
Anti-Inflammatory Effects of Geniposidic Acid on Porphyromonas gingivalis-Induced Periodontitis in Mice
 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. GPA
2.2. Cell Culture and Stimulation of HGECs by P. gingivalis
2.3. RNA Extraction and Quantitative PCR Analysis
2.4. Detection of Phosphorylated Proteins
2.5. Generation of P. gingivalis-Induced Experimental Periodontitis Mouse Model
2.6. Evaluation of Alveolar Bone Level in Mouse
2.7. Histological Examination
2.8. Detection of IL-6 Levels in HGECs Supernatant and Mouse Serum
2.9. Western Blotting Analysis
2.10. Osteoclast Differentiation Assay
2.11. Statistical Analysis
3. Results
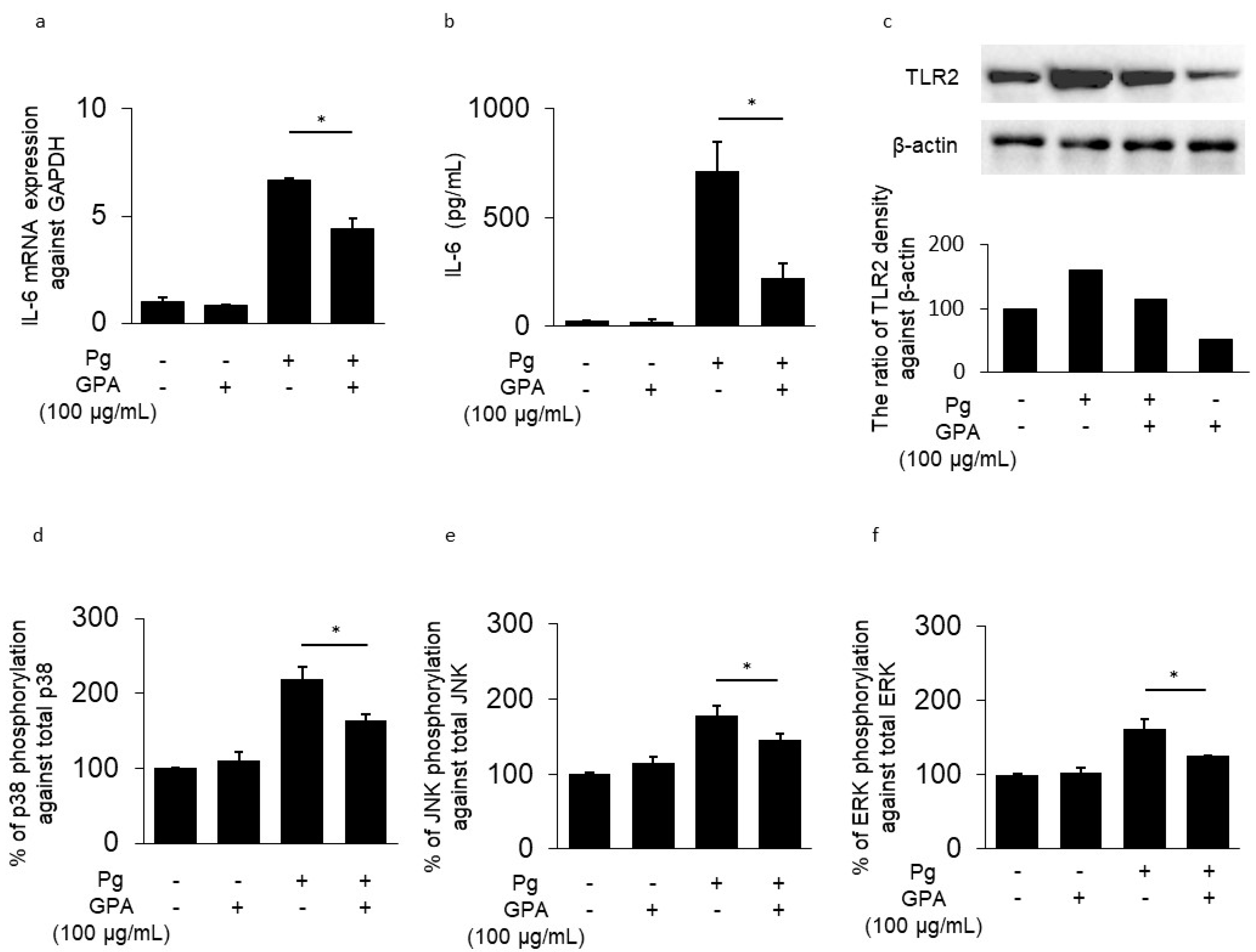
3.1. IL-6 and TLR2 Expression in HGECs
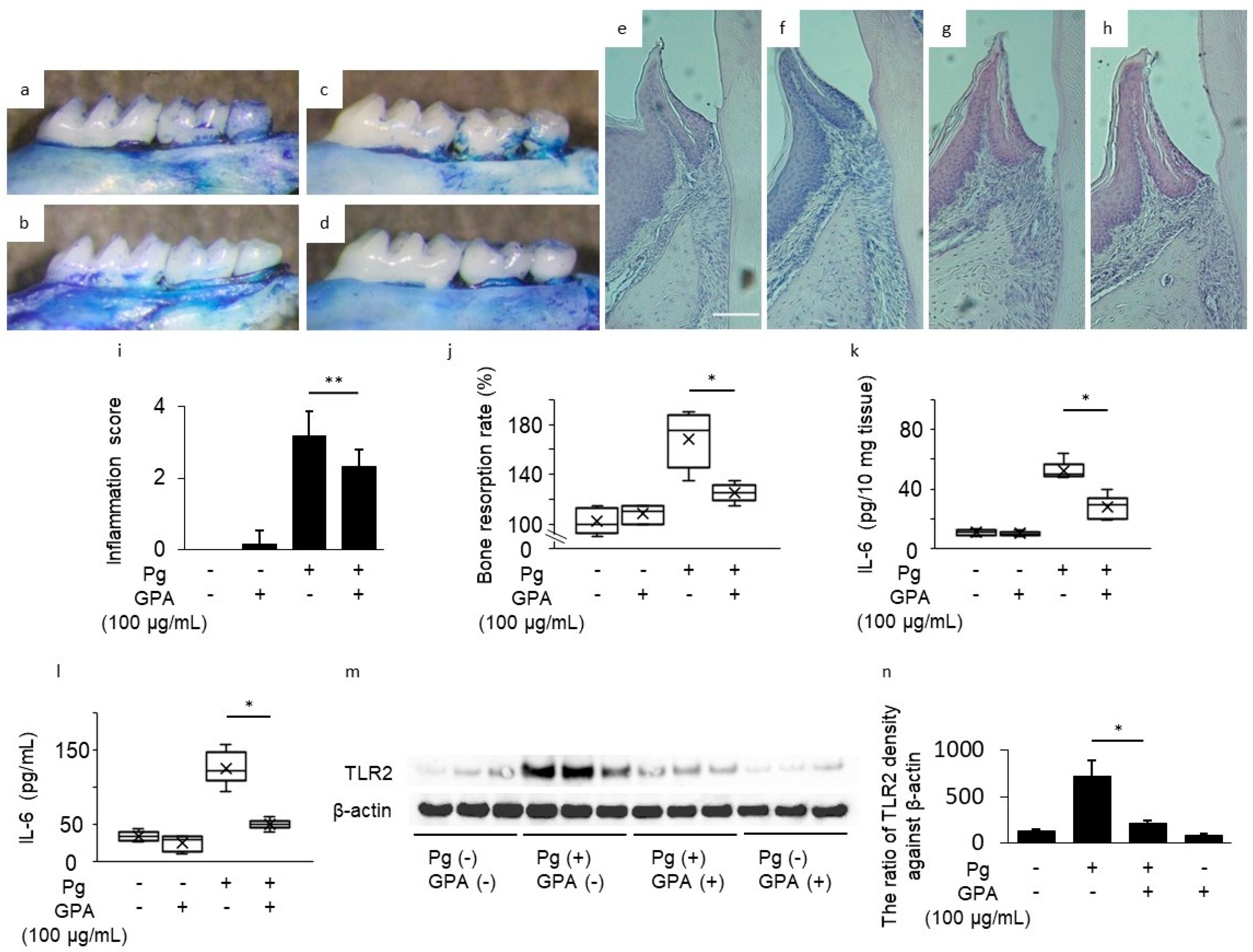
3.2. Inhibitory Effect of GPA in P. gingivalis-Induced Mouse Periodontitis Model
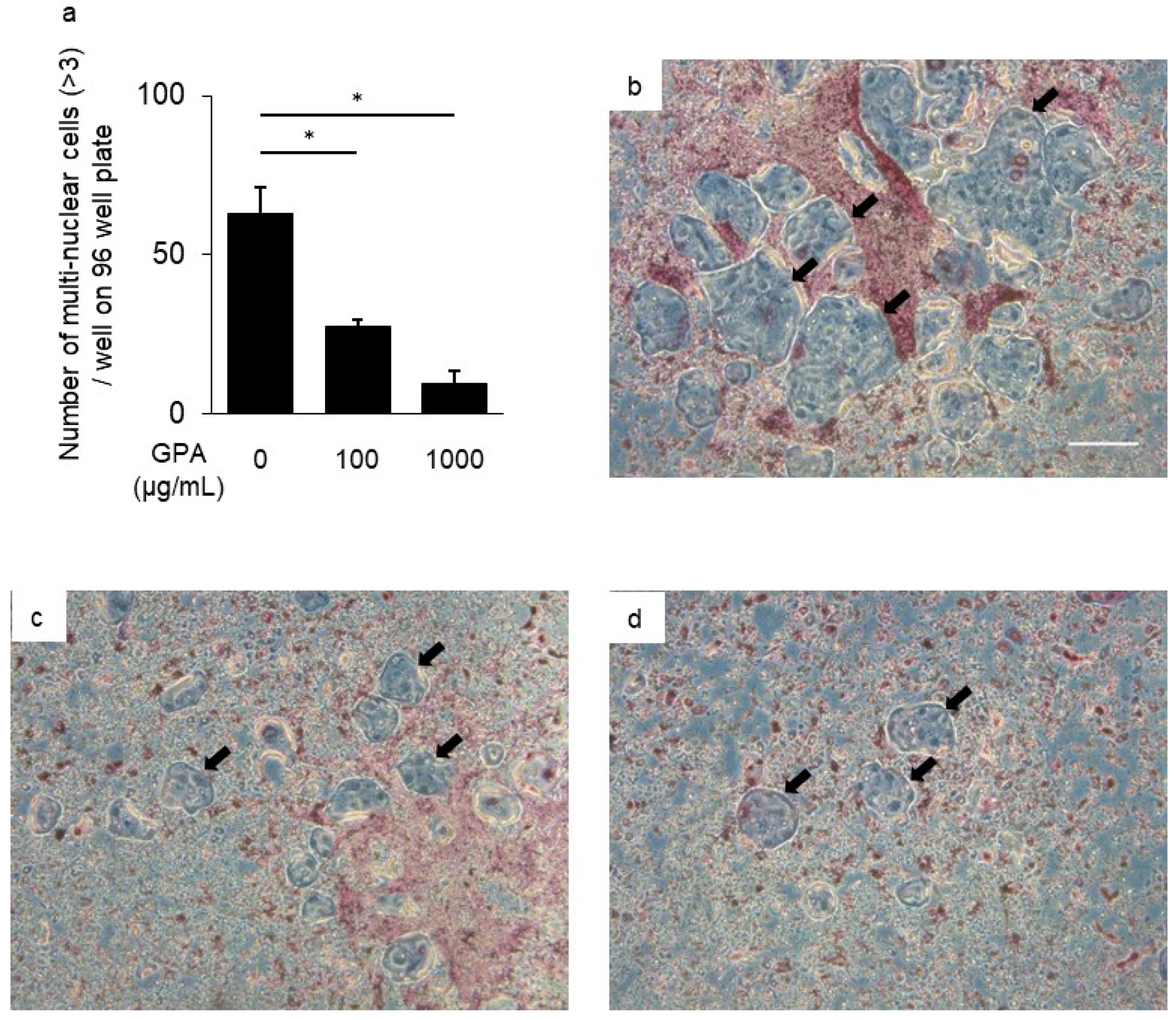
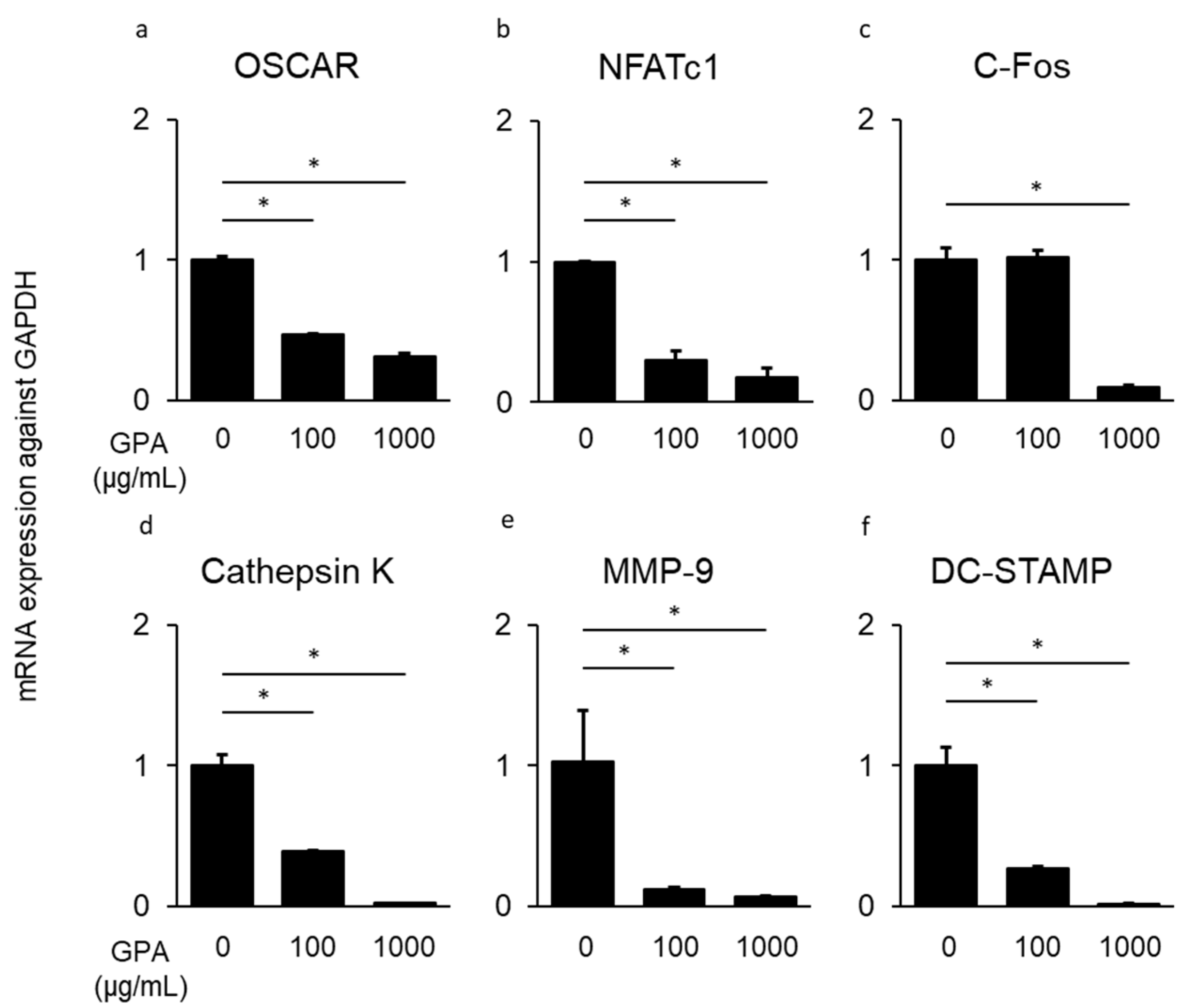
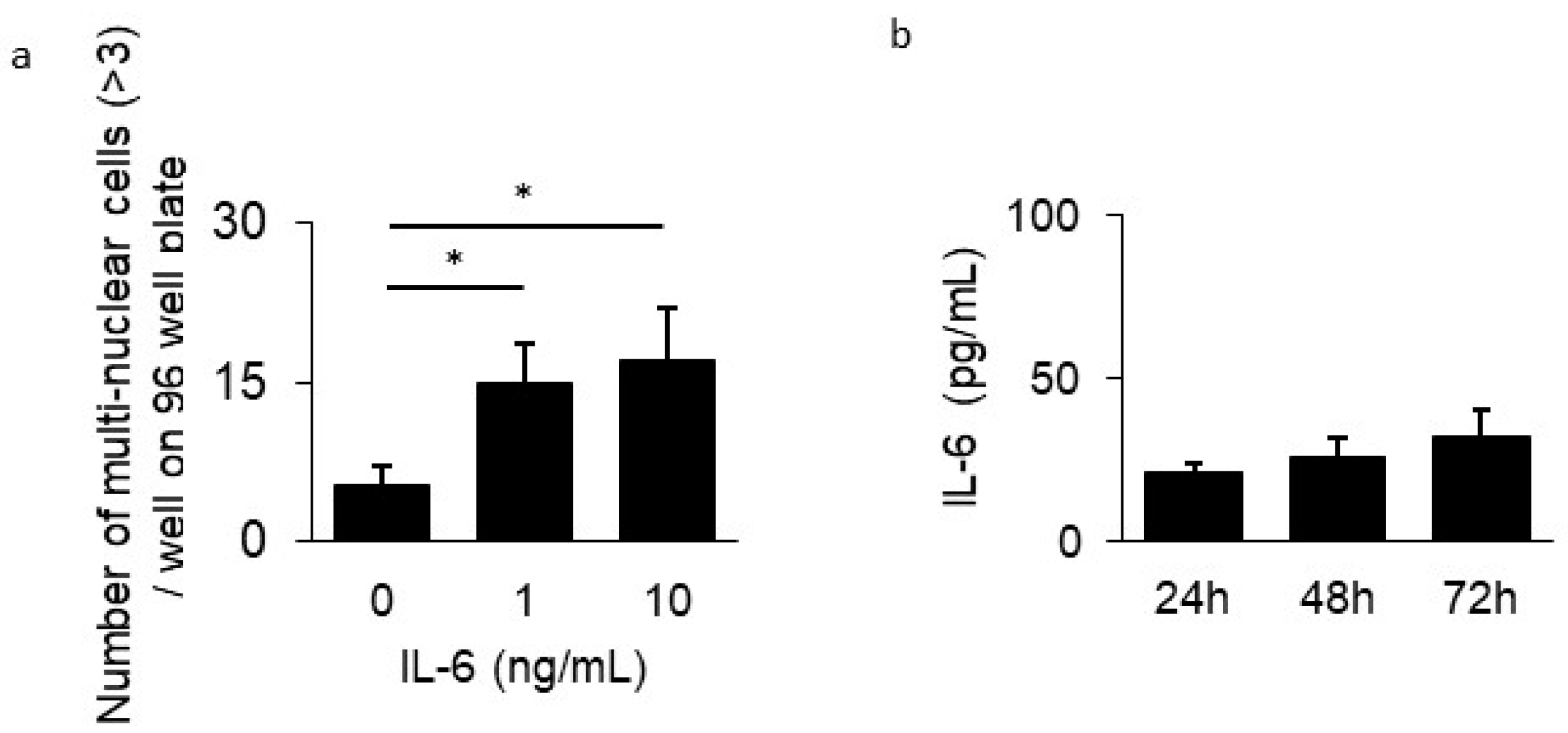
3.3. Inhibitory Effect of GPA in Osteoclast Differentiation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Socransky, S.S.; Haffajee, A.D. Implications of periodontal microbiology for the treatment of periodontal infections. Compend. Suppl. 1994, S684–685, 688–693. [Google Scholar]
- Hajishengallis, G. Immunomicrobial pathogenesis of periodontitis: Keystones, pathobionts, and host response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Amano, A. Disruption of epithelial barrier and impairment of cellular function by Porphyromonas gingivalis. Front. Biosci. 