
Breast Cancer and p16: Role in Proliferation, Malignant Transformation and Progression

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Immunohistochemical (IHC) Procedure
2.3. Evaluation of IHC Staining
2.4. Statistical Data Processing
3. Results
3.1. General Characteristics
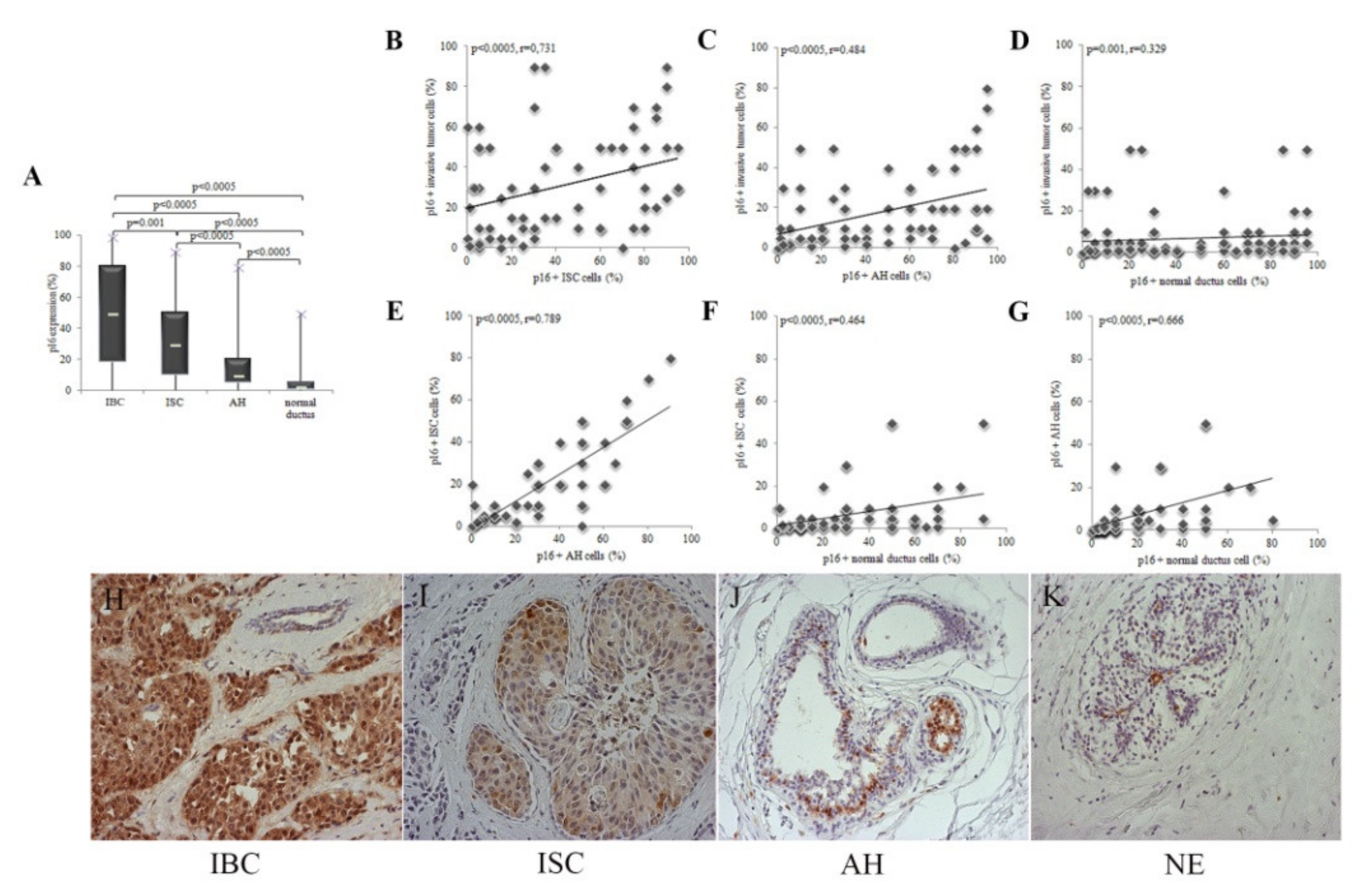
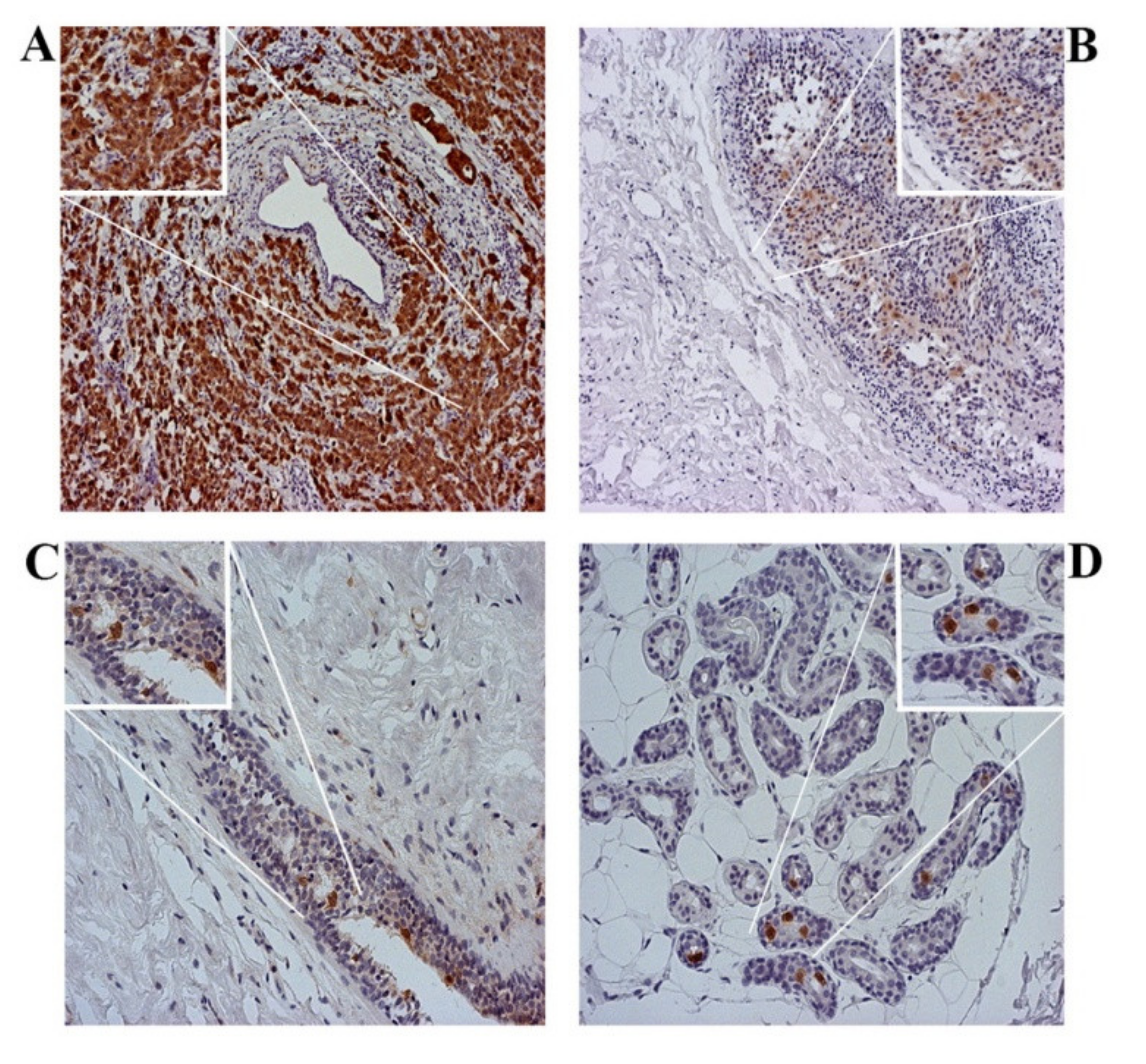
3.2. p16 Expression Increases with the Progression of Cytological Changes in the Epithelium
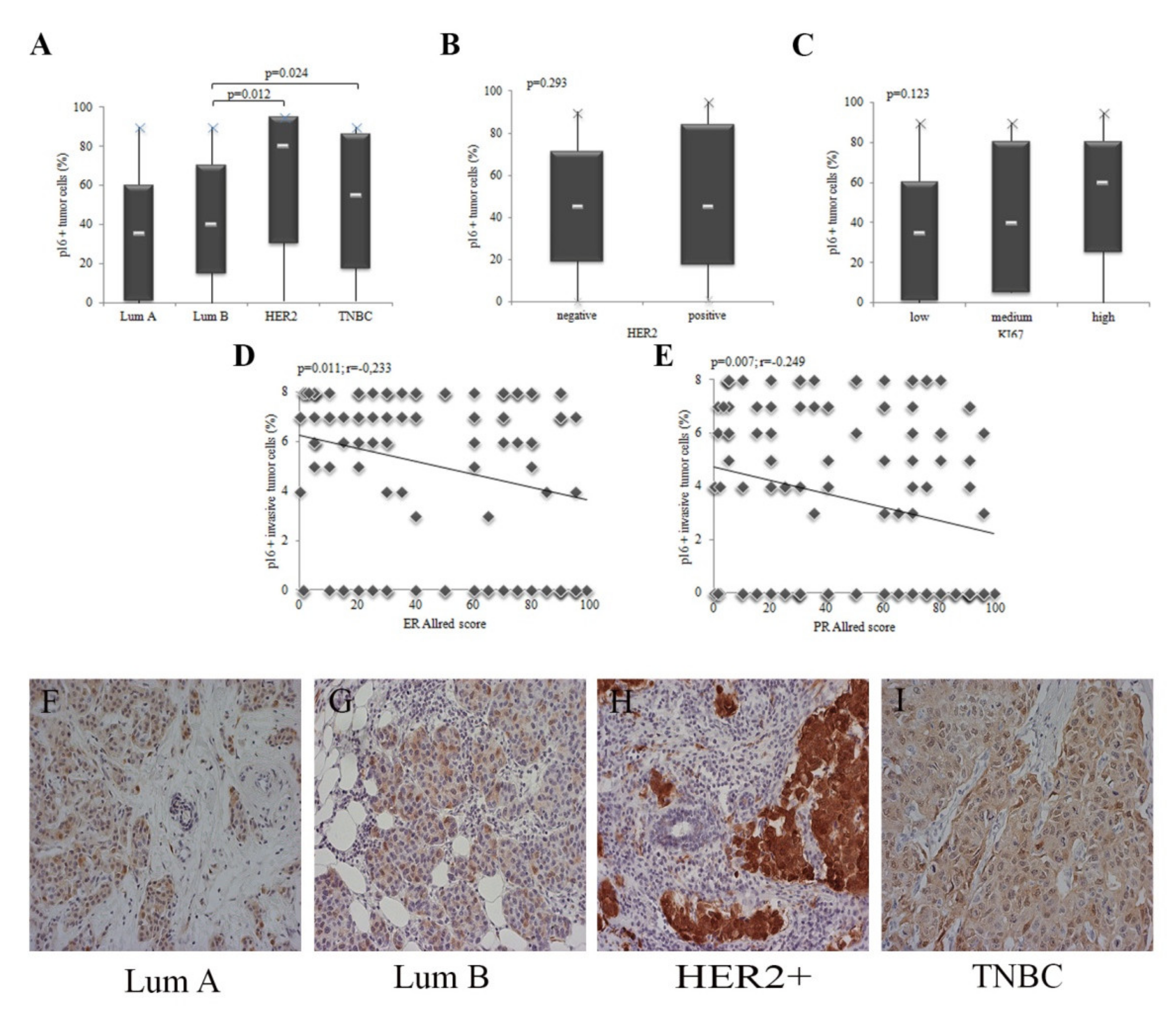
3.3. The Expression of p16 in Tumor Cells Depend on the Molecular Subtype of IBC
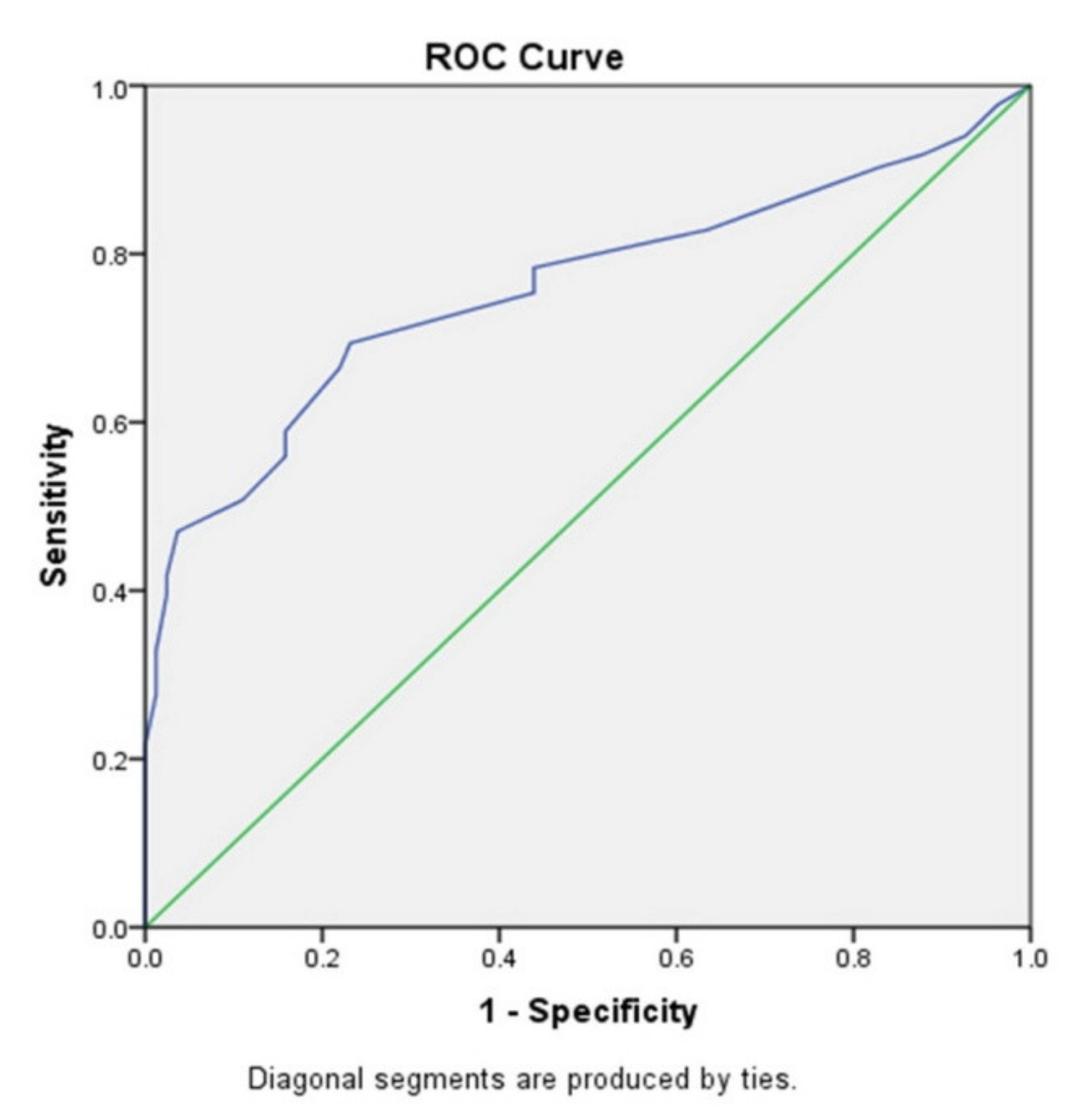
3.4. p16 Expression Is a Marker of Breast Tumor Progression
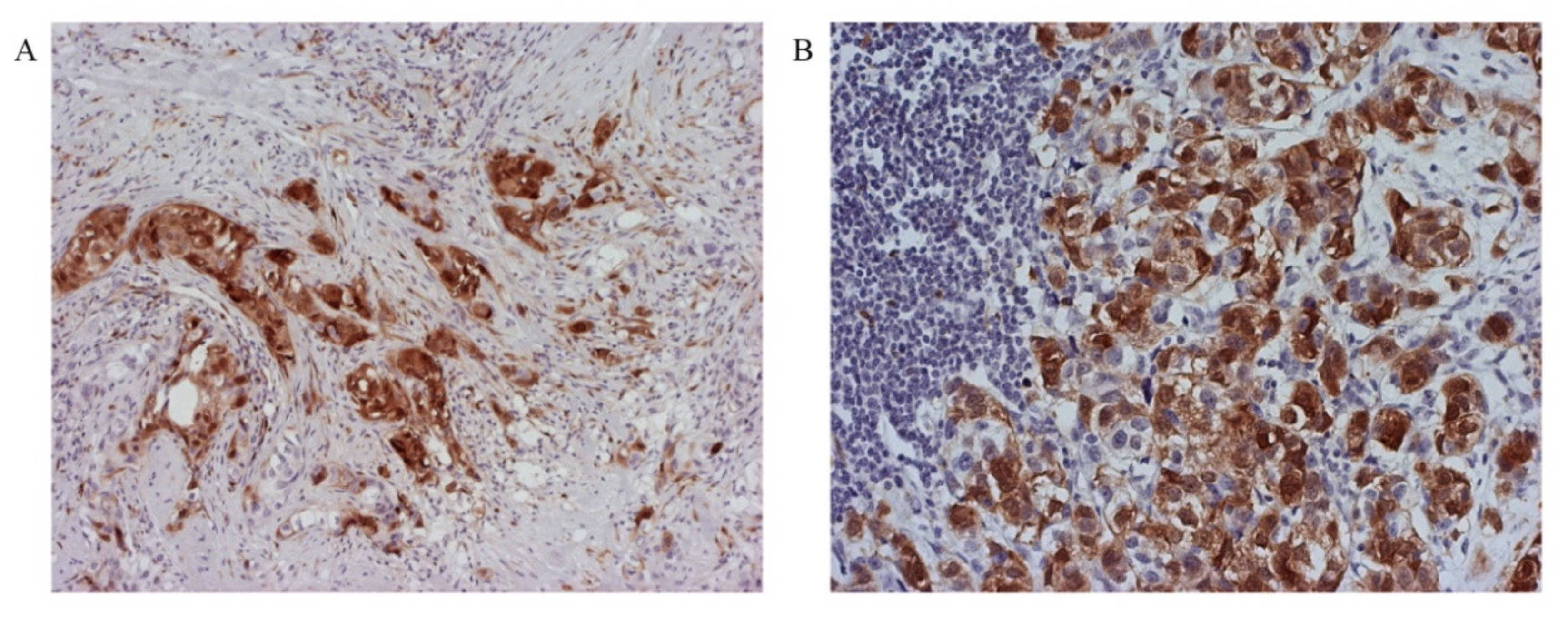
3.5. Tumor Invasiveness Depends on the Subcellular Localization of p16 Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turashvili, G.; Brogi, E. Tumor Heterogeneity in Breast Cancer. Front. Med. 2017, 4, 227. [Google Scholar] [CrossRef] [Green Version]
- Ragab, H.M.; Samy, N.; Afify, M.; El Maksoud, N.A.; Shaaban, H.M. Assessment of Ki-67 as a potential biomarker in patients with breast cancer. J. Genet. Eng. Biotechnol. 2018, 16, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Yen, M.C.; Chang, W.A.; Tsai, P.H.; Pan, Y.C.; Liao, S.H.; Kuo, P.L. CXCL17-derived CD11b(+) Gr-1(+) myeloid-derived suppressor cells contribute to lung metastasis of breast cancer through platelet-derived growth factor-BB. Breast Cancer Res. 2019, 21, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, T.M.; Kim, J.Y.; Kim, Y.J.; Sung, D.; Oh, E.; Jang, S.; Farrand, L.; Hoang, V.H.; Nguyen, C.T.; Ann, J.; et al. C-terminal HSP90 inhibitor L80 elicits anti-metastatic effects in triple-negative breast cancer via STAT3 inhibition. Cancer Lett. 2019, 447, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.; An, H.J.; Kim, S.K.; Lee, S.A.; Kim, S.; Lim, S.M.; Kim, G.M.; Sohn, J.; Moon, Y.W. Molecular mechanisms of resistance to CDK4/6 inhibitors in breast cancer: A review. Int. J. Cancer 2019, 145, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The Roles of Cyclin-Dependent Kinases in Cell-Cycle Progression and Therapeutic Strategies in Human Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topacio, B.R.; Zatulovskiy, E.; Cristea, S.; Xie, S.; Tambo, C.S.; Rubin, S.M.; Sage, J.; Kõivomägi, M.; Skotheim, J.M. Cyclin D-Cdk4,6 Drives Cell-Cycle Progression via the Retinoblastoma Protein’s C-Terminal Helix. Mol. Cell. 2019, 74, 758–770.e4. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Chowdhury, A.; Dey, S.; Roychoudhury, A.; Ganguly, A.; Bhattacharyya, D.; Roychoudhury, S. Deregulation of Rb-E2F1 axis causes chromosomal instability by engaging the transactivation function of Cdc20-anaphase-promoting complex/cyclosome. Mol. Cell Biol. 2015, 35, 356–369. [Google Scholar] [CrossRef] [Green Version]
- McNair, C.; Xu, K.; Mandigo, A.C.; Benelli, M.; Leiby, B.; Rodrigue, D.; Lindberg, J.; Gronberg, H.; Crespo, M.; De Laere, B.; et al. Differential impact of RB status on E2F1 reprogramming in human cancer. J. Clin. Investig. 2018, 128, 341–358. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Calcinotto, A.; Kohli, J.; Zagato, E.; Pellegrin, L.; Demaria, M.; Alimonti, A. Cellular Senescence: Aging, Cancer, and Injury. Physiol. Rev. 2019, 99, 1047–1078. [Google Scholar] [CrossRef] [PubMed]
- Pack, L.R.; Daigh, L.H.; Meyer, T. Putting the brakes on the cell cycle: Mechanisms of cellular growth arrest. Curr. Opin. Cell Biol. 2019, 60, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, J.; Liu, H.; Ohno, Y.; Xu, C. Targeting senescent cells and tumor therapy (Review). Int. J. Mol. Med. 2020, 46, 1603–1610. [Google Scholar] [PubMed]
- Zeng, S.; Shen, W.H.; Liu, L. Senescence and Cancer. Cancer Transl. Med. 2018, 4, 70–74. [Google Scholar]
