
Evidence for Two Modes of Binding of the Negative Allosteric Modulator SB269,652 to the Dopamine D2 Receptor


Abstract
:1. Introduction
2. Methods
2.1. Molecular Biology
2.2. Oocyte Preparation
2.3. Dopamine Receptor Ligands
2.4. Electrophysiological Methods
2.5. Data Analysis
3. Results
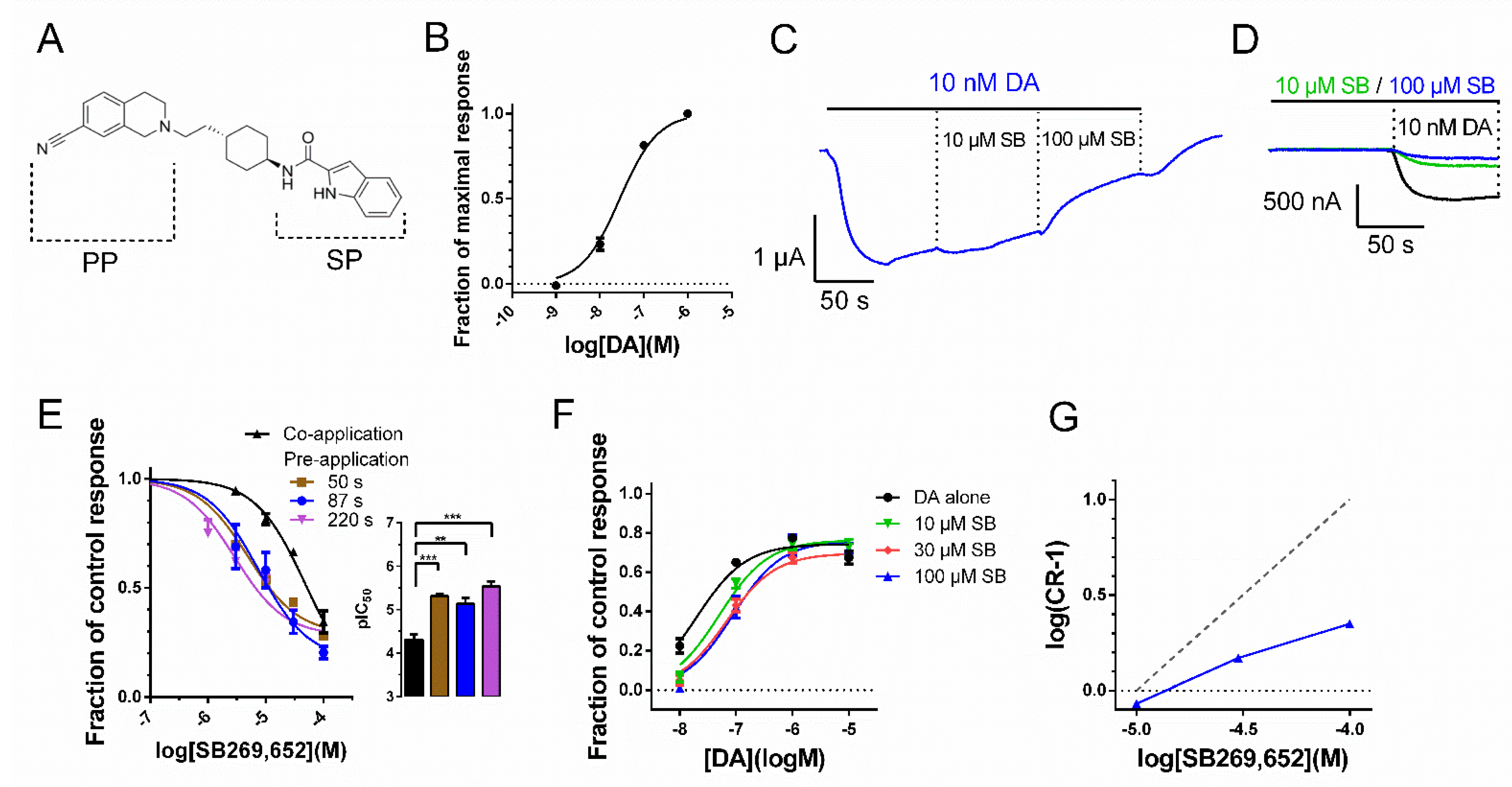
3.1. Temporal Dependency of SB269,652-Mediated D2R Response Inhibition
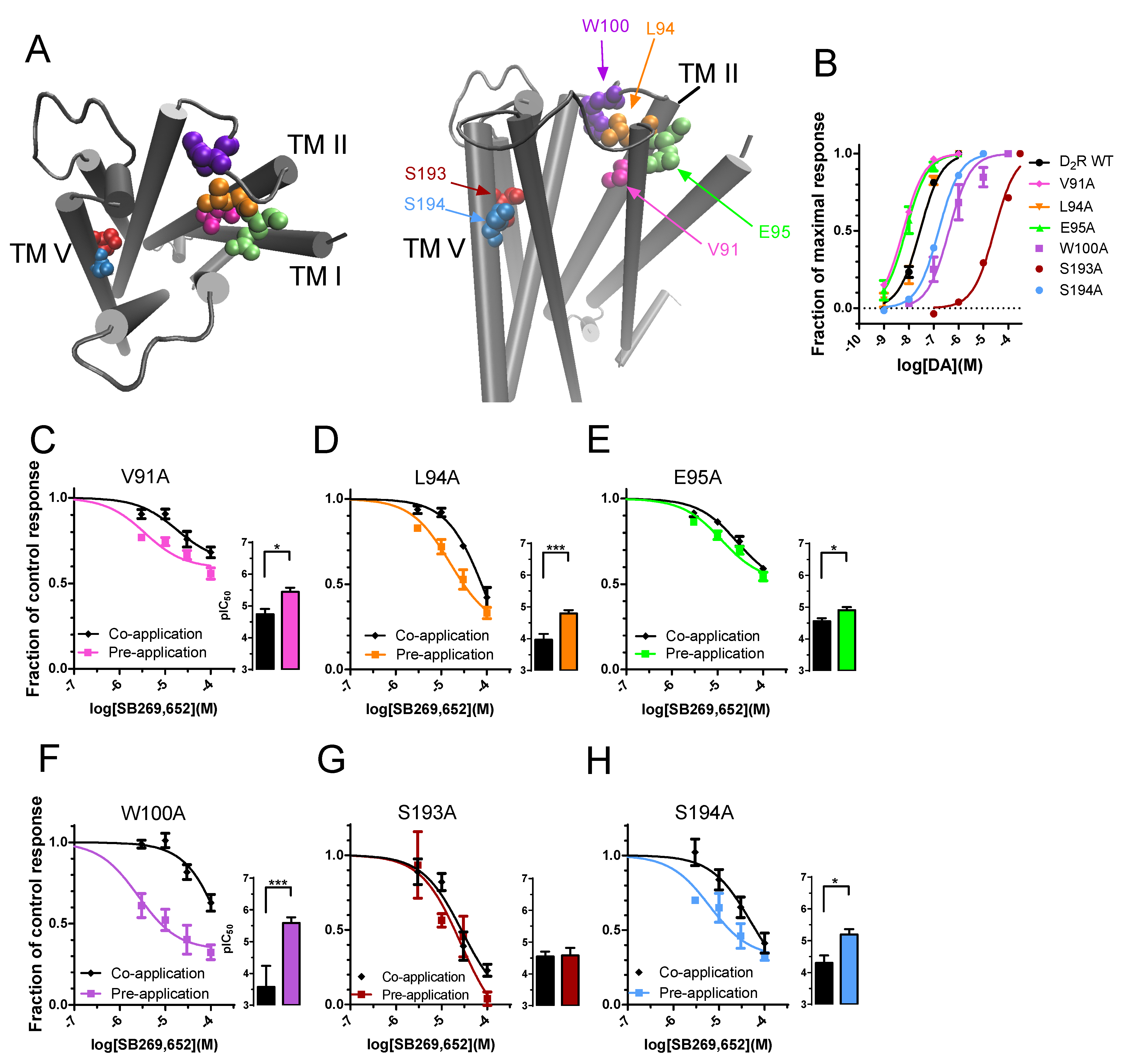
3.2. The Effect of Pre-Application on SB269,652 Potency Is Dependent on Secondary Binding Pocket Integrity
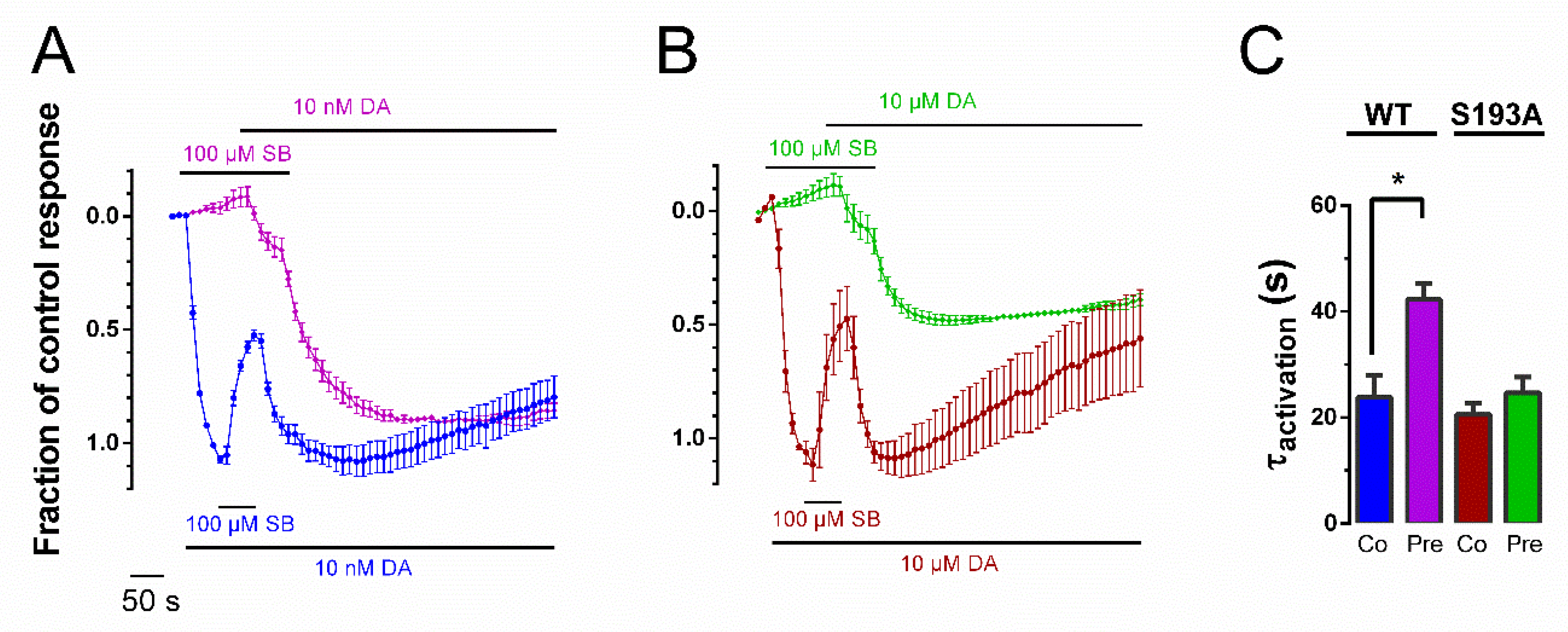
3.3. Pre-Application Slows Rate of Recovery from SB269,652 Modulation
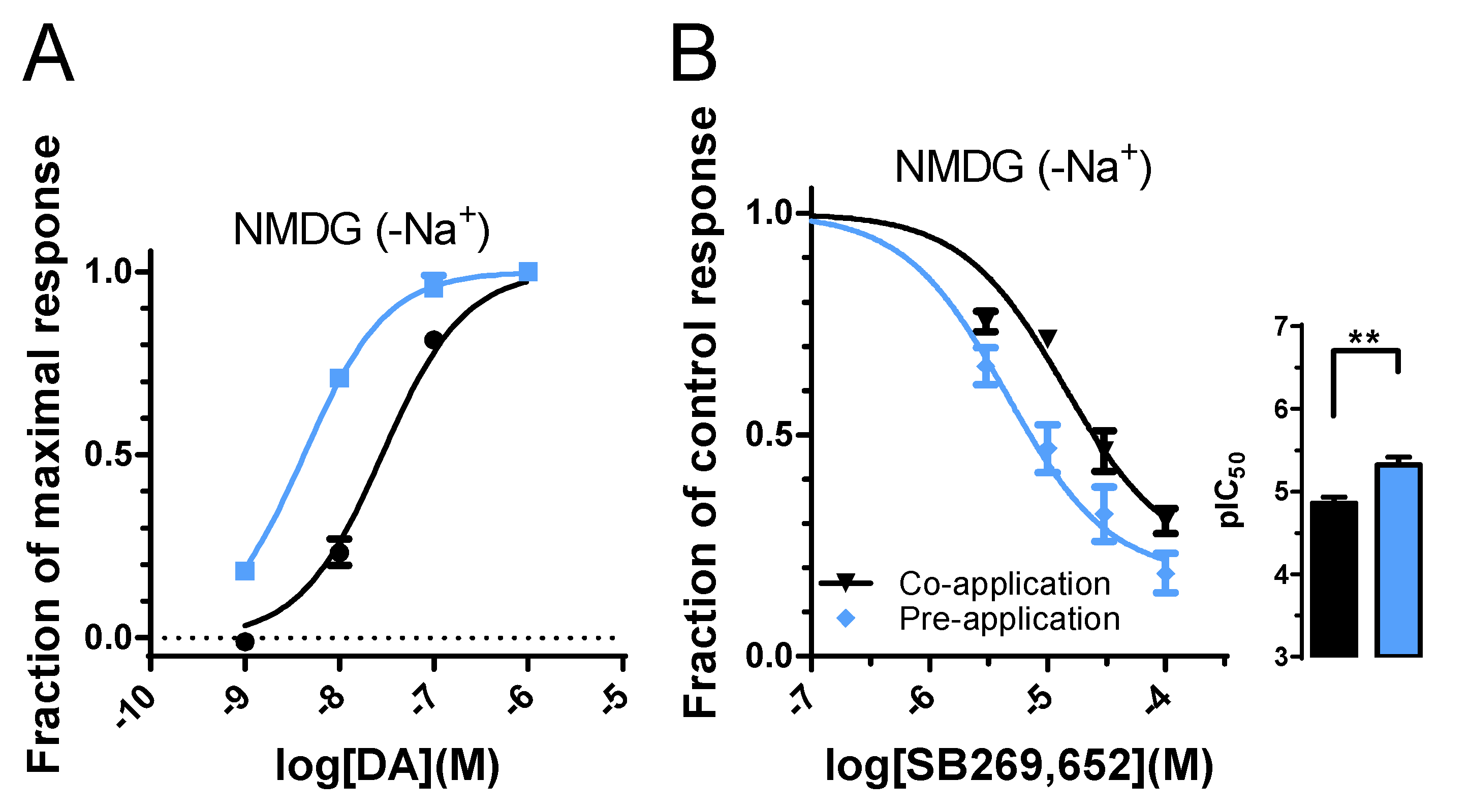
3.4. Absence of Sodium Ions Reduces Pre-Application-Induced Increase in SB269,652 Potency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.Y. New Trends in the Treatment of Schizophrenia. CNS Neurol. Disord. Drug Targets 2017, 16, 900–906. [Google Scholar] [CrossRef]
- Kopinathan, A.; Scammells, P.J.; Lane, J.R.; Capuano, B. Multivalent approaches and beyond: Novel tools for the investigation of dopamine D2 receptor pharmacology. Future Med. Chem. 2016, 8, 1349–1372. [Google Scholar] [CrossRef]
- Rossi, M.; Fasciani, I.; Marampon, F.; Maggio, R.; Scarselli, M. The First Negative Allosteric Modulator for Dopamine D. Mol. Pharmacol. 2017, 91, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Silvano, E.; Millan, M.J.; Mannoury la Cour, C.; Han, Y.; Duan, L.; Griffin, S.A.; Luedtke, R.R.; Aloisi, G.; Rossi, M.; Zazzeroni, F.; et al. The tetrahydroisoquinoline derivative SB269,652 is an allosteric antagonist at dopamine D3 and D2 receptors. Mol. Pharmacol. 