2007, 12, 3965–3974. [Google Scholar] [CrossRef] [PubMed]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 2014, 476068. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, M. RANKL and osteoimmunology in periodontitis. J. Bone Miner. Metab. 2021, 39, 82–90. [Google Scholar] [CrossRef]
- Laugisch, O.; Johnen, A.; Maldonado, A.; Ehmke, B.; Bürgin, W.; Olsen, I.; Potempa, J.; Sculean, A.; Duning, T.; Eick, S. Periodontal pathogens and associated intrathecal antibodies in early stages of Alzheimer’s disease. J. Alzheimer’s Dis. 2018, 66, 105–114. [Google Scholar] [CrossRef]
- Paraskevas, K.I. Smoking, periodontitis, and Buerger disease. Ann. Vasc. Surg. 2016, 33, 265. [Google Scholar] [CrossRef]
- Gare, J.; Kanoute, A.; Meda, N.; Viennot, S.; Bourgeois, D.; Carrouel, F. Periodontal Conditions and Pathogens Associated with Pre-Eclampsia: A Scoping Review. Int. J. Environ. Res. Public Health 2021, 18, 7194. [Google Scholar] [CrossRef]
- Bourgeois, D.; Inquimbert, C.; Ottolenghi, L.; Carrouel, F. Periodontal pathogens as risk factors of cardiovascular diseases, diabetes, rheumatoid arthritis, cancer, and chronic obstructive pulmonary disease—Is there cause for consideration? Microorganisms 2019, 7, 424. [Google Scholar] [CrossRef]
- Yoneda, M.; Naka, S.; Nakano, K.; Wada, K.; Endo, H.; Mawatari, H.; Imajo, K.; Nomura, R.; Hokamura, K.; Ono, M.; et al. Involvement of a periodontal pathogen, Porphyromonas gingivalis on the pathogenesis of non-alcoholic fatty liver disease. BMC Gastroenterol. 2012, 12, 16. [Google Scholar] [CrossRef]