- Scott, A.; Bai, F.; Chan, H.L.; Liu, S.; Ma, J.; Slingerland, J.M.; Robbins, D.J.; Capobianco, A.J.; Pei, X.H. p16INK4a suppresses BRCA1-deficient mammary tumorigenesis. Oncotarget 2016, 7, 84496–84507. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.H.; Liu, H.G. Immunohistochemical detection and clinicopathological significance of JARID1B/KDM5B and P16 expression in invasive ductal carcinoma of the breast. Genet. Mol. Res. 2015, 14, 5417–5426. [Google Scholar] [CrossRef]
- Gavressea, T.; Kalogeras, K.T.; Koliou, G.A.; Zagouri, F.; Lazaridis, G.; Gogas, H.; Tsigaridas, K.; Kourtas, A.; Petraki, K.; Markopoilos, C.; et al. The Prognostic Value of the Immunohistochemical Expression of Phosphorylated RB and p16 Proteins in Association with Cyclin D1 and the p53 Pathway in a Large Cohort of Patients with Breast Cancer Treated with Taxane-based Adjuvant Chemotherapy. Anticancer Res. 2017, 37, 2947–2957. [Google Scholar]
- Peurala, E.; Koivunen, P.; Haapasaari, K.M.; Bloigu, R.; Jukkola-Vuorinen, A. The prognostic significance and value of cyclin D1, CDK4 and p16 in human breast cancer. Breast Cancer Res. 2013, 15, R5. [Google Scholar] [CrossRef] [Green Version]
- Harbhajanka, A.; Lamzabi, I.; Bitterman, P.; Reddy, V.B.; Ghai, R.; Gattuso, P. Correlation of p16 Expression on Cancer and Stromal Cells With Clinicopathologic and Immunohistochemical Features of Lobular Breast Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2019, 27, 658–662. [Google Scholar] [CrossRef]
- Pare, R.; Soon, P.S.; Shah, A.; Lee, C.S. Differential expression of senescence tumour markers and its implications on survival outcomes of breast cancer patients. PLoS ONE 2019, 14, e0214604. [Google Scholar] [CrossRef]
- Wang, L.; Zhan, X.; Shen, X.; Li, M.; Yang, J.; Yu, W.; Chen, H.; Jin, B.; Mao, Z. P16 promotes the growth and mobility potential of breast cancer both in vitro and in vivo: The key role of the activation of IL-6/JAK2/STAT3 signaling. Mol. Cell Biochem. 2018, 446, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. (Eds.) Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Churchill Livingstone/Elsevier Science: London, UK, 2018. [Google Scholar]
- Tan, P.H.; Ellis, I.; Allison, K.; Brogi, E.; Fox, S.B.; Lakhani, S.; Lazar, A.J.; Morris, E.A.; Sahin, A.; Salgado, R.; et al. WHO Classification of Tumours Editorial Board. The 2019 World Health Organization classification of tumours of the breast. Histopathology 2020, 77, 181–185. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thürlimann, B.; Senn, H.J.; Panel members. Strategies for subtypes-dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar] [PubMed]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. American Society of Clinical Oncology; College of American Pathologists Recommendations for Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer. Arch. Pathol. Lab. Med. 2014, 138, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Focke, C.M.; Bürger, H.; van Diest, P.J.; Finsterbusch, K.; Gläser, D.; Korsching, E.; Decker, T. German Breast Screening Pathology Initiative. Interlaboratory variability of Ki67 staining in breast cancer. Eur. J. Cancer 2017, 84, 219–227. [Google Scholar] [CrossRef]
- Gnant, M.; Thomssen, C.; Harbeck, N. St. Gallen/Vienna 2015: A Brief Summary of the Consensus Discussion. Breast Care 2015, 10, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Yoon, N.; Yoon, G.; Park, C.K.; Kim, H.S. Stromal p16 expression is significantly increased in malignant ovarian neoplasms. Oncotarget 2016, 7, 64665–64673. [Google Scholar] [CrossRef] [Green Version]
- Schalper, K.A.; Velcheti, V.; Carvajal, D.; Wimberly, H.; Brown, J.; Pusztai, L.; Rimm, D.L. In situ tumor PD-L1 mRNA expression is associated with increased TILs and better outcome in breast carcinomas. Clin. Cancer Res. 2014, 20, 2773–2782. [Google Scholar] [CrossRef] [Green Version]