2010, 78, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.R.; Donthamsetti, P.; Shonberg, J.; Draper-Joyce, C.J.; Dentry, S.; Michino, M.; Shi, L.; López, L.; Scammells, P.J.; Capuano, B.; et al. A new mechanism of allostery in a G protein-coupled receptor dimer. Nat. Chem. Biol. 2014, 10, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Mistry, S.N.; Shonberg, J.; Draper-Joyce, C.J.; Klein Herenbrink, C.; Michino, M.; Shi, L.; Christopoulos, A.; Capuano, B.; Scammells, P.J.; Lane, J.R. Discovery of a Novel Class of Negative Allosteric Modulator of the Dopamine D2 Receptor Through Fragmentation of a Bitopic Ligand. J. Med. Chem. 2015, 58, 6819–6843. [Google Scholar] [CrossRef]
- Ågren, R.; Zeberg, H.; Stępniewski, T.M.; Free, R.B.; Reilly, S.W.; Luedtke, R.R.; Århem, P.; Ciruela, F.; Sibley, D.R.; Mach, R.H.; et al. Ligand with Two Modes of Interaction with the Dopamine D2 Receptor-An Induced-Fit Mechanism of Insurmountable Antagonism. ACS Chem. Neurosci. 2020, 11, 3130–3143. [Google Scholar] [CrossRef]
- Shonberg, J.; Draper-Joyce, C.; Mistry, S.N.; Christopoulos, A.; Scammells, P.J.; Lane, J.R.; Capuano, B. Structure-activity study of N-((trans)-4-(2-(7-cyano-3,4-dihydroisoquinolin-2(1H)-yl)ethyl)cyclohexyl)-1H-indole-2-carboxamide (SB269652), a bitopic ligand that acts as a negative allosteric modulator of the dopamine D2 receptor. J. Med. Chem. 2015, 58, 5287–5307. [Google Scholar] [CrossRef]
- Draper-Joyce, C.J.; Michino, M.; Verma, R.K.; Klein Herenbrink, C.; Shonberg, J.; Kopinathan, A.; Scammells, P.J.; Capuano, B.; Thal, D.M.; Javitch, J.A.; et al. The structural determinants of the bitopic binding mode of a negative allosteric modulator of the dopamine D2 receptor. Biochem. Pharmacol. 2018, 148, 315–328. [Google Scholar] [CrossRef]
