
Does Whole Grain Consumption Alter Gut Microbiota and Satiety?

Abstract
:1. Introduction
2. Background: Whole Grains
3. Defining Whole Grains
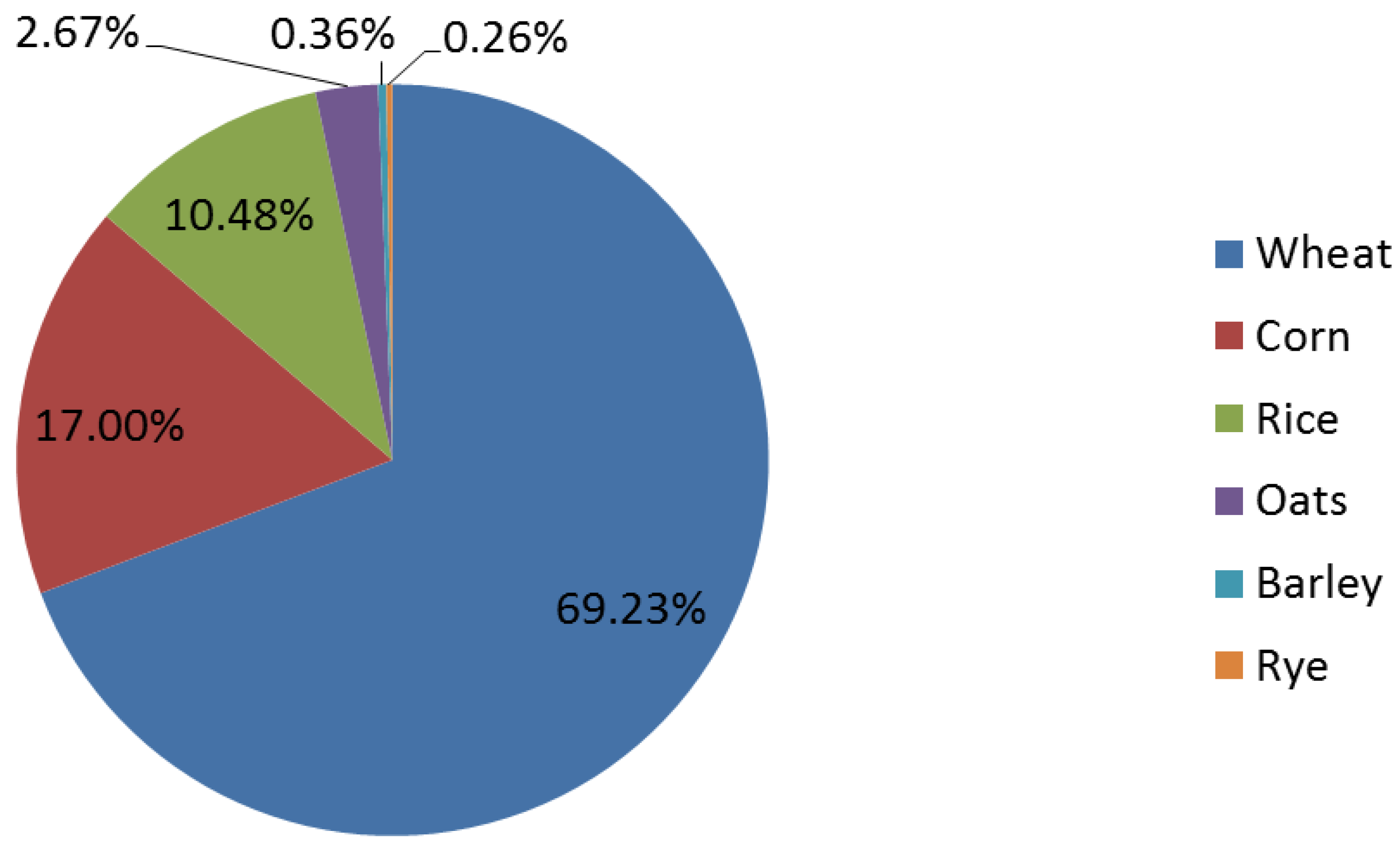
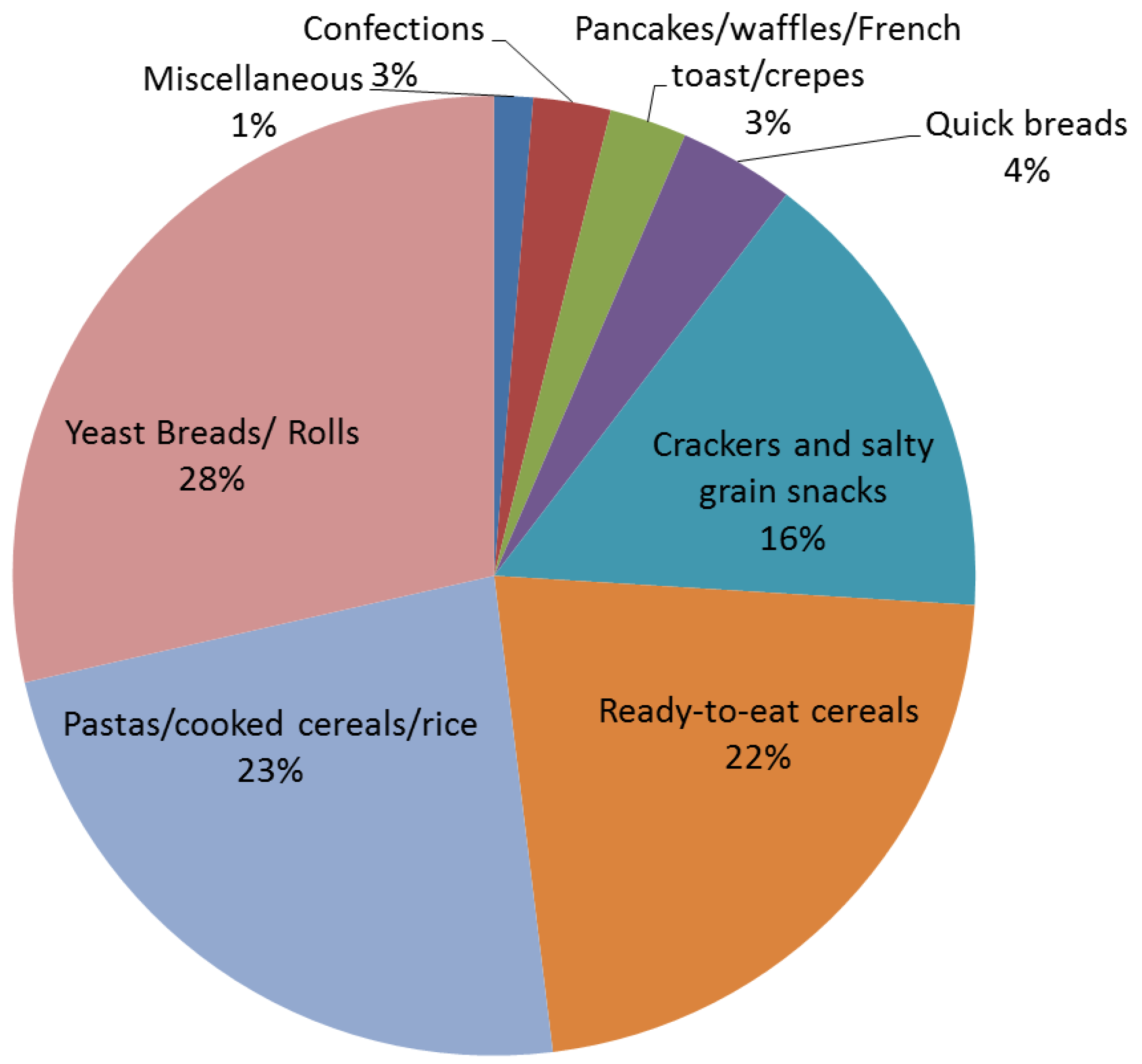
4. Whole Grain Consumption in the United States
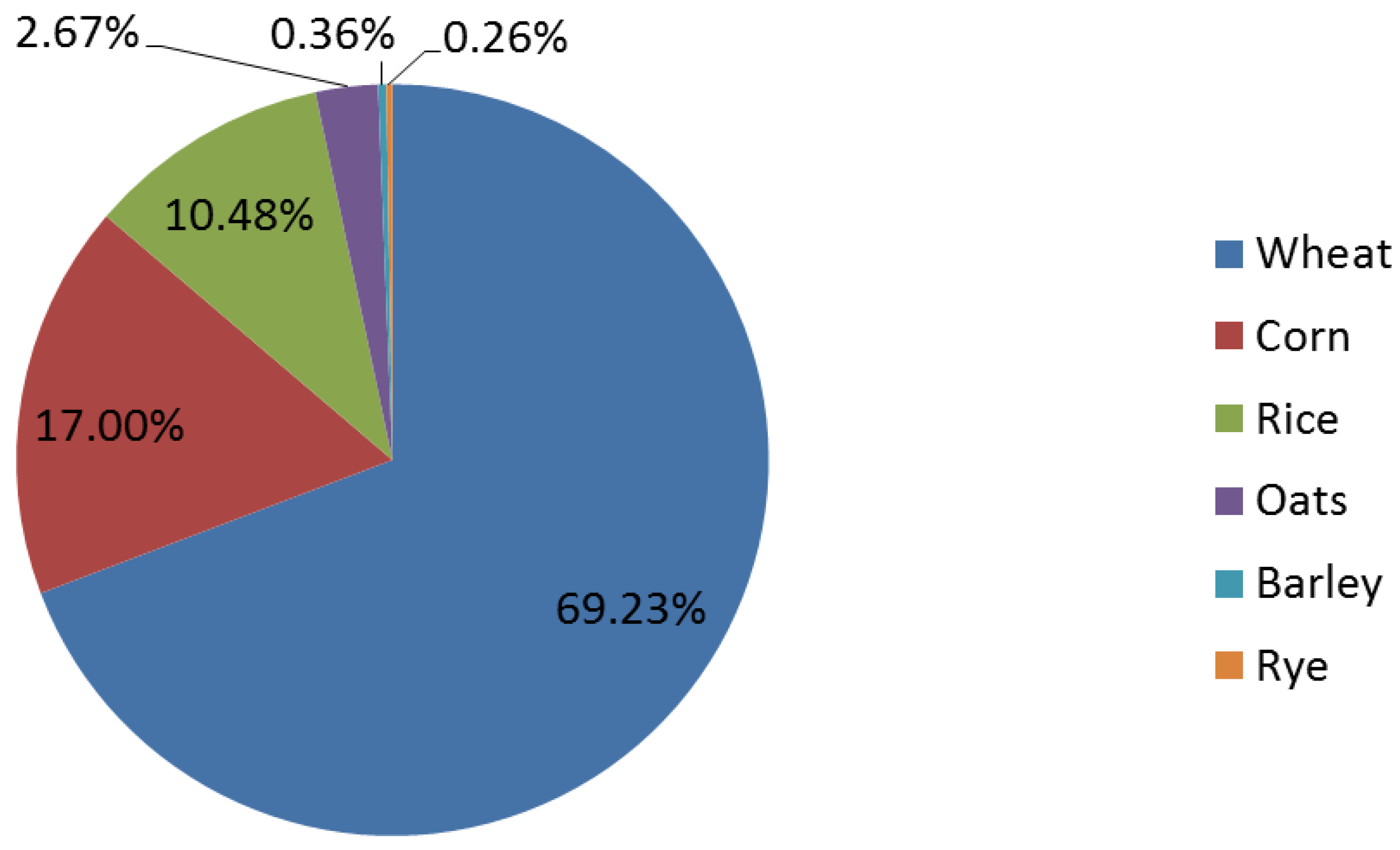
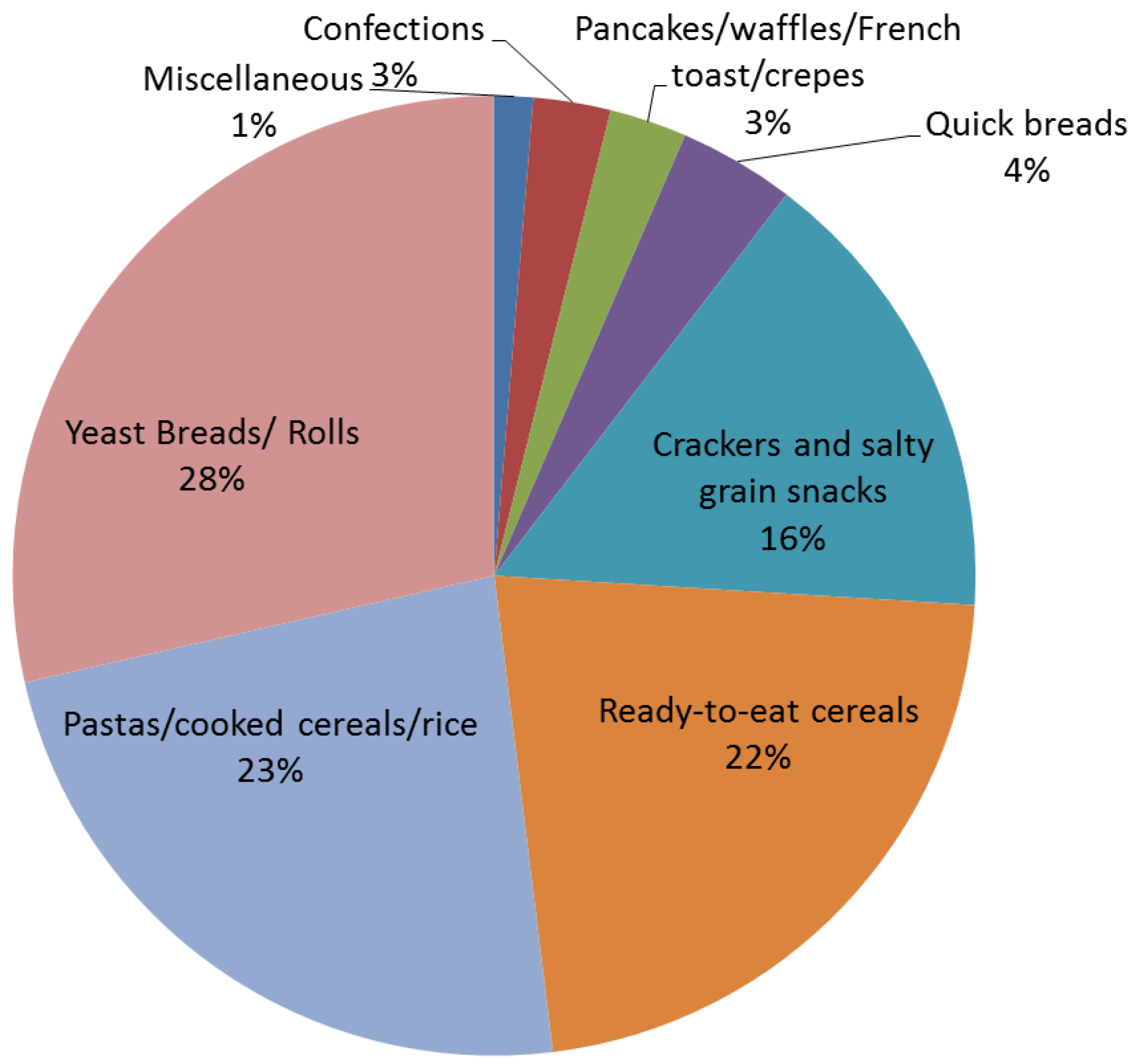
5. A Primer on Gastrointestinal Microbiota
6. Link between Whole Grains, Gastrointestinal Microbiota and Satiety
6.1. Bioactive Components of Whole Grains
6.2. Short Chain Fatty Acids from the Breakdown of Dietary Fibers
6.2.1. Butyrate
6.2.2. Propionate
6.2.3. Acetate
6.3. Bile Acids
7. Whole Grain Consumption and Changes in Gastrointestinal Microbiota
7.1. Microbiota Studies: Wheat

{kind=link}
{kind=link}
| Grain | Citation | Study Design | Subjects | Method | Results |
|---|---|---|---|---|---|
| Wheat | Costabile et al., 2008In text citation: [73] | Randomized crossover design 2 arms:
| 16 females, 15 males Age (years) 20–42 Mean: 25 BMI (kg/m2) 20–30 | FISH targeting: Atopobium group, Bifidobacterium spp., Eubacterium rectale group, Clostridium histolyticum group and Lactobacillus/Enterococcus |
|
| Wheat | Christensen et al., 2013 In text citation: [74] | Open label parallel intervention
| 72 post-menopausal females Age (years) 45–70 BMI (kg/m2) 27–37 | Quantitative PCR targeting: Bacteroidetes, Firmicutes, Bacteroides spp., Prevotella spp., Lactobacillus spp., Enterobacteriaceae, Bifidobacterium spp.:
|
|
| Corn | Carvalho-Wells et al., 2010 In text citation: [75] | Randomized crossover design 2 arms:
| 21 females, 11 males Age (years) 32 ± 8 BMI (kg/m2) 23.3 ± 0.6 | FISH targeting: Bacteroides spp., Bifidobacterium spp., Clostridium histolyticum/perfringens subgroup, Lactobacillus-Enterococcus subgroup and total bacteria. |
|
| Barley and Rice | Martinez et al., 2013 In text citation: [76] | Randomized crossover design 3 arms:
| 17 females, 11 males Age (years) 25.9 ± 5.5 BMI (kg/m2) 25.1 ± 4.5 | Pyrosequencing |