- Fujita, T.; Yoshimoto, T.; Kajiya, M.; Ouhara, K.; Matsuda, S.; Takemura, T.; Akutagawa, K.; Takeda, K.; Mizuno, N.; Kurihara, H. Regulation of defensive function on gingival epithelial cells can prevent periodontal disease. Jpn. Dent. Sci. Rev. 2018, 54, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Savitri, I.J.; Ouhara, K.; Fujita, T.; Kajiya, M.; Miyagawa, T.; Kittaka, M.; Yamakawa, M.; Shiba, H.; Kurihara, H. Irsogladine maleate inhibits Porphyromonas gingivalis-mediated expression of toll-like receptor 2 and interleukin-8 in human gingival epithelial cells. J. Periodontal. Res. 2015, 50, 486–493. [Google Scholar] [CrossRef]
- Akutagawa, K.; Fujita, T.; Ouhara, K.; Takemura, T.; Tari, M.; Kajiya, M.; Matsuda, S.; Kuramitsu, S.; Mizuno, N.; Shiba, H.; et al. Glycyrrhizic acid suppresses inflammation and reduces the increased glucose levels induced by the combination of Porphyromonas gulae and ligature placement in diabetic model mice. Int. Immunopharmacol. 2019, 68, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Song, Y.R.; Park, M.H.; Chung, H.Y.; Na, H.S.; Chung, J. Catechin ameliorates Porphyromonas gingivalis-induced inflammation via the regulation of TLR2/4 and inflammasome signaling. J. Periodontol. 2020, 91, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Cheng, S.; Li, L.; Liu, Y.; Wang, D.; Liu, G. Natural anti-inflammatory compounds as drug candidates for inflammatory bowel disease. Front. Pharmacol. 2021, 12, 684486. [Google Scholar] [CrossRef]