- Sahlholm, K.; Barchad-Avitzur, O.; Marcellino, D.; Gomez-Soler, M.; Fuxe, K.; Ciruela, F.; Arhem, P. Agonist-specific voltage sensitivity at the dopamine D2S receptor--molecular determinants and relevance to therapeutic ligands. Neuropharmacology 2011, 61, 937–949. [Google Scholar] [CrossRef] [Green Version]
- Papke, R.L.; Stokes, C. Working with OpusXpress: Methods for high volume oocyte experiments. Methods 2010, 51, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Ågren, R.; Stepniewski, T.M.; Zeberg, H.; Selent, J.; Sahlholm, K. Dopamine D2 Receptor Agonist Binding Kinetics-Role of a Conserved Serine Residue. Int. J. Mol. Sci. 2021, 22, 4078. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Fan, L.; Tan, L.; Chen, Z.; Qi, J.; Nie, F.; Luo, Z.; Cheng, J.; Wang, S. Haloperidol bound D(2) dopamine receptor structure inspired the discovery of subtype selective ligands. Nat. Commun. 2020, 11, 1074. [Google Scholar] [CrossRef]
- Sahlholm, K.; Zeberg, H.; Nilsson, J.; Ögren, S.O.; Fuxe, K.; Århem, P. The fast-off hypothesis revisited: A functional kinetic study of antipsychotic antagonism of the dopamine D2 receptor. Eur. Neuropsychopharmacol. 2016, 26, 467–476. [Google Scholar] [CrossRef]
- Wang, S.; Che, T.; Levit, A.; Shoichet, B.K.; Wacker, D.; Roth, B.L. Structure of the D2 dopamine receptor bound to the atypical antipsychotic drug risperidone. Nature 2018, 555, 269–273. [Google Scholar] [CrossRef]