|
| Mixed Whole Grains | Ampatzolou et al., 2015 In text citation: [77] | Randomized crossover design 2 arms:
6 weeks/arm | 21 females, 12 males Age (years) 40–65 Mean:48.8 BMI (kg/m2) 20–35 | FISH targeting: Clostridium coccoides/Eubacterium rectale group, Bifidobacterium genus, Lactobacillus-Enterococcus group, Lactobacillus-Enterococcus group, Bacteroides-Prevotella group, Clostridium histolyticum group and Escherichia coli |
|
| Mixed Whole Grains | Ross et al., 2011 In text citation: [78] | Randomized crossover design 2 arms:
| 11 females, 6 males Age (years) 34.1 ± 3 BMI (kg/m2) 23.1 ± 0.8 | Quantitative PCR targeting: total bacteria, Bacteroides, Bifidobacterium, Clostridium coccoides, Clostridium leptum, Enterobacteria, Enterococcus spp. and Lactobacillus |
|
7.2. Microbiota Studies: Corn
7.3. Microbiota Studies: Barley and Rice
7.4. Microbiota Studies: Mixed Whole Grains
7.5. Microbiota Studies: In Vitro Studies
7.6. Concluding Remarks on Whole Grains and Microbiota
8. Whole Grains and Satiety
| Grain Type | Citation | Study Design | Subjects | Outcomes | Results |
|---|---|---|---|---|---|
| Whole grain wheat compared to refined grain wheat | Kristensen et al., 2010 In text citation: [89] | Randomized crossover design, 4 arms:
| 10 females, 6 males Age (years) 24.1 ± 3.8 BMI (kg/m2) 21.7 ± 2.2 |
| Whole grain bread: tended to increase fullness (p = 0.096) and satiety (p = 0.078) compared to the refined grain bread
|
| Whole grain wheat compared to refined grain wheat | Bodinham et al., 2011 In text citation: [90] | Balanced randomized crossover design 2 arms:
2 test rolls for 3 weeks culminating in a test day; no rolls were consumed on the test days | 9 females, 5 males Age (years) 26 ± 1.4 BMI (kg/m2) 21.8 ±0.8 |
|
|
| Whole grain oats compared to milled oat cereal | Rebello et al., 2013 In text citation: [91] | Randomized crossover design, 2 arms:
| 29 females 17 males Age range: 18–75 years (mean: 34.1 ± 14) BMI range: 17.9–60.1 kg/m2 (mean: 26.1 ± 7.2) |
