

Immunohistochemical Detection of Estrogen Receptor-Beta (ERβ) with PPZ0506 Antibody in Murine Tissue: From Pitfalls to Optimization




Abstract
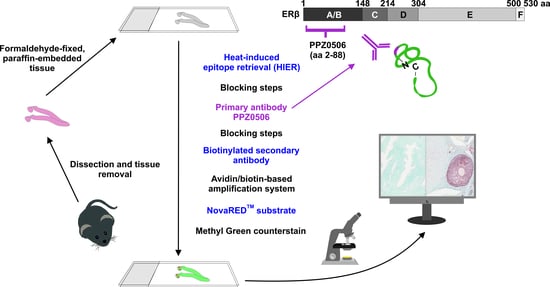
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Hematoxylin-Eosin Staining
2.4. Immunohistochemistry Staining Protocol
2.5. RNA Extraction and RT-PCR
2.6. Protein Extraction and Western Blot Analysis
2.7. Aligment of ERβ Protein Sequence in Different Species
3. Results
3.1. Detection of ERβ in Murine Tissues
3.2. Antigen Retrieval Is Mandatory for Nuclear ERβ Detection
3.3. Immunhistochemical Detection of Nucelar ERβ
3.4. Titration of PPZ0506 Primary Antibody
3.5. Non-Specific Background Staining
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Actb | gene encoding beta actin |
| BLAST | basic local alignment search tool |
| BSA | bovine serum albumin |
| CRISPR | clustered regulatory interspaced short palindromic repeats |
| Esr | gene encoding estrogen receptor |
| ERα | Estrogen receptor alpha |
| ERβ | Estrogen receptor beta |
| FBS | fetal bovine serum |
| HIER | Heat-induced epitope retrieval |
| Igs | Immunoglobulins |
| PPZ0506 | Estrogen beta monoclonal antibody, clone #PPZ0506 |
| Rn18s | gene encoding 18S ribosomal RNA |
| RT-PCR | reverse transcription polymerase chain reaction |
References
- Greene, G.L.; Gilna, P.; Waterfield, M.; Baker, A.; Hort, Y.; Shine, J. Sequence and expression of human estrogen receptor complementary DNA. Science 1986, 231, 1150–1154. [Google Scholar] [CrossRef]
- White, R.; Lees, J.A.; Needham, M.; Ham, J.; Parker, M. Structural organization and expression of the mouse estrogen receptor. Mol. Endocrinol. 1987, 1, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Sakai, M.; Muramatsu, M. Molecular cloning and characterization of rat estrogen receptor cDNA. Nucleic Acids Res. 1987, 15, 2499–2513. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, M.; Inoue, S. Estrogen receptors: How do they control reproductive and nonreproductive functions? Biochem. Biophys. Res. Commun. 2000, 270, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mahboobifard, F.; Pourgholami, M.H.; Jorjani, M.; Dargahi, L.; Amiri, M.; Sadeghi, S.; Tehrani, F.R. Estrogen as a key regulator of energy homeostasis and metabolic health. Biomed. Pharmacother. 2022, 156, 113808. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef]
- Couse, J.F.; Lindzey, J.; Grandien, K.; Gustafsson, J.A.; Korach, K.S. Tissue distribution and quantitative analysis of estrogen receptor-alpha (ERalpha) and estrogen receptor-beta (ERbeta) messenger ribonucleic acid in the wild-type and ERalpha-knockout mouse. Endocrinology 1997, 138, 4613–4621. [Google Scholar] [CrossRef]
- Hiroi, H.; Inoue, S.; Watanabe, T.; Goto, W.; Orimo, A.; Momoeda, M.; Tsutsumi, O.; Taketani, Y.; Muramatsu, M. Differential immunolocalization of estrogen receptor α and in rat ovary and uterus. J. Mol. Endocrinol. 1999, 22, 37–44. [Google Scholar] [CrossRef]
- Sar, M.; Welsch, F. Differential expression of estrogen receptor-beta and estrogen receptor-alpha in the rat ovary. Endocrinology 1999, 140, 963–971. [Google Scholar] [CrossRef]
- Hishikawa, Y.; Damavandi, E.; Izumi, S.; Koji, T. Molecular histochemical analysis of estrogen receptor alpha and beta expressions in the mouse ovary: In situ hybridization and Southwestern histochemistry. Med. Electron. Microsc. 2003, 36, 67–73. [Google Scholar] [CrossRef]