- Fowler, J.C.; Bhattacharya, S.; Urban, J.D.; Vaidehi, N.; Mailman, R.B. Receptor conformations involved in dopamine D(2L) receptor functional selectivity induced by selected transmembrane-5 serine mutations. Mol. Pharmacol. 2012, 81, 820–831. [Google Scholar] [CrossRef] [Green Version]
- Cox, B.A.; Henningsen, R.A.; Spanoyannis, A.; Neve, R.L.; Neve, K.A. Contributions of conserved serine residues to the interactions of ligands with dopamine D2 receptors. J. Neurochem. 1992, 59, 627–635. [Google Scholar] [CrossRef]
- Stepniewski, T.M.; Mancini, A.; Ågren, R.; Torrens-Fontanals, M.; Semache, M.; Bouvier, M.; Sahlholm, K.; Breton, B.; Selent, J. Mechanistic insights into dopaminergic and serotonergic neurotransmission—Concerted interactions with helices 5 and 6 drive the functional outcome. Chem. Sci. 2021, 12, 10990–11003. [Google Scholar] [CrossRef]
- Woodward, R.; Coley, C.; Daniell, S.; Naylor, L.H.; Strange, P.G. Investigation of the role of conserved serine residues in the long form of the rat D2 dopamine receptor using site-directed mutagenesis. J. Neurochem. 1996, 66, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Coley, C.; Woodward, R.; Johansson, A.M.; Strange, P.G.; Naylor, L.H. Effect of multiple serine/alanine mutations in the transmembrane spanning region V of the D2 dopamine receptor on ligand binding. J. Neurochem. 2000, 74, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Draper-Joyce, C.J.; Verma, R.K.; Michino, M.; Shonberg, J.; Kopinathan, A.; Klein Herenbrink, C.; Scammells, P.J.; Capuano, B.; Abramyan, A.M.; Thal, D.M.; et al. The action of a negative allosteric modulator at the dopamine D2 receptor is dependent upon sodium ions. Sci. Rep. 2018, 8, 1208. [Google Scholar] [CrossRef] [Green Version]