- Pare, R.; Shin, J.S.; Lee, C.S. Increased expression of senescence markers p14(ARF) and p16(INK4a) in breast cancer is associated with an increased risk of disease recurrence and poor survival outcome. Histopathology 2016, 69, 479–491. [Google Scholar] [CrossRef]
- Braig, M.; Lee, S.; Loddenkemper, C.; Rudolph, C.; Peters, A.H.; Schlegelberger, B.; Stein, H.; Dörken, B.; Jenuwein, T.; Schmitt, C.A. Oncogene induced senescence as an initial barrier in lymphoma development. Nature 2005, 436, 660–665. [Google Scholar] [CrossRef]
- Collado, M.; Blasco, M.A.; Serrano, M. Cellular senescence in cancer and aging. Cell 2007, 130, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Fry, E.A. Aberrant expression of p16INK4a in human cancers—A new biomarker? Cancer Rep. Rev. 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Michaloglou, C.; Vredeveld, L.C.; Soengas, M.S.; Denoyelle, C.; Kuilman, T.; van der Horst, C.M.; Majoor, D.M.; Shay, J.W.; Mooi, W.J.; Peeper, D.S. BRAFE600-associated senescence-like cell cycle arrest of human naevi. Nature 2005, 436, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Courtois-Cox, S.; Jones, S.L.; Cichowski, K. Many roads lead to oncogene-induced senescence. Oncogene. 2008, 27, 2801–2809. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, J.; Torrice, C.; Ramsey, M.R.; Kovalev, G.I.; Al-Regaiey, K.; Su, L.; Sharpless, N.E. Ink4a/Arf expression is a biomarker of aging. J. Clin. Investig. 2004, 114, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Sargen, M.R.; Merrill, S.L.; Chu, E.Y.; Nathanson, K.L. CDKN2A mutations with p14 loss predisposing to multiple nerve sheath tumours, melanoma, dysplastic naevi and internal malignancies: A case series and review of the literature. Br. J. Dermatol. 2016, 175, 785–789. [Google Scholar] [CrossRef]
- Joselow, A.; Lynn, D.; Terzian, T.; Box, N.F. Senescence-Like Phenotypes in HumanNevi. Methods Mol. Biol. 2017, 1534, 175–184. [Google Scholar]
- Romagosa, C.; Simonetti, S.; Serrano, C. Senescence markers in Schwannomas. Virchows Arch. 2009, 455, 372. [Google Scholar]
- Perrone, F.; Tabano, S.; Colombo, F.; Dagrada, G.; Birindelli, S.; Gronchi, A.; Colecchia, M.; Pierotti, M.A.; Pilotti, S. p15INK4b, p14ARF, and p16 inactivation in sporadic and neurofibromatosis type 1-related malignant peripheral nerve sheath tumors. Clin. Cancer Res. 2003, 9, 4132–4138. [Google Scholar]
- Damsky, W.; Micevic, G.; Meeth, K.; Muthusamy, V.; Curley, D.P.; Santhanakrishnan, M.; Erdelyi, I.; Platt, J.T.; Huang, L.; Theodosakis, N.; et al. mTORC1 activation blocks BrafV600E-induced growth arrest but is insufficient for melanoma formation. Cancer Cell 2015, 27, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.Y.; Furth, E.E.; Mick, R.; Koh, J.; Takayama, T.; Niitsu, Y.; Enders, G.H. p16(INK4a) expression begins early in human colon neoplasia and correlates inversely with markers of cell proliferation. Gastroenterology 2000, 119, 929–942. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Bamberger, A.M.; Rieck, G.; Kelp, B.; Löning, T. Overexpression of the p16 cell cycle inhibitor in breast cancer is associated with a more malignant phenotype. Breast Cancer Res. Treat. 2001, 67, 61–70. [Google Scholar] [CrossRef]
- Di Vinci, A.; Perdelli, L.; Banelli, B.; Salvi, S.; Casciano, I.; Gelvi, I.; Allemanni, G.; Margallo, E.; Gatteschi, B.; Romani, M. p16(INK4a) promoter methylation and protein expression in breast fibroadenoma and carcinoma. Int. J. Cancer 2005, 114, 414–421. [Google Scholar] [CrossRef]
- Zhao, P.; Mao, X.; Talbot, I.C. Aberrant cytological localization of p16 and CDK4 in colorectal epithelia in the normal adenoma carcinoma sequence. World J. Gastroenterol. 2006, 12, 6391–6396. [Google Scholar] [CrossRef]
- Hilliard, N.J.; Krahl, D.; Sellheyer, K. p16 expression differentiates between desmoplastic Spitz nevus and desmoplastic melanoma. J. Cutan. Pathol. 2009, 36, 753–759. [Google Scholar] [CrossRef]