- Gonzalez, T.L.; Rae, J.M.; Colacino, J.A.; Richardson, R.J. Homology models of mouse and rat estrogen receptor-α ligand-binding domain created by in silico mutagenesis of a human template: Molecular docking with 17ß-estradiol, diethylstilbestrol, and paraben analogs. Comput. Toxicol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, S.; Mäkelä, S.; Weihua, Z.; Warner, M.; Rosenlund, B.; Salmi, S.; Hovatta, O.; Gustafsson, J.A. Localization of oestrogen receptors alpha and beta in human testis. Mol. Hum. Reprod. 2001, 7, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Fietz, D.; Ratzenböck, C.; Hartmann, K.; Raabe, O.; Kliesch, S.; Weidner, W.; Klug, J.; Bergmann, M. Expression pattern of estrogen receptors α and β and G-protein-coupled estrogen receptor 1 in the human testis. Histochem. Cell Biol. 2014, 142, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Sundberg, M.; Pristovsek, N.; Ibrahim, A.; Jonsson, P.; Katona, B.; Clausson, C.M.; Zieba, A.; Ramström, M.; Söderberg, O.; et al. Insufficient antibody validation challenges oestrogen receptor beta research. Nat. Commun. 2017, 8, 15840. [Google Scholar] [CrossRef]
- Pavao, M.; Traish, A.M. Estrogen receptor antibodies: Specificity and utility in detection, localization and analyses of estrogen receptor alpha and beta. Steroids 2001, 66, 1–16. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef]
- Krege, J.H.; Hodgin, J.B.; Couse, J.F.; Enmark, E.; Warner, M.; Mahler, J.F.; Sar, M.; Korach, K.S.; Gustafsson, J.A.; Smithies, O. Generation and reproductive phenotypes of mice lacking estrogen receptor b. Proc. Natl. Acad. Sci. USA 1998, 95, 15677–15682. [Google Scholar] [CrossRef]
- Dupont, S.; Krust, A.; Gansmuller, A.; Dierich, A.; Chambon, P.; Mark, M. Effect of single and compound knockouts of estrogen receptors a (ERa) and b (ERb) on mouse reproductive phenotypes. Development 2000, 127, 4277–4291. [Google Scholar] [CrossRef]
- Wada-Hiraike, O.; Hiraike, H.; Okinaga, H.; Imamov, O.; Barros, R.P.; Morani, A.; Omoto, Y.; Warner, M.; Gustafsson, J.A. Role of estrogen receptor beta in uterine stroma and epithelium: Insights from estrogen receptor beta−/− mice. Proc. Natl. Acad. Sci. USA 2006, 103, 18350–18355. [Google Scholar] [CrossRef]
- Antal, M.C.; Krust, A.; Chambon, P.; Mark, M. Sterility and absence of histopathological defects in nonreproductive organs of a mouse ERbeta-null mutant. Proc. Natl. Acad. Sci. USA 2008, 105, 2433–2438. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Kissling, G.E.; Fieselman, K.E.; Jayes, F.L.; Gerrish, K.E.; Korach, K.S. Biological and biochemical consequences of global deletion of exon 3 from the ERa gene. FASEB J. 2010, 24, 4660–4667. [Google Scholar] [CrossRef] [PubMed]
- Antonson, P.; Omoto, Y.; Humire, P.; Gustafsson, J.Å. Generation of ERα-floxed and knockout mice using the Cre/LoxP system. Biochem. Biophys. Res. Commun. 2012, 424, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, S.C.; Korach, K.S. Estrogen receptors: New directions in the new millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Antonson, P.; Apolinário, L.M.; Shamekh, M.M.; Humire, P.; Poutanen, M.; Ohlsson, C.; Nalvarte, I.; Gustafsson, J.Å. Generation of an all-exon Esr2 deleted mouse line: Effects on fertility. Biochem. Biophys. Res. Commun. 2020, 529, 231–237. [Google Scholar] [CrossRef]