- Kwon, S.H.; Kim, M.J.; Ma, S.X.; You, I.J.; Hwang, J.Y.; Oh, J.H.; Kim, S.Y.; Kim, H.C.; Lee, S.Y.; Jang, C.G. Eucommia ulmoides Oliv. Bark. protects against hydrogen peroxide-induced neuronal cell death in SH-SY5Y cells. J. Ethnopharmacol. 2012, 142, 337–345. [Google Scholar] [CrossRef]
- Kwon, S.H.; Ma, S.X.; Hwang, J.Y.; Ko, Y.H.; Seo, J.Y.; Lee, B.R.; Lee, S.Y.; Jang, C.G. The anti-inflammatory Activity of Eucommia ulmoides Oliv. Bark. Involves NF-kappaB Suppression and Nrf2-dependent HO-1 Induction in BV-2 Microglial Cells. Biomol. Ther. 2016, 24, 268–282. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, M.; Li, W.; Li, K.; Li, P.; Hu, Y.; Yang, Z. Protective effects of Eucommia ulmoides Oliv. bark and leaf on amyloid β-induced cytotoxicity. Environ. Toxicol. Pharmacol. 2009, 28, 342–349. [Google Scholar] [CrossRef]
- Nicolae, V.; Neamtu, B.; Picu, O.; Stefanache, M.A.M.; Cioranu, V.S.I. The comparitive evaluation of salivary biomarkers (calcium, phosphate, salivary pH) in the cario-resistance versus cario-activity. Rev. Chim. 2016, 68, 821–824. [Google Scholar]
- Ouhara, K.; Munenaga, S.; Kajiya, M.; Takeda, K.; Matsuda, S.; Sato, Y.; Hamamoto, Y.; Iwata, T.; Yamasaki, S.; Akutagawa, K.; et al. The induced RNA-binding protein, HuR, targets 3′-UTR region of IL-6 mRNA and enhances its stabilization in periodontitis. Clin. Exp. Immunol. 2018, 192, 325–336. [Google Scholar] [CrossRef]