- Ciancetta, A.; Kaur Gill, A.; Ding, T.; Karlov, D.S.; Chalhoub, G.; McCormick, P.J.; Tikhonova, I.G. Probe Confined Dynamic Mapping for G Protein-Coupled Receptor Allosteric Site Prediction. ACS Cent. Sci. 2021, 7, 1847–1862. [Google Scholar] [CrossRef]
- Free, R.B.; Cuoco, C.A.; Xie, B.; Namkung, Y.; Prabhu, V.V.; Willette BK, A.; Day, M.M.; Sanchez-Soto, M.; Lane, J.R.; Laporte, S.A.; et al. Pharmacological Characterization of the Imipridone Anticancer Drug ONC201 Reveals a Negative Allosteric Mechanism of Action at the D. Mol. Pharmacol. 2021, 100, 372–387. [Google Scholar] [CrossRef]
- Vauquelin, G. Distinct in vivo target occupancy by bivalent- and induced-fit-like binding drugs. Br. J. Pharmacol. 2017, 174, 4233–4246. [Google Scholar] [CrossRef] [Green Version]
- Ferruz, N.; Doerr, S.; Vanase-Frawley, M.A.; Zou, Y.; Chen, X.; Marr, E.S.; Nelson, R.T.; Kormos, B.L.; Wager, T.T.; Hou, X.; et al. Dopamine D3 receptor antagonist reveals a cryptic pocket in aminergic GPCRs. Sci. Rep. 2018, 8, 897. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, S.A.; Kelly, B.; Valant, C.; Michaelis, J.A.; Mastromihalis, O.; Thompson, G.; Venkatakrishnan, A.J.; Hertig, S.; Scammells, P.J.; Sexton, P.M.; et al. Cryptic pocket formation underlies allosteric modulator selectivity at muscarinic GPCRs. Nat. Commun. 2019, 10, 3289. [Google Scholar] [CrossRef]
- Moritz, A.E.; Bonifazi, A.; Guerrero, A.M.; Kumar, V.; Free, R.B.; Lane, J.R.; Verma, R.K.; Shi, L.; Newman, A.H.; Sibley, D.R. Evidence for a stereoselective mechanism for bitopic activity by extended-length antagonists of the D3 dopamine receptor. ACS Chem. Neurosci. 2020, 11, 3309–3320. [Google Scholar] [CrossRef]