| Oatmeal: reduced hunger (p = 0.0009), desire to eat (p = 0.0002) and prospective intake (p = 0.0012) and increased fullness (p = 0.005) compared to milled oat cereal |
| Whole grain oats compared to milled oat cereal | Rebello et al., 2014 In text citation: [84] | Randomized crossover design 3 arms:
| 28 females, 20 males Age (years) 29.8 ±9.9 BMI range: 18.5- 29.9 kg/m2(mean 27.1 ± 6.7) |
| Rolled oats: reduced the ratings of prospective intake more than milled oat cereal (p < 0.05) Instant oats: increased fullness more than rolled oats at 60 min (p < 0.05) increased fullness (p = 0.04), decreased desire to eat (p = 0.01), reduced prospective intake (p < 0.01) more than milled oat cereal over the course of four hours Rolled oats reduced prospective intake (p = 0.04) compared to milled oat cereal |
| Whole grain barley compared to refined grain wheat | Johansson et al., 2013 In text citation: [92] | Randomized crossover design with 2 arms:
evening before test day; the barley kernels were given in a 96.8-g portion; the refined wheat bread was given in a 119.7-g portion | 13 females, 6 males Age (years) 24.2 ±1.9 BMI (kg/m2) 22.3 ± 2.0 | The 5.5-hour test protocol included:
| Barley kernels relative to refined wheat bread: reduced perceived hunger levels following lunch (p < 0.05) less energy intake at ad libitum test lunch (p < 0.05) higher levels of breath hydrogen (p < 0.001) decreased incremental blood glucose AUC (p < 0.01) higher GLP-1 (p < 0.05) higher adiponectin (p < 0.05) lower serum fasting FFA (p < 0.05) |
| Whole grain rye compared to refined grain wheat | Isaksson et al., 2012 In text citation: [93] | Randomized crossover design 2 arms:
Rye porridge = 193 kcal Wheat bread = 185 kcal | 19 females, 5 males Age (years) 33 ± 13 BMI (kg/m2): 23.4 ± 2.2 | The 8-hour test day included:
| Rye porridge relative to refined wheat bread: subjects reported less hunger and desire to eat and higher satiety over the 4 hours after breakfast (all p-values ≤ 0.001); but no differences after the 4 hours higher levels of breath hydrogen between 4 and 8 hours post-test breakfast (p ≤ 0.05)No difference in food diaries |
| Rye (various forms) compared to wheat (various forms) | Rosen, Ostman and Bjorck, 2011 In text citation: [94] | Randomized crossover design, 7 arms:
| 5 females, 5 males Age (years) 26.0 ± 1.1 BMI (kg/m2) 22.6 ± 0.4 | The 7-hour test day included:
| Rye kernels: Reduced desire to eat compared to all other meal types Led to less energy intake at ad libitum lunch compared to refined wheat bread Rye kernels and whole grain rye bread: Induced higher breath hydrogen levels in the late post-breakfast period compared to the refined wheat bread and the refined rye bread (p ≤ 0.05) Whole wheat kernels: reduced serum FFA compared to refined wheat bread |
| Milled Whole Grain rye and whole rye kernels compared to refined grain wheat | Ibrugger et al., 2014 In text citation: [81] | Randomized crossover study 3 arms:
Rye kernels portion = 147 g Refined wheat bread portion = 111 g | 12 males Age (years) 25.6 ± 3.9 BMI (kg/m2) 23.1 ± 1.2 | Measurements included:
| No differences in VAS satiety between grain treatments Whole grain rye bread and rye kernels: increased breath hydrogen (p < 0.01 and p < 0.05, respectively) Reduced energy intake at lunch (by 11% p < 0.01 and by 7% p < 0.05, respectively) compared to refined wheat bread |
| Whole grain corn at low and high levels compared to refined grain wheat | Luhovyy et al., 2014 In text citation: [95] | Randomized crossover design, 3 arms: High amylose maize flour was mixed with refined wheat flour
| 30 males Age (years) 22.9 ± 0.6 BMI (kg/m2) 22.6 ± 0.3 |
| No effects of maize flour at either level on VAS satiety assessment or energy intake at ad libitum test lunch between interventions |
| Whole grains wheat whole grain barley refined grain rice | Schroeder et al., 2009 In text citation: [96] | Randomized crossover study, single blind, 3 arms:
Wheat cereal portion = 220 kcalBarley cereal portion = 200 kcal Rice cereal portion = 210 kcal | 35 females, 12 males Age Range: 19–58 years (mean: 31 ± 11)B MI Range: 18.8–30.7 kg/m2 (mean: 23 ± 3) |
| No difference in self-reported satiety scores between treatments Barley cereal: Overall VAS/SLIM scores for hunger were lower before lunch compared to before breakfast (p = 0.002)No differences between treatments in energy intake at ad libitum lunch |
8.1. Satiety Studies: Comparison of Whole versus Refined Wheat
8.2. Satiety Studies: Comparison of Oats with Different Levels of Processing
8.3. Satiety Studies: Comparison of Various Rye Products with Reference to Refined Wheat
8.4. Satiety Studies: Comparison of Whole Maize versus Refined Wheat
8.5. Satiety Studies: Comparison of Barley Kernels to a Reference Condition of Refined Wheat
8.6. Satiety Studies, Comparison of Cereals: Whole Grain Wheat, Whole Grain Barley and Refined Rice
9. Conclusions and Considerations for Future Studies
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferruzzi, M.G.; Jonnalagadda, S.S.; Liu, S.M.; Marquart, L.; McKeown, N.; Reicks, M.; Riccardi, G.; Seal, C.; Slavin, J.; Thielecke, F.; et al. Developing a standard definition of whole-grain foods for dietary recommendations: Summary. Report of a multidisciplinary expert roundtable discussion. Adv. Nutr. 2014, 5, 164–176. [Google Scholar] [CrossRef] [PubMed]
- OʼNeil, C.E.; Zanovec, M.; Cho, S.S.; Nicklas, T.A. Whole grain and fiber consumption are associated with lower body weight measures in us adults: National health and nutrition examination survey 1999–2004. Nutr. Res. 2010, 30, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P. Whole Grain Wheat Intake—Impact on Weight Loss, Body Composition and Cardiometabolic Factors—Results from Intervention Studies. In Processing of IUNS 20th International Congress of Nutrition, Granada, Spain, 15–20 September 2013.