- Bai, P.; Xiao, X.; Zou, J.; Cui, L.; Bui Nguyen, T.M.; Liu, J.; Xiao, J.; Chang, B.; Wu, J.; Wang, H. Expression of p14(ARF), p15(INK4b), p16(INK4a) and skp2 increases during esophageal squamous cell cancer progression. Exp. Ther. Med. 2012, 3, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Rosen, D.G.; Yao, J.L.; Huang, J.; Liu, J. Expression of p14ARF, p15INK4b, p16, and DCR2 increases during prostate cancer progression. Mod. Pathol. 2006, 19, 1339–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasanac, D.; Stojkovic-Filipovic, J.; Bosic, M.; Tomanovic, N.; Manojlovic-Gacic, E. Expression of G1/S-cyclins and cyclin-dependent kinase inhibitors in actinic keratosis and squamous cell carcinoma. J. Cutan. Pathol. 2016, 43, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Lynch, B.C.; Lathrop, S.L.; Ye, D.; Ma, T.Y.; Cerilli, L.A. Expression of the p16(INK4a) gene product in premalignant and malignant epithelial lesions of the gallbladder. Ann. Diagn. Pathol. 2008, 12, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Zouheir, Y.; Fechtali, T.; Elgnaoui, N. Human Papillomavirus Genotyping and p16(INK4a) Expression in Cervical Lesions: A Combined Test to Avoid Cervical Cancer Progression. J. Cancer Prev. 2016, 21, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Mendaza, S.; Fernández-Irigoyen, J.; Santamaría, E.; Zudaire, T.; Guarch, R.; Guerrero-Setas, D.; Vidal, A.; Santos-Salas, J.; Matias-Guiu, X.; Ausín, K.; et al. Absence of Nuclear p16 Is a Diagnostic and Independent Prognostic Biomarker in Squamous Cell Carcinoma of the Cervix. Int. J. Mol. Sci. 2020, 21, 2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebok, P.; Roming, M.; Kluth, M.; Koop, C.; Özden, C.; Taskin, B.; Hussein, K.; Lebeau, A.; Witzel, I.; Wölber, L.; et al. p16 overexpression and 9p21 deletion are linked to unfavorable tumor phenotype in breast cancer. Oncotarget 2016, 7, 81322–81331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, S.P.; Wang, H.L.; Peng, W.; Liu, H.J.; Hou, L.; Zhang, B. Aberrant expression and correlative analysis of P16 in breast cancers. Beijing Da Xue Xue Bao. 2012, 44, 755–759. [Google Scholar] [PubMed]
- Sugianto, J.; Sarode, V.; Peng, Y. Ki-67 expression is increased in p16-expressing triple-negative breast carcinoma and correlates with p16 only in p53-negative tumors. Hum. Pathol. 2014, 45, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Subhawong, A.P.; Subhawong, T.; Nassar, H.; Kouprina, N.; Begum, S.; Vang, R.; Westra, W.H.; Argani, P. Most basal-like breast carcinomas demonstrate the same Rb-/p16+ immunophenotype as the HPV-related poorly differentiated squamous cell carcinomas which they resemble morphologically. Am. J. Surg. Pathol. 2009, 33, 163–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, M.L.; Berman, H.K.; Miller, C.; Kozakiewicz, K.; Chew, K.; Moore, D.; Rabban, J.; Chen, Y.Y.; Kerlikowske, K.; Tlsty, T.D. Abrogated response to cellular stress identifies DCIS associated with subsequent tumor events and defines basal-like breast tumors. Cancer Cell. 2007, 12, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Kerlikowske, K.; Molinaro, A.M.; Gauthier, M.L.; Berman, H.K.; Waldman, F.; Bennington, J.; Sanchez, H.; Jimenez, C.; Stewart, K.; Chew, K.; et al. Biomarker Expression and Risk of Subsequent Tumors After Initial Ductal Carcinoma In Situ Diagnosis. J. Natl. Cancer Inst. 2010, 102, 627–637. [Google Scholar] [CrossRef]
- Shin, E.; Jung, W.H.; Koo, J.S. Expression of p16 and pRB in invasive breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 8209–8217. [Google Scholar]
- Ortiz, A.B.; Garcia, D.; Vicente, Y.; Palka, M.; Bellas, C.; Martin, P. Prognostic significance of cyclin D1 protein expression and gene amplification in invasive breast carcinoma. PLoS ONE 2017, 12, e0188068. [Google Scholar] [CrossRef]