- Lane, J.R.; Abramyan, A.M.; Adhikari, P.; Keen, A.C.; Lee, K.H.; Sanchez, J.; Verma, R.K.; Lim, H.D.; Yano, H.; Javitch, J.A.; et al. Distinct inactive conformations of the dopamine D2 and D3 receptors correspond to different extents of inverse agonism. eLife 2020, 9, e52189. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Fenalti, G.; Abola, E.E.; Roth, B.L.; Cherezov, V.; Stevens, R.C. Allosteric sodium in class A GPCR signaling. Trends Biochem. Sci. 2014, 39, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agren, R.; Sahlholm, K.; Nilsson, J.; Arhem, P. Point mutation of a conserved aspartate, D69, in the muscarinic M2 receptor does not modify voltage-sensitive agonist potency. Biochem. Biophys. Res. Commun. 2018, 496, 101–104. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D2R Construct | pEC50 ± SEM | n |
|---|---|---|
| WT | 7.55 ± 0.04 | 8 |
| V91A | 8.23 ± 0.02 | 4 |
| L94A | 7.55 ± 0.07 | 4 |
| E95A | 8.12 ± 0.08 | 3 |
| W100A | 6.38 ± 0.11 | 5 |
| S193A | 4.56 ± 0.05 | 3 |
| S194A | 6.80 ± 0.02 | 3 |
| WT, NMDG | 8.37 ± 0.03 | 3 |
| D2R Construct/ Condition | Co-Application pIC50 ± SEM | Co-Application IC50 (µM) | n | Pre-Application pIC50 ± SEM | Pre-Application IC50 (µM) | n | Fold Change Pre/Co-Application | Co- vs. Pre-Application (p-Value) |
|---|---|---|---|---|---|---|---|---|
| WT | 4.31 ± 0.14 | 49.0 | 3–6 | 5.13 ± 0.16 | 7.4 | 4–7 | 6.6 | 0.0023 |
| V91A | 4.74 ± 0.21 | 18.2 | 7–9 | 5.45 ± 0.15 | 3.5 | 5 | 5.1 | 0.023 |
| L94A | 3.97 ± 0.23 | 107.2 | 5–7 | 4.80 ± 0.11 | 15.8 | 5 | 6.8 | 0.0004 |
| E95A | 4.56 ± 0.10 | 27.5 | 9–10 | 4.91 ± 0.10 | 12.3 | 6 | 2.2 | 0.0349 |
| W100A | 3.58 ± 0.82 | 263.0 | 5–7 | 5.59 ± 0.21 | 2.6 | 6 | 102.3 | 0.0001 |