- Rebello, C.J.; Greenway, F.L.; Finley, J.W. Whole grains and pulses: A comparison of the nutritional and health benefits. J. Agric. Food Chem. 2014, 62, 7029–7049. [Google Scholar] [CrossRef] [PubMed]
- Borneo, R.; Leon, A.E. Whole grain cereals: Functional components and health benefits. Food Funct. 2012, 3, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.A.M.; Dimberg, L.; Aman, P.; Landberg, R. Recent findings on certain bioactive components in whole grain wheat and rye. J. Cereal Sci. 2014, 59, 294–311. [Google Scholar] [CrossRef]
- Sanz-Penella, M.J.; Haros, M. Wheat and Rice in Disease Prevention and Health; Academic Press: Waltham, MA, USA, 2014; Chapter 2; pp. 17–31. [Google Scholar]
- Ndolo, V.U.; Beta, T. Comparative studies on composition and distribution of phenolic acids in cereal grain botanical fractions. Cereal Chem. 2014, 91, 522–530. [Google Scholar] [CrossRef]
- Ortiz-Robledo, F.; Villanueva-Fierro, I.; Oomah, B.D.; Lares-Asef, I.; Proal-Najera, J.B.; Navar-Chaidez, J.J. Avenanthramides and nutritional components of four mexican oat (avena sativa l.) varieties. Agrociencia 2013, 47, 225–232. [Google Scholar]
- Bjorck, I.; Ostman, E.; Kristensen, M.; Anson, N.M.; Price, R.K.; Haenen, G.R.M.M.; Havenaar, R.; Knudsen, K.E.B.; Frid, A.; Mykkanen, H.; et al. Cereal grains for nutrition and health benefits: Overview of results from in vitro, animal and human studies in the healthgrain project. Trends Food Sci. Technol. 2012, 25, 87–100. [Google Scholar] [CrossRef]
- Okarter, N.; Liu, R.H. Health benefits of whole grain phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Pol, K.; Christensen, R.; Bartels, E.M.; Raben, A.; Tetens, I.; Kristensen, M. Whole grain and body weight changes in apparently healthy adults: A systematic review and meta-analysis of randomized controlled studies. Am. J. Clin. Nutr. 2013, 98, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.H.; West, S.G.; Vanden Heuvel, J.P.; Jonnalagadda, S.S.; Ross, A.B.; Hill, A.M.; Grieger, J.A.; Lemieux, S.K.; Kris-Etherton, P.M. Effects of whole and refined grains in a weight-loss diet on markers of metabolic syndrome in individuals with increased waist circumference: A randomized controlled-feeding trial. Am. J. Clin. Nutr. 2014, 100, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Thielecke, F.; Jonnalagadda, S.S. Can whole grain help in weight management? J. Clin. Gastroenterol. 2014, 48, S70–S77. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; de Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Higo, A.; Ibe, N.; Kurimoto, Y.; Ootubo, S. The physical properties of 12 cereal flours including millets and whole grain (in Japanese). J. Jpn. Soc. Food Sci. Technol. 2014, 61, 117–126. [Google Scholar] [CrossRef]
- American Association of Cereal Chemists International. Whole grain definition. Cereal Foods World 1999, 45, 79. [Google Scholar]
- Van der Kamp, J.W.; Poutanen, K.; Seal, C.J.; Richardson, D.P. The HEALTHGRAIN definition of “whole grain”. Food Nutr. Res. 2014, 58. [Google Scholar] [CrossRef] [PubMed]
- Food Availability (Per Capita) Data System. Available online: http://ers.usda.gov/data-products/food-availability-%28per-capita%29-data-system.aspx (accessed on 20 April 2015).