- Chen, P.; Li, B.; Ou-Yang, L. Role of estrogen receptors in health and disease. Front. Endocrinol. 2022, 13, 839005. [Google Scholar] [CrossRef]
- Tang, Z.R.; Zhang, R.; Lian, Z.X.; Deng, S.L.; Yu, K. Estrogen-receptor expression and function in female reproductive disease. Cells 2019, 8, 1123. [Google Scholar] [CrossRef]
- Schumer, S.T.; Cannistra, S.A. Granulosa cell tumor of the ovary. J. Clin. Oncol. 2003, 21, 1180–1189. [Google Scholar] [CrossRef]
- Li, X.; Tian, B.; Liu, M.; Miao, C.; Wang, D. Adult-type granulosa cell tumor of the ovary. Am. J. Cancer Res. 2022, 12, 3495–3511. [Google Scholar]
- Brink, G.J.; Groeneweg, J.W.; Hooft, L.; Zweemer, R.P.; Witteveen, P.O. Response to systemic therapies in ovarian adult granulosa cell tumors: A literature review. Cancers 2022, 14, 2998. [Google Scholar] [CrossRef]
- Ciucci, A.; Ferrandina, G.; Mascilini, F.; Filippetti, F.; Scambia, G.; Zannoni, G.F.; Gallo, D. Estrogen receptor β: Potential target for therapy in adult granulosa cell tumors? Gynecol. Oncol. 2018, 150, 158–165. [Google Scholar] [CrossRef]
- Wu, X.; Subramaniam, M.; Negron, V.; Cicek, M.; Reynolds, C.; Lingle, W.L.; Goetz, M.P.; Ingle, J.N.; Spelsberg, T.C.; Hawse, J.R. Development, characterization, and applications of a novel estrogen receptor beta monoclonal antibody. J. Cell. Biochem. 2012, 113, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Otsuka, M.; Kanaya, M.; Higo, S.; Hattori, Y.; Ozawa, H. Applicability of anti-human estrogen receptor β antibody PPZ0506 for the immunodetection of rodent estrogen receptor β proteins. Int. J. Mol. Sci. 2019, 20, 6312. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Ishii, H.; Higo, S.; Otsuka, M.; Kanaya, M.; Matsumoto, K.; Ozawa, M.; Ozawa, H. Optimization of immunohistochemical detection of rat ESR2 proteins with well-validated monoclonal antibody PPZ0506. Mol. Cell. Endocrinol. 2021, 523, 111145. [Google Scholar] [CrossRef] [PubMed]
- Klüber, P.; Meurer, S.K.; Lambertz, J.; Schwarz, R.; Zechel-Gran, S.; Braunschweig, T.; Hurka, S.; Domann, E.; Weiskirchen, R. Depletion of Lipocalin 2 (LCN2) in mice leads to dysbiosis and persistent colonization with segmented filamentous bacteria. Int. J. Mol. Sci. 2021, 22, 13156. [Google Scholar] [CrossRef]
- Scudamore, C.L. Reproductive system. In A Practical Guide to the Histology of the Mouse, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 87–107. ISBN 9781119941200. [Google Scholar] [CrossRef]
- Vidal, J.D.; Filgo, A.J. Evaluation of the estrous cycle, reproductive tract, and mammary gland in female mice. Curr. Protoc. Mouse Biol. 2017, 7, 306–325. [Google Scholar] [CrossRef]
- Ajayi, A.F.; Akhigbe, R.E. Staging of the estrous cycle and induction of estrus in experimental rodents: An update. Fertility Res. Pract. 2020, 6, 5. [Google Scholar] [CrossRef]
- Schröder, S.K.; Schüler, H.M.; Petersen, K.V.; Tesauro, C.; Knudsen, B.R.; Pedersen, F.S.; Krus, F.; Buhl, E.M.; Roeb, E.; Roderfeld, M.; et al. Genetic and molecular characterization of the immortalized murine hepatic stellate cell line GRX. Cells 2022, 11, 1504. [Google Scholar] [CrossRef]
- Madden, T. The BLAST Sequence Analysis Tool. 2002 Oct 9 [Updated 2003 Aug 13]. In The NCBI Handbook [Internet]; McEntyre, J., Ostell, J., Eds.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2002; Chapter 16. Available online: http://www.ncbi.nlm.nih.gov/books/NBK21097/ (accessed on 25 November 2022).
