, L.Y. TMEM16B calcium activated chloride channels regulate action potential firing in lateral septum and aggression in male mice. J. Neurosci. 2019, 39, 7102–7117. [Google Scholar] [CrossRef]
- Alfonsa, H.; Merricks, E.M.; Codadu, N.K.; Cunningham, M.O.; Deisseroth, K.; Racca, C.; Trevelyan, A.J. The contribution of raised intraneuronal chloride to epileptic network activity. J. Neurosci. 2015, 35, 7715–7726. [Google Scholar] [CrossRef]
- Liu, S.; Feng, J.; Luo, J.; Yang, P.; Brett, T.J.; Hu, H. Eact, a small molecule activator of TMEM16A, activates TRPV1 and elicits pain- and itch-related behaviours. Br. J. Pharmacol. 2016, 173, 1208–1218. [Google Scholar] [CrossRef] [Green Version]
- Akita, T.; Fukuda, A. Intracellular Cl− dysregulation causing and caused by pathological neuronal activity. Pflügers Arch.-Eur. J. Physiol. 2020, 472, 977–987. [Google Scholar] [CrossRef]
- Cho, S.J.; Vaca, M.A.; Miranda, C.J.; N’Gouemo, P. Inhibition of transient potential receptor vanilloid type 1 suppresses seizure susceptibility in the genetically epilepsy-prone rat. CNS Neurosci. Ther. 2018, 24, 18–28. [Google Scholar] [CrossRef]
- Naritoku, D.K.; Mecozzi, L.B.; Aiello, M.T.; Faingold, C.L. Repetition of audiogenic seizures in genetically epilepsy-prone rats induces cortical epileptiform activity and additional seizure behaviors. Exp. Neurol. 1992, 115, 317–324. [Google Scholar] [CrossRef]
- Martinez, L.A.; Lai, Y.-C.; Holder, J.L.; Anderson, A.E. Genetics in epilepsy. Neurol. Clin. 2021, 39, 743–777. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, M.; Simms, M.; N’Gouemo, P. Activation of Calcium-Activated Chloride Channels Suppresses Inherited Seizure Susceptibility in Genetically Epilepsy-Prone Rats. Biomedicines 2022, 10, 449. https://doi.org/10.3390/biomedicines10020449
Thomas M, Simms M, N’Gouemo P. Activation of Calcium-Activated Chloride Channels Suppresses Inherited Seizure Susceptibility in Genetically Epilepsy-Prone Rats. Biomedicines. 2022; 10(2):449. https://doi.org/10.3390/biomedicines10020449
Chicago/Turabian StyleThomas, Miracle, Mark Simms, and Prosper N’Gouemo. 2022. "Activation of Calcium-Activated Chloride Channels Suppresses Inherited Seizure Susceptibility in Genetically Epilepsy-Prone Rats" Biomedicines 10, no. 2: 449. https://doi.org/10.3390/biomedicines10020449
APA StyleThomas, M., Simms, M., & N’Gouemo, P. (2022). Activation of Calcium-Activated Chloride Channels Suppresses Inherited Seizure Susceptibility in Genetically Epilepsy-Prone Rats. Biomedicines, 10(2), 449. https://doi.org/10.3390/biomedicines10020449