| S193A | 4.52 ± 0.19 | 30.2 | 6 | 4.58 ± 0.28 | 26.3 | 3 | 1.1 | 0.9236 |
| S194A | 4.30 ± 0.29 | 50.1 | 4–5 | 5.20 ± 0.19 | 6.3 | 3 | 7.9 | 0.0156 |
| WT/NMDG | 4.86 ± 0.08 | 13.8 | 5–8 | 5.35 ± 0.12 | 4.5 | 5 | 3.1 | 0.0026 |
| D2R Construct | Negative Cooperativity of SB269,652 Interaction with DA [11] a | Maximal Response Inhibition (Present Study) |
|---|---|---|
| WT | 1.23 ± 0.14 (0.54 ± 0.11) b | 0.83 ± 0.07 |
| V91A | ↓ 0.48 ± 0.16 * | ↓ (0.41 ± 0.03) *** |
| L94A | ↓ 0.70 ± 0.08 * | ↔ (0.76 ± 0.06) |
| E95A | ↓ 0.32 ± 0.14 * | ↓ (0.48 ± 0.03) ** |
| W100A | n/a | ↔ (0.66 ± 0.05) |
| S193A | ↑ (1.47 ± 0.15 *) b | ↑ (1.18 ± 0.24) * |
| S194A | ↓ 0.52 ± 0.09 * | ↔ (0.68 ± 0.06) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ågren, R.; Sahlholm, K. Evidence for Two Modes of Binding of the Negative Allosteric Modulator SB269,652 to the Dopamine D2 Receptor. Biomedicines 2022, 10, 22. https://doi.org/10.3390/biomedicines10010022
Ågren R, Sahlholm K. Evidence for Two Modes of Binding of the Negative Allosteric Modulator SB269,652 to the Dopamine D2 Receptor. Biomedicines. 2022; 10(1):22. https://doi.org/10.3390/biomedicines10010022
Chicago/Turabian StyleÅgren, Richard, and Kristoffer Sahlholm. 2022. "Evidence for Two Modes of Binding of the Negative Allosteric Modulator SB269,652 to the Dopamine D2 Receptor" Biomedicines 10, no. 1: 22. https://doi.org/10.3390/biomedicines10010022
APA StyleÅgren, R., & Sahlholm, K. (2022). Evidence for Two Modes of Binding of the Negative Allosteric Modulator SB269,652 to the Dopamine D2 Receptor. Biomedicines, 10(1), 22. https://doi.org/10.3390/biomedicines10010022