- Shiba, H.; Tsuda, H.; Kajiya, M.; Fujita, T.; Takeda, K.; Hino, T.; Kawaguchi, H.; Kurihara, H. Neodymium-doped yttrium-aluminium–garnet laser irradiation abolishes the increase in interleukin-6 levels caused by peptidoglycan through the p38 mitogen-activated protein kinase pathway in human pulp cells. J. Endod. 2009, 35, 373–376. [Google Scholar] [CrossRef]
- Ouhara, K.; Iwasaki, Y.; Kajiya, M.; Savitri, I.J.; Kitagawa, M.; Tokunaga, N.; Shintani, T.; Ogawa, I.; Hino, T.; Fujita, T.; et al. The differential expression of mgl mRNA by Porphyromonas gingivalis affects the production of methyl mercaptan. Oral Dis. 2015, 21, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, M.; Ouhara, K.; Kajiya, M.; Munenaga, S.; Kittaka, M.; Yamasaki, S.; Takeda, K.; Takeshita, K.; Mizuno, N.; Fujita, T.; et al. Porphyromonas gingivalis infection exacerbates the onset of rheumatoid arthritis in SKG mice. Clin. Exp. Immunol. 2016, 186, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Paster, B.J.; Komatsuzawa, H.; Ernst, C.W.; Goncalves, R.B.; Sasaki, H.; Ouhara, K.; Stashenko, P.P.; Sugai, M.; Taubman, M.A. Cross-reactive adaptive immune response to oral commensal bacteria results in an induction of receptor activator of nuclear factor-kappaB ligand (RANKL)-dependent periodontal bone resorption in a mouse model. Oral Microbiol. Immunol. 2007, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Gallant, R.; Ishida, S.; Kittaka, M.; Yoshitaka, T.; Fox, D.A.; Morita, Y.; Nishida, K.; Rottapel, R.; Ueki, Y. Loss of SH3 domain-binding protein 2 function suppresses bone destruction in tumor necrosis factor-driven and collagen-induced arthritis in mice. Arthritis Rheumatol. 