- Shan, M.; Zhang, X.; Liu, X.; Qin, Y.; Liu, T.; Liu, Y.; Wang, J.; Zhong, Z.; Zhang, Y.; Geng, J.; et al. P16 and p53 play distinct roles in different subtypes of breast cancer. PLoS ONE 2013, 8, e76408. [Google Scholar]
- Abou-Bakr, A.A.; Eldweny, H.I. p16 expression correlates with basal-like triple-negative breast carcinoma. Ecancermedicalscience 2013, 7, 317. [Google Scholar]
- Bohn, O.L.; Fuertes-Camilo, M.; Navarro, L.; Saldivar, J.; Sanchez-Sosa, S. p16INK4a expression in basal-like breast carcinoma. Int. J. Clin. Exp. Pathol. 2010, 3, 600–607. [Google Scholar] [PubMed]
- Kobierzycki, C.; Grzegrzolka, J.; Glatzel-Plucinska, N.; Piotrowska, A.; Wojnar, A.; Smolarz, B.; Romanowicz, H.; Dziegiel, P. Expression of p16 and SATB1 in Invasive Ductal Breast Cancer—A Preliminary Study. In Vivo 2018, 32, 731–736. [Google Scholar] [CrossRef]
- Evangelou, K.; Bramis, J.; Peros, I.; Zacharatos, P.; Dasiou-Plakida, D.; Kalogeropoulos, N.; Asimacopoulos, P.J.; Kittas, C.; Marinos, E.; Gorgoulis, V.G. Electron microscopy evidence that cytoplasmic localization of the p16(INK4A) ‘nuclear’ cyclindependent kinase inhibitor (CKI) in tumor cells is specific and not an artifact. A study in non-small cell lung carcinomas. Biotech. Histochem. 2004, 79, 5–10. [Google Scholar] [CrossRef]
- Romagosa, C.; Simonetti, S.; López-Vicente, L.; Mazo, A.; Lleonart, M.E.; Castellvi, J.; Ramon y Cajal, S. p16(Ink4a) overexpression in cancer: A tumor suppressor gene associated with senescence and high-grade tumors. Oncogene 2011, 30, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Gump, J.; Stokoe, D.; McCormick, F. Phosphorylation of p16 correlates with Cdk4 association. J. Biol. Chem. 2003, 278, 6619–6622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, K.; Landberg, G. Subcellular localization, modification and protein complex formation of the cdk-inhibitor p16 in Rb-functional and Rb-inactivated tumor cells. Int. J. Cancer 2006, 118, 1120–1125. [Google Scholar] [CrossRef]
- Souza-Rodrigues, E.; Estanyol, J.M.; Friedrich-Heineken, E.; Olmedo, E.; Vera, J.; Canela, N.; Brun, S.; Agell, N.; Hübscher, U.; Bachs, O.; et al. Proteomic analysis of p16 binding proteins. Proteomics 2007, 7, 4102–4111. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.W.; Wu, J.; Cai, L.; Liu, B.Y.; Gao, Y.; Chen, G.Q.; Fu, G.H. Expression of anion exchanger 1 sequestrates p16 in the cytoplasm in gastric and colonic adenocarcinoma. Neoplasia 2007, 9, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Chung, T.; Do, S.I.; Na, K.; Kim, G.; Jeong, Y.I.; Kim, Y.W.; Kim, H.S. Stromal p16 Overexpression in Gastric-type Mucinous Carcinoma of the Uterine Cervix. Anticancer Res. 2018, 38, 3551–3558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Schrauder, M.; Oswald, U.; Knoll, C.; Sellberg, P.; Palmqvist, R.; Niedobitek, G.; Brabletz, T.; Kirchner, T. The invasion front of human colorectal adenocarcinomas shows co-localization of nuclear beta-catenin, cyclin D1, and p16INK4A and is a region of low proliferation. Am. J. Pathol. 2001, 159, 1613–1617. [Google Scholar] [CrossRef]
- Natarajan, E.; Saeb, M.; Crum, C.P.; Woo, S.B.; McKee, P.H.; Rheinwald, J.G. Co-expression of p16(INK4A) and laminin 5 gamma2 by microinvasive and superficial squamous cell carcinomas in vivo and by migrating wound and senescent keratinocytes in culture. Am. J. Pathol. 2003, 163, 477–491. [Google Scholar] [CrossRef]
- Svensson, S.; Nilsson, K.; Ringberg, A.; Landberg, G. Invade or proliferate? Two contrasting events in malignant behavior governed by p16(INK4a) and an intact Rb pathway illustrated by a model system of basal cell carcinoma. Cancer Res. 2003, 63, 1737–1742. [Google Scholar] [PubMed]