- McGill, C.R.; Fulgoni, V.L.; Devareddy, L. Ten-year trends in fiber and whole grain intakes and food sources for the United States population: National health and nutrition examination survey 2001–2010. Nutrients 2015, 7, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Turroni, F.; Canchaya, C.; Vaughan, E.E.; OʼToole, P.W.; van Sinderen, D. Microbial diversity in the human intestine and novel insights from metagenomics. Front. Biosci. 2009, 14, 3214–3221. [Google Scholar] [CrossRef]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Kellow, N.J.; Coughlan, M.T.; Reid, C.M. Metabolic benefits of dietary prebiotics in human subjects: A systematic review of randomised controlled trials. Br. J. Nutr. 2014, 111, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Neyrinck, A.M.; Backhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Endocrinol. 2011, 7, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Munoz-Tamayo, R.; Paslier, D.L.E.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 11, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Tuohy, K.M.; Conterno, L.; Gasperotti, M.; Viola, R. Up-regulating the human intestinal microbiome using whole plant foods, polyphenols, and/or fiber. J. Agric. Food Chem. 2012, 60, 8776–8782. [Google Scholar] [CrossRef] [PubMed]
- Schloissnig, S.; Arumugam, M.; Sunagawa, S.; Mitreva, M.; Tap, J.; Zhu, A.; Waller, A.; Mende, D.R.; Kultima, J.R.; Martin, J.; et al. Genomic variation landscape of the human gut microbiome. Nature 2013, 493, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Duncan, S.H.; Louis, P.; Flint, H.J. Phylogeny, culturing, and metagenomics of the human gut microbiota. Trends Microbiol. 2014, 22, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Hooda, S.; Boler, B.M.V.; Serao, M.C.R.; Staeger, M.A.; Boileau, T.W.; Dowd, S.E.; Fahey, G.C., Jr.; Swanson, K.S. 454 pyrosequencing reveals a beneficial shift in fecal microbiota of healthy adult men consuming polydextrose or soluble corn fiber. Faseb J. 2012, 142, 1259–1265. [Google Scholar]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.A.; Walker, A.W.; Berry, S.H.; Duncan, S.H.; Farquarson, F.M.; Louis, P.; Thomson, J.M.; Satsangi, J.; Flint, H.J.; Parkhill, J.; et al. The impact of different DNA extraction kits and laboratories upon the assessment of human gut microbiota composition by 16s rRNA gene sequencing. PLoS ONE 2014, 9, e9. [Google Scholar] [CrossRef] [PubMed]
- Hartvigsen, M.L.; Laerke, H.N.; Overgaard, A.; Holst, J.J.; Knudsen, K.E.B.; Hermansen, K. Postprandial effects of test meals including concentrated arabinoxylan and whole grain rye in subjects with the metabolic syndrome: A randomised study. Eur. J. Clin. Nutr. 2014, 68, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.K.; Cao, X.H.; Zhou, J.Y.H. Effect of resistant starch structure on short-chain fatty acids production by human gut microbiota fermentation in vitro. Starch-Starke 2013, 65, 509–516. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. Fems Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; McCartney, A.L. What do we mean when we refer to bacteroidetes populations in the human gastrointestinal microbiota? Fems Microbiol. Lett. 2009, 299, 175–183. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Schippa, S.; Conte, M.P. Dysbiotic events in gut microbiota: Impact on human health. Nutrients 2014, 6, 5786–5805. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.D.; Sokol, H.; Dore, J.; Corthier, G.; Furet, J.P. The firmicutes/bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Raoult, D. Use of pyrosequencing and DNA barcodes to monitor variations in firmicutes and bacteroidetes communities in the gut microbiota of obese humans. BMC Genomics 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Caporaso, J.G.; Hooda, S.; Brulc, J.M.; Fahey, G.C.; Swanson, K.S. Fiber supplementation influences phylogenetic structure and functional capacity of the human intestinal microbiome: Follow-up of a randomized controlled trial. Am. J. Clin. Nutr. 2015, 101, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Bunesova, V.; Vlkova, E.; Rada, V.; Killer, J.; Musilova, S. Bifidobacteria from the gastrointestinal tract of animals: Differences and similarities. Benef. Microbes 2014, 5, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Mirande, C.; Kadlecikova, E.; Matulova, M.; Capek, P.; Bernalier-Donadille, A.; Forano, E.; Bera-Maillet, C. Dietary fibre degradation and fermentation by two xylanolytic bacteria bacteroides xylanisolvens xb1at and roseburia intestinalis xb6b4 from the human intestine. J. Appl. Microbiol. 2010, 109, 451–460. [Google Scholar] [PubMed]
- Xiong, W.L.; Giannone, R.J.; Morowitz, M.J.; Banfield, J.F.; Hettich, R.L. Development of an enhanced metaproteomic approach for deepening the microbiome characterization of the human infant gut. J. Proteome Res. 2015, 14, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Umu, O.C.O.; Oostindjer, M.; Pope, P.B.; Svihus, B.; Egelandsdal, B.; Nes, I.F.; Diep, D.B. Potential applications of gut microbiota to control human physiology. Antonie Van Leeuwenhoek 2013, 104, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.J.; Slavin, J.L. The effect of fiber on satiety and food intake: A systematic review. J. Am. Coll. Nutr. 2013, 32, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Bornhorst, G.M.; Stroebinger, N.; Rutherfurd, S.M.; Singh, R.P.; Moughan, P.J. Properties of gastric chyme from pigs fed cooked brown or white rice. Food Biophys. 2013, 8, 12–23. [Google Scholar] [CrossRef]