2015, 67, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, Y.; Ouhara, K.; Munenaga, S.; Shoji, M.; Ozawa, T.; Hisatsune, J.; Kado, I.; Kajiya, M.; Matsuda, S.; Kawai, T.; et al. Effect of Porphyromonas gingivalis infection on gut dysbiosis and resultant arthritis exacerbation in mouse model. Arthritis Res. Ther. 2020, 22, 249. [Google Scholar] [CrossRef]
- O’Brien, W.; Fissel, B.M.; Maeda, Y.; Yan, J.; Ge, X.; Gravallese, E.M.; Aliprantis, A.O.; Charles, J.F. RANK-independent osteoclast formation and bone erosion in inflammatory arthritis. Arthritis Rheumatol. 2016, 68, 2889–2900. [Google Scholar] [CrossRef]
- Song, B.; Zhang, Y.L.; Chen, L.J.; Zhou, T.; Huang, W.K.; Zhou, X.; Shao, L.Q. The role of toll-like receptors in periodontitis. Oral Dis. 2017, 23, 168–180. [Google Scholar] [CrossRef]
- Hans, M.; Hans, V.M. Toll-like receptors and their dual role in periodontitis: A review. J. Oral Sci. 2011, 53, 263–271. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef]
- Shoji, M.; Yoshimura, A.; Yoshioka, H.; Takade, A.; Takuma, Y.; Yukitake, H.; Naito, M.; Hara, Y.; Yoshida, S.; Nakayama, K. Recombinant Porphyromonas gingivalis FimA preproprotein expressed in Escherichia coli is lipidated and the mature or processed recombinant FimA protein forms a short filament in vitro. Can. J. Microbiol. 2010, 56, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Cueno, M.E.; Kamio, N.; Iinuma, T.; Hasegawa, Y.; Imai, K. Porphyromonas gingivalis Mfa1 fimbria putatively binds to TLR2 and induces both IL-6 and IL-8 production in human bronchial epithelial cells. Biochem. Biophys. Res. Commun. 2022, 589, 35–40. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K. Importance of heterogeneity in Porhyromonas gingivalis lipopolysaccharide lipid A in tissue specific inflammatory signalling. J. Oral Microbiol. 2018, 10, 1440128. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, G.; Luan, Q.; Ma, Y.; Yu, X. Porphyromonas gingivalis lipopolysaccharide-induced B cell differentiation by toll-like receptors 2 and 4. Protein Pept. Lett. 2022, 29, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Martu, M.A.; Maftei, G.A.; Sufaru, I.G.; Jelihovschi, I.; Luchian, I.; Hurjui, L.; Martu, I.; Pasarin, L. COVID-19 and Periodontal Disease-Ethiopathogenic and Clinical Implications. Rom. J. Oral Rehab. 2020, 12, 116–124. [Google Scholar]
- Zhou, Z.; Hou, J.; Mo, Y.; Ren, M.; Yang, G.; Qu, Z.; Hu, Y. Geniposidic acid ameliorates spatial learning and memory deficits and alleviates neuroinflammation via inhibiting HMGB-1 and downregulating TLR4/2 signaling pathway in APP/PS1 mice. Eur. J. Pharmacol. 2020, 869, 172857. [Google Scholar] [CrossRef]
- Li, X.; Jiang, J.; Yang, Z.; Jin, S.; Lu, X.; Qian, Y. Galangin suppresses RANKL-induced osteoclastogenesis via inhibiting MAPK and NF-kappaB signalling pathways. J. Cell Mol. Med. 2021, 25, 4988–5000. [Google Scholar] [CrossRef]
- Shi, Q.; Cao, J.; Fang, L.; Zhao, H.; Liu, Z.; Ran, J.; Zheng, X.; Li, X.; Zhou, Y.; Ge, D.; et al. Geniposide suppresses LPS-induced nitric oxide, PGE2 and inflammatory cytokine by downregulating NF-kappaB, MAPK and AP-1 signaling pathways in macrophages. Int. Immunopharmacol. 2014, 20, 298–306. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamura, T.; Zhai, R.; Takemura, T.; Ouhara, K.; Taniguchi, Y.; Hamamoto, Y.; Fujimori, R.; Kajiya, M.; Matsuda, S.; Munenaga, S.; et al. Anti-Inflammatory Effects of Geniposidic Acid on Porphyromonas gingivalis-Induced Periodontitis in Mice. Biomedicines 2022, 10, 3096. https://doi.org/10.3390/biomedicines10123096
Tamura T, Zhai R, Takemura T, Ouhara K, Taniguchi Y, Hamamoto Y, Fujimori R, Kajiya M, Matsuda S, Munenaga S, et al. Anti-Inflammatory Effects of Geniposidic Acid on Porphyromonas gingivalis-Induced Periodontitis in Mice. Biomedicines. 2022; 10(12):3096. https://doi.org/10.3390/biomedicines10123096
Chicago/Turabian StyleTamura, Tetsuya, Ruoqi Zhai, Tasuku Takemura, Kazuhisa Ouhara, Yuri Taniguchi, Yuta Hamamoto, Ryousuke Fujimori, Mikihito Kajiya, Shinji Matsuda, Syuichi Munenaga, and et al. 2022. "Anti-Inflammatory Effects of Geniposidic Acid on Porphyromonas gingivalis-Induced Periodontitis in Mice" Biomedicines 10, no. 12: 3096. https://doi.org/10.3390/biomedicines10123096
APA StyleTamura, T., Zhai, R., Takemura, T., Ouhara, K., Taniguchi, Y., Hamamoto, Y., Fujimori, R., Kajiya, M., Matsuda, S., Munenaga, S., Fujita, T., & Mizuno, N. (2022). Anti-Inflammatory Effects of Geniposidic Acid on Porphyromonas gingivalis-Induced Periodontitis in Mice. Biomedicines, 10(12), 3096. https://doi.org/10.3390/biomedicines10123096