- Palmqvist, R.; Rutegard, J.N.; Bozoky, B.; Landberg, G.; Stenling, R. Human colorectal cancers with an intact p16/ cyclin D1/pRb pathway have up-regulated p16 expression and decreased proliferation in small invasive tumor clusters. Am. J. Pathol. 2000, 157, 1947–1953. [Google Scholar] [CrossRef] [Green Version]
- Moritani, S.; Ichihara, S.; Hasegawa, M.; Iwakoshi, A.; Murakami, S.; Sato, T.; Okamoto, T.; Mori, Y.; Kuhara, H.; Silverberg, S.G. Stromal p16 expression differentiates endometrial polyp from endometrial hyperplasia. Virchows Arch. 2012, 461, 141–148. [Google Scholar] [CrossRef]
- Davalos, A.R.; Coppe, J.P.; Campisi, J.; Desprez, P.Y. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2010, 29, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Coppe, J.P.; Desprez, P.Y.; Krtolica, A.; Campisi, J. The senescence-associated secretory phenotype: The dark side of tumor suppression. Annu. Rev. Pathol. 2010, 5, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Coppe, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008, 6, 2853–2868. [Google Scholar] [CrossRef]
- Orr, F.W.; Wang, H.H. Tumor cell interactions with the microvasculature: A rate-limiting step in metastasis. Surg. Oncol. Clin. N. Am. 2001, 10, 357–381. [Google Scholar] [CrossRef]
- Baker, D.J.; Alimirah, F.; van Deursen, J.M.; Campisi, J.; Hildesheim, J. Oncogenic senescence: A multi-functional perspective. Oncotarget 2017, 8, 27661–27672. [Google Scholar] [CrossRef] [Green Version]
- Lecot, P.; Alimirah, F.; Desprez, P.Y.; Campisi, J.; Wiley, C. Context-dependent effects of cellular senescence in cancer development. Br. J. Cancer 2016, 114, 1180–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppé, J.P.; Kauser, K.; Campisi, J.; Beauséjour, C.M. Secretion of vascular endothelial growth factor by primary human fibroblasts at senescence. J. Biol. Chem. 2006, 281, 29568–29574. [Google Scholar] [CrossRef] [Green Version]
- Kuilman, T.; Michaloglou, C.; Vredeveld, L.C.; Douma, S.; van Doorn, R.; Desmet, C.J.; Aarden, L.A.; Mooi, W.J.; Peeper, D.S. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell 2008, 133, 1019–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Choi, Y.W.; Lee, J.; Soh, E.Y.; Kim, J.H.; Park, T.J. Senescent tumor cells lead the collective invasion in thyroid cancer. Nat. Commun. 2017, 8, 15208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Au, B.; Kulkarni, M.; Shen, Y.; Lim, K.J.; Maimaiti, J.; Wong, C.K.; Luijten, M.N.H.; Chong, H.C.; Lim, E.H.; et al. Chromosomal instability-induced senescence potentiates cell non-autonomous tumourigenic effects. Oncogenesis 2018, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Ritschka, B.; Storer, M.; Mas, A.; Heinzmann, F.; Ortells, M.C.; Morton, J.P.; Sansom, O.J.; Zender, L.; Keyes, W.M. The senescence-associated secretory phenotype induces cellular plasticity and tissue regeneration. Genes Dev. 2017, 31, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Krtolica, A.; Parrinello, S.; Lockett, S.; Desprez, P.Y.; Campisi, J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: A link between cancer and aging. Proc. Natl. Acad. Sci. USA 2001, 98, 12072–12077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Hornsby, P.J. Senescent human fibroblasts increase the early growth of xenograft tumors via matrix metalloproteinase secretion. Cancer Res. 2007, 67, 3117–3126. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, S.; Coppe, J.P.; Krtolica, A.; Campisi, J. Stromal–epithelial interactions in aging and cancer: Senescent fibroblasts alter epithelial cell differentiation. J. Cell Sci. 2005, 118 Pt 3, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.K.; Chuang, E.Y.; Little, J.B.; Yuan, Z.M. Cellular mechanisms for low-dose ionizing radiation-induced perturbation of the breast tissue microenvironment. Cancer Res. 2005, 65, 6734–6744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.; Yoshizuka, N.; New, L.; Moser, B.A.; Li, Y.; Liao, R.; Xie, C.; Chen, J.; Deng, Q.; Yamout, M.; et al. PRAK is essential for ras-induced senescence and tumor suppression. Cell 2007, 128, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Shendrik, I.; Peacocke, M.; Peehl, D.; Buttyan, R.; Ikeguchi, E.F.; Katz, A.E.; Benson, M.C. Expression of senescence-associated beta-galactosidase in enlarged prostates from men with benign prostatic hyperplasia. Urology 2000, 56, 160–166. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Murakami, M.; Qian, L.W.; Sato, N.; Nagai, E.; Matsumoto, K.; Nakamura, T.; Tanaka, M. Radiation to stromal fibroblasts increases invasiveness of pancreatic cancer cells through tumor–stromal interactions. Cancer Res. 2004, 64, 3215–3222. [Google Scholar] [CrossRef] [Green Version]
- Coppe, J.P.; Boysen, M.; Sun, C.H.; Wong, B.J.; Kang, M.K.; Park, N.H.; Desprez, P.Y.; Campisi, J.; Krtolica, A. A role for fibroblasts in mediating the effects of tobacco-induced epithelial cell growth and invasion. Mol. Cancer Res. 2008, 6, 1085–1098. [Google Scholar] [CrossRef] [Green Version]
- Székely, B.; Silber, A.L.; Pusztai, L. New Therapeutic Strategies for Triple-Negative Breast Cancer. Oncology 2017, 31, 130–137. [Google Scholar]
- Yoon, E.C.; Wilson, P.; Zuo, T.; Pinto, M.; Cole, K.; Harigopal, M. High frequency of p16 and SOX10 coexpression but not androgen receptor expression in triple-negative breast cancers. Hum. Pathol. 2020, 102, 13–22. [Google Scholar] [CrossRef]
- Bogina, G.S.; Lunardi, G.; Marcolini, L.; Brunelli, M.; Bortesi, L.; Marconi, M.; Coati, F.; Valerio, M.; Guerriero, M.; Massocco, A.; et al. P16 but not retinoblastoma expression is related to clinical outcome in no-special-type triple-negative breast carcinomas. Mod. Pathol. 2014, 27, 204–213. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | N | % | |
|---|---|---|---|
| Side | left | 66 | 44.9 |
| right | 81 | 55.1 | |
| Histological type | lobular | 18 | 12.4 |
| ductal | 123 | 84.8 | |
| other | 4 | 2.8 | |
| Histological grade | HG1 | 17 | 11.9 |
| HG2 | 73 | 51 | |
| HG3 | 53 | 37.1 | |
| Nuclear grade | NG1 | 17 | 15.2 |
| NG2 | 64 | 57.1 | |
| NG3 | 31 | 27.7 | |
| Tumor necrosis | absent | 26 | 21.7 |
| present | 94 | 78.3 | |
| Perineural invasion | absent | 101 | 68.7 |
| present | 46 | 31.3 | |
| Lymphatic invasion | absent | 72 | 48.9 |
| present | 75 | 51.1 | |
| Vascular invasion | absent | 113 | 76.9 |
| present | 34 | 23.1 | |
| Molecular subtypes | Luminal A | 30 | 20.4 |
| Luminal B | 76 | 51.7 | |
| HER2+ | 19 | 12.9 | |
| TNBC | 22 | 15 | |
| HER2 | negative | 115 | 79.3 |
| positive | 30 | 20.7 | |
| Ki67 | low | 30 | 20.9 |
| medium | 42 | 29.4 | |
| high | 71 | 49.7 | |
| T status | T1 | 48 | 35.8 |
| T2 | 64 | 47.8 | |
| T3 | 9 | 6.7 | |
| T4 | 13 | 9.7 | |
| N status | N0 | 50 | 37.3 |
| N1 | 48 | 35.8 | |
| N2 | 19 | 14.2 | |
| N3 | 17 | 12.7 | |
| Molecular Subtypes | N | % |
|---|---|---|
| Luminal A | 30 | 20.40 |
| Luminal B | 76 | 51.70 |
| HER2+ | 19 | 12.93 |
| TNBC | 22 | 14.97 |
| Total | 147 | 100.0 |
| Variables | p16 Cut off 17.5 | Chi-Square | df | P | ||
|---|---|---|---|---|---|---|
| − | + | |||||
| Mononuclear infiltrate | absent | 2 (10%) | 2 (2.7%) | 7.959 | 3 | 0.047 |
| low | 13 (65%) | 30 (41.1%) | ||||
| medium | 5 (25%) | 30 (41.1%) | ||||
| high | 0 (0%) | 11 (15.1%) | ||||
| Stromal fibroblasts | absent | 2 (9.1%) | 2 (3.2%) | 8.120 | 3 | 0.044 |
| low | 7 (31.8%) | 32 (50.8%) | ||||
| medium | 3 (13.6%) | 17 (27%) | ||||
| high | 10 (45.5%) | 12 (19%) | ||||
| Histological type | lobular | 4 (13.8%) | 6 (6.7%) | 1.410 | 2 | 0.494 |
| ductal | 24 (82.8%) | 80 (89.9%) | ||||
| other | 1 (3.4%) | 3 (3.4%) | ||||
| Histological grade | HG1 | 2 (8.3%) | 8 (10.5%) | 2.709 | 2 | 0.258 |
| HG2 | 15 (62.5%) | 33 (43.4%) | ||||
| HG3 | 7 (29.2%) | 35 (46.1%) | ||||