- Joyce, S.A.; Gahan, C.G.M. The gut microbiota and the metabolic health of the host. Curr. Opin. Gastroenterol. 2014, 30, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Sharma, V.; Elmen, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Estrela, S.; Whiteley, M.; Brown, S.P. The demographic determinants of human microbiome health. Trends Microbiol. 2015, 23, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Petschow, B.; Dore, J.; Hibberd, P.; Dinan, T.; Reid, G.; Blaser, M.; Cani, P.D.; Degnan, F.H.; Foster, J.; Gibson, G.; et al. Probiotics, prebiotics, and the host microbiome: The science of translation. Ann. N. Y. Acad. Sci. 2013, 1306, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Panickar, K.S. Effects of dietary polyphenols on neuroregulatory factors and pathways that mediate food intake and energy regulation in obesity. Mol. Nutr. Food Res. 2013, 57, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Cani, P.D. Gut microbiota and GLP-1. Rev. Endocr. Metab. Disord. 2014, 15, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.B. Concepts in functional foods: The case of inulin and oligofructose. J. Nutr. 1999, 129, S1398–S1401. [Google Scholar] [CrossRef]
- Geurts, L.; Neyrinck, A.M.; Delzenne, N.M.; Knauf, C.; Cani, P.D. Gut microbiota controls adipose tissue expansion, gut barrier and glucose metabolism: Novel insights into molecular targets and interventions using prebiotics. Benef. Microbes 2014, 5, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J. Obesity and the gut microbiota. J. Clin. Gastroenterol. 2011, 45, S128–S132. [Google Scholar] [CrossRef] [PubMed]
- Arora, T.; Sharma, R.; Frost, G. Propionate. Anti-obesity and satiety enhancing factor? Appetite 2011, 56, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.; Goszleth, G.; Frenyo, V.L. The major regulators of feed-intake: Ghrelin, leptin and their interactions. Literature review. Magy. Allatorv. Lapja 2012, 134, 504–512. [Google Scholar]
- Owyang, C.; Heldsinger, A. Vagal control of satiety and hormonal regulation of appetite. J. Neurogastroenterol. Motil. 2011, 17, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Sagen, E.K.; Varghese, Z.; MacDougall, K.; Preston, T. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adult. BMJ 2014. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Vella, A. Effects of GLP-1 on appetite and weight. Rev. Endoc. Metab. Disord. 2014, 15, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Tagliabue, A.; Elli, M. The role of gut microbiota in human obesity: Recent findings and future perspectives. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Giacco, R.; Della Pepa, G.; Luongo, D.; Riccardi, G. Whole grain intake in relation to body weight: From epidemiological evidence to clinical trials. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Niwano, Y.; Adachi, T.; Kashimura, J.; Sakata, T.; Sasaki, H.; Sekine, K.; Yamamoto, S.; Yonekubo, A.; Kimura, S. Is glycemic index of food a feasible predictor of appetite, hunger, and satiety? J. Nutr. Sci. Vitaminol. 2009, 55, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.E.B.; Laerke, H.N. Whole grain cereals and gut health. Agro Food Ind. Hi-Tech. 2008, 19, 6–8. [Google Scholar]
- McIntosh, G.H.; Noakes, M.; Royle, P.J.; Foster, P.R. Whole-grain rye and wheat foods and markers of bowel health in overweight middle-aged men. Am. J. Clin. Nutr. 2003, 77, 967–974. [Google Scholar] [PubMed]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Foglian, V.; Leonard, C.; Gibson, G.R.; Tuohy, K.M. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2008, 99, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.G.; Licht, T.R.; Kristensen, M.; Bahl, M.I. Bifidogenic effect of whole-grain wheat during a 12-week energy-restricted dietary intervention in postmenopausal women. Eur. J. Clin. Nutr. 2013, 67, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Wells, A.L.; Helmolz, K.; Nodet, C.; Molzer, C.; Leonard, C.; McKevith, B.; Thielecke, F.; Jackson, K.G.; Tuohy, K.M. Determination of the in vivo prebiotic potential of a maize-based whole grain breakfast cereal: A human feeding study. Br. J. Nutr. 2010, 104, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.Y.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. Isme J. 2013, 7, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Ampatzoglou, A.; Atwal, K.K.; Maidens, C.M.; Williams, C.L.; Ross, A.B.; Thielecke, F.; Jonnalagadda, S.S.; Kennedy, O.B.; Yaqoob, P. Increased whole grain consumption does not affect blood biochemistry, body composition, or gut microbiology in healthy, low-habitual whole grain consumers. J. Nutr. 2015, 145, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Bruce, S.J.; Blondel-Lubrano, A.; Oguey-Araymon, S.; Beaumont, M.; Bourgeois, A.; Nielsen-Moennoz, C.; Vigo, M.; Fay, L.B.; Kochhar, S.; et al. A whole-grain cereal-rich diet increases plasma betaine, and tends to decrease total and ldl-cholesterol compared with a refined-grain diet in healthy subjects. Br. J. Nutr. 2011, 105, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Martinez, I.; Rose, D.J. Holobiont nutrition: Considering the role of the gastrointestinal microbiota in the health benefits of whole grains. Gut Microb. 2013, 4, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.L.; Lovegrove, J.A.; Tuohy, K.M. In vitro evaluation of the microbiota modulation abilities of different sized whole oat grain flakes. Anaerobe 2010, 16, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Ibrugger, S.; Vigsnaes, L.K.; Blennow, A.; Skuflic, D.; Raben, A.; Lauritzen, L.; Kristensen, M. Second meal effect on appetite and fermentation of wholegrain rye foods. Appetite 2014, 80, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.L.; Lovegrove, J.A.; Tuohy, K.M. In vitro fermentation characteristics of whole grain wheat flakes and the effect of toasting on prebiotic potential. J. Med. Food 2012, 15, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.H.A