- Nelson, A.W.; Groen, A.J.; Miller, J.L.; Warren, A.Y.; Holmes, K.A.; Tarulli, G.A.; Tilley, W.D.; Katzenellenbogen, B.S.; Hawse, J.R.; Gnanapragasam, V.J.; et al. Comprehensive assessment of estrogen receptor beta antibodies in cancer cell line models and tissue reveals critical limitations in reagent specificity. Mol. Cell. Endocrinol. 2017, 440, 138–150. [Google Scholar] [CrossRef]
- Warner, M.; Fan, X.; Strom, A.; Wu, W.; Gustafsson, J.Å. 25 years of ERβ: A personal journey. J. Mol. Endocrinol. 2021, 68, R1–R9. [Google Scholar] [CrossRef]
- Shi, S.R.; Key, M.E.; Kalra, K.L. Antigen retrieval in formalin-fixed, paraffin-embedded tissues: An enhancement method for immunohistochemical staining based on microwave oven heating of tissue sections. J. Histochem. Cytochem. 1991, 39, 741–748. [Google Scholar] [CrossRef]
- Suurmeijer, A.J.; Boon, M.E. Notes on the application of microwaves for antigen retrieval in paraffin and plastic tissue sections. Eur. J. Morphol. 1993, 31, 144–150. [Google Scholar] [PubMed]
- Krenacs, L.; Krenacs, T.; Stelkovics, E.; Raffeld, M. Heat-induced antigen retrieval for immunohistochemical reactions in routinely processed paraffin sections. Methods Mol. Biol. 2010, 588, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Bandrowski, A.; Carr, S.; Edwards, A.; Ellenberg, J.; Lundberg, E.; Rimm, D.L.; Rodriguez, H.; Hiltke, T.; Snyder, M.; et al. A proposal for validation of antibodies. Nat. Methods 2016, 13, 823–827. [Google Scholar] [CrossRef]
- MacNeil, T.; Vathiotis, I.A.; Martinez-Morilla, S.; Yaghoobi, V.; Zugazagoitia, J.; Liu, Y.; Rimm, D.L. Antibody validation for protein expression on tissue slides: A protocol for immunohistochemistry. BioTechniques 2020, 69, 460–468. [Google Scholar] [CrossRef]
- Alkan, S.S. Monoclonal antibodies: The story of a discovery that revolutionized science and medicine. Nat. Rev. Immunol. 2004, 4, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Parray, H.A.; Shukla, S.; Samal, S.; Shrivastava, T.; Ahmed, S.; Sharma, C.; Kumar, R. Hybridoma technology a versatile method for isolation of monoclonal antibodies, its applicability across species, limitations, advancement and future perspectives. Int. Immunopharmacol. 2020, 85, 106639. [Google Scholar] [CrossRef]
- Goodpaster, T.; Randolph-Habecker, J. A flexible mouse-on-mouse immunohistochemical staining technique adaptable to biotin-free reagents, immunofluorescence, and multiple antibody staining. J. Histochem. Cytochem. 2014, 62, 197–204. [Google Scholar] [CrossRef]
- Rachman, F.; Casimiri, V.; Psychoyos, A.; Bernard, O. Immunoglobulins in the mouse uterus during the oestrous cycle. J. Reprod. Fertil. 1983, 69, 17–21. [Google Scholar] [CrossRef]
- Baker, M. Antibody anarchy: A call to order. Nature 2015, 527, 545–551. [Google Scholar] [CrossRef]
- Baker, M. Reproducibility crisis: Blame it on the antibodies. Nature 2015, 521, 274–276. [Google Scholar] [CrossRef]
- Weller, M.G. Quality issues of research antibodies. Anal. Chem. Insights 2016, 11, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Picard, D.; Kumar, V.; Chambon, P.; Yamamoto, K.R. Signal transduction by steroid hormones: Nuclear localization is differentially regulated in estrogen and glucocorticoid receptors. Cell Regul. 1990, 1, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Dapson, R.W. Macromolecular changes caused by formalin fixation and antigen retrieval. Biotech. Histochem. 2007, 82, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.A.; Strom, A.; Warner, M. Update on ERbeta. J. Steroid Biochem. Mol. Biol. 2019, 191, 105312. [Google Scholar] [CrossRef]
- Mal, R.; Magner, A.; David, J.; Datta, J.; Vallabhaneni, M.; Kassem, M.; Manouchehri, J.; Willingham, N.; Stover, D.; Vandeusen, J.; et al. Estrogen receptor beta (ERβ): A ligand activated tumor suppressor. Front. Oncol. 2020, 10, 587386. [Google Scholar] [CrossRef]
- Ramos-Vara, J.A.; Miller, M.A. When tissue antigens and antibodies get along: Revisiting the technical aspects of immunohistochemistry--the red, brown, and blue technique. Vet. Pathol. 2014, 51, 42–87. [Google Scholar] [CrossRef]
- Hewitt, S.M.; Baskin, D.G.; Frevert, C.W.; Stahl, W.L.; Rosa-Molinar, E. Controls for immunohistochemistry: The Histochemical Society’s standards of practice for validation of immunohistochemical assays. J. Histochem. Cytochem. 2014, 62, 693–697. [Google Scholar] [CrossRef]
- Ward, J.M.; Rehg, J.E. Rodent immunohistochemistry: Pitfalls and troubleshooting. Vet. Pathol. 2014, 51, 88–101. [Google Scholar] [CrossRef]
- Tsutsumi, Y. Pitfalls and caveats in applying chromogenic immunostaining to histopathological diagnosis. Cells 2021, 10, 1501. [Google Scholar] [CrossRef]
- Lu, Q.L.; Partridge, T.A. A new blocking method for application of murine monoclonal antibody to mouse tissue sections. J. Histochem. Cytochem. 1998, 46, 977–984. [Google Scholar] [CrossRef]
- Ozawa, M.; Hattori, Y.; Higo, S.; Otsuka, M.; Matsumoto, K.; Ozawa, H.; Ishii, H. Optimized mouse-on-mouse immunohistochemical detection of mouse ESR2 proteins with PPZ0506 monoclonal antibody. Acta Histochem. Cytochem. 2022, 55, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 25 November 2022).
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; reprinted 1992; Universities Federation for Animal Welfare: Wheathampstead, UK, 1959; Available online: https://caat.jhsph.edu/principles/the-principles-of-humane-experimental-technique (accessed on 25 November 2022).
- Hawse, J.R.; Carter, J.M.; Aspros, K.; Bruinsma, E.S.; Koepplin, J.W.; Negron, V.; Subramaniam, M.; Ingle, J.N.; Rech, K.L.; Goetz, M.P. Optimized immunohistochemical detection of estrogen receptor beta using two validated monoclonal antibodies confirms its expression in normal and malignant breast tissues. Breast Cancer Res. Treat. 2020, 179, 241–249. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, S.K.; Tag, C.G.; Kessel, J.C.; Antonson, P.; Weiskirchen, R. Immunohistochemical Detection of Estrogen Receptor-Beta (ERβ) with PPZ0506 Antibody in Murine Tissue: From Pitfalls to Optimization. Biomedicines 2022, 10, 3100. https://doi.org/10.3390/biomedicines10123100
Schröder SK, Tag CG, Kessel JC, Antonson P, Weiskirchen R. Immunohistochemical Detection of Estrogen Receptor-Beta (ERβ) with PPZ0506 Antibody in Murine Tissue: From Pitfalls to Optimization. Biomedicines. 2022; 10(12):3100. https://doi.org/10.3390/biomedicines10123100
Chicago/Turabian StyleSchröder, Sarah K., Carmen G. Tag, Jan C. Kessel, Per Antonson, and Ralf Weiskirchen. 2022. "Immunohistochemical Detection of Estrogen Receptor-Beta (ERβ) with PPZ0506 Antibody in Murine Tissue: From Pitfalls to Optimization" Biomedicines 10, no. 12: 3100. https://doi.org/10.3390/biomedicines10123100
APA StyleSchröder, S. K., Tag, C. G., Kessel, J. C., Antonson, P., & Weiskirchen, R. (2022). Immunohistochemical Detection of Estrogen Receptor-Beta (ERβ) with PPZ0506 Antibody in Murine Tissue: From Pitfalls to Optimization. Biomedicines, 10(12), 3100. https://doi.org/10.3390/biomedicines10123100