| Nuclear grade | NG1 | 1 (4.8%) | 11 (14.8%) | 2.468 | 2 | 0.291 |
| NG2 | 15 (71.4%) | 40 (54.1%) | ||||
| NG3 | 5 (23.8%) | 23 (31.1%) | ||||
| Tumor necrosis | absent | 6 (23,1%) | 20 (21.3%) | 0.000 | 1 | 1.000 |
| present | 20 (76,9%) | 74 (78.7%) | ||||
| Perineural invasion | absent | 18 (60%) | 65 (72.2%) | 1.055 | 1 | 0.304 |
| present | 12 (40%) | 25 (27.8%) | ||||
| Lymphatic invasion | absent | 10 (33.3%) | 45 (50%) | 1.891 | 1 | 0.169 |
| present | 20 (66.7%) | 45 (50%) | ||||
| Vascular invasion | absent | 23 (76.7%) | 66 (73.3%) | 0.014 | 1 | 0.904 |
| present | 7 (23.3%) | 24 (26.7%) | ||||
| Molecular subtypes | Luminal A | 4 (14.3%) | 11 (13.8%) | 3.029 | 3 | 0.387 |
| Luminal B | 19 (67.9%) | 41 (51.2%) | ||||
| HER2+ | 2 (7.1%) | 11 (13.8%) | ||||
| TNBC | 3 (10.7%) | 17 (21.2%) | ||||
| HER2 | negative | 24 (82.8%) | 67 (75.3%) | 0.334 | 1 | 0.563 |
| positive | 5 (17.2%) | 22 (24.7%) | ||||
| Ki67 | low | 4 (16.7%) | 11 (15.3%) | 7.217 | 2 | 0.027 |
| medium | 11 (45.8%) | 14 (19.4%) | ||||
| high | 9 (37.5%) | 47 (65.3%) | ||||
| T status | T1 | 6 (22.2%) | 30 (37%) | 3.802 | 3 | 0.284 |
| T2 | 14 (51.9%) | 40 (49.4%) | ||||
| T3 | 3 (11.1%) | 3 (3.7%) | ||||
| T4 | 4 (14.8%) | 8 (9.9%) | ||||
| N status | N0 | 6 (33.3%) | 24 (40.7%) | 0.959 | 3 | 0.811 |
| N1 | 9 (50%) | 22 (37.3%) | ||||
| N2 | 2 (11.1%) | 8 (13.6%) | ||||
| N3 | 1 (5.6%) | 5 (8.4%) | ||||
| p16 Nuclear vs. Cytoplasmic Expression | ||||
|---|---|---|---|---|
| p16 IBC n (%) | p16 ISC n (%) | p16 AH n (%) | p16 NE n (%) | |
| negative | 3 (2.3) | 3 (3.8) | 3 (3.7) | 3 (2.8) |
| nuclear | 28 (21.1) | 20 (25.6) | 38 (46.3) | 99 (90.8) |
| cytoplasmic | 102 (76.6) | 55 (70.6) | 41 (50) | 7 (6.4) |
| Total | 133 (100) | 78 (100) | 82 (100) | 109 (100) |
| Variables | p16 Expression in IBC | Chi-Square | df | P | ||
|---|---|---|---|---|---|---|
| Nuclear | Cytoplasmic | |||||
| Mononuclear infiltrate | absent | 1 (5%) | 2 (2.9%) | 16.236 | 6 | 0.013 |
| low | 14 (70%) | 26 (37.7%) | ||||
| medium | 5 (25%) | 30 (43.5%) | ||||
| high | 0 (0%) | 11 (15.9%) | ||||
| Stromal fibroblasts | absent | 1 (5%) | 2 (3.2%) | 11.638 | 6 | 0.071 |
| low | 8 (40%) | 31 (50%) | ||||
| medium | 6 (30%) | 13 (21%) | ||||
| high | 5 (25%) | 16 (25.8%) | ||||
| Tumor necrosis | absent | 7 (58.3%) | 19 (17.6%) | 6.609 | 2 | 0.037 |
| present | 5 (41.7%) | 89 (82.4%) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jovanovic, D.V.; Mitrovic, S.L.; Milosavljevic, M.Z.; Ilic, M.B.; Stankovic, V.D.; Vuletic, M.S.; Dimitrijevic Stojanovic, M.N.; Milosev, D.B.; Azanjac, G.L.; Nedeljkovic, V.M.; et al. Breast Cancer and p16: Role in Proliferation, Malignant Transformation and Progression. Healthcare 2021, 9, 1240. https://doi.org/10.3390/healthcare9091240
Jovanovic DV, Mitrovic SL, Milosavljevic MZ, Ilic MB, Stankovic VD, Vuletic MS, Dimitrijevic Stojanovic MN, Milosev DB, Azanjac GL, Nedeljkovic VM, et al. Breast Cancer and p16: Role in Proliferation, Malignant Transformation and Progression. Healthcare. 2021; 9(9):1240. https://doi.org/10.3390/healthcare9091240
Chicago/Turabian StyleJovanovic, Dalibor V., Slobodanka L. Mitrovic, Milos Z. Milosavljevic, Milena B. Ilic, Vesna D. Stankovic, Milena S. Vuletic, Milica N. Dimitrijevic Stojanovic, Danijela B. Milosev, Goran L. Azanjac, Vladica M. Nedeljkovic, and et al. 2021. "Breast Cancer and p16: Role in Proliferation, Malignant Transformation and Progression" Healthcare 9, no. 9: 1240. https://doi.org/10.3390/healthcare9091240
APA StyleJovanovic, D. V., Mitrovic, S. L., Milosavljevic, M. Z., Ilic, M. B., Stankovic, V. D., Vuletic, M. S., Dimitrijevic Stojanovic, M. N., Milosev, D. B., Azanjac, G. L., Nedeljkovic, V. M., & Radovanovic, D. (2021). Breast Cancer and p16: Role in Proliferation, Malignant Transformation and Progression. Healthcare, 9(9), 1240. https://doi.org/10.3390/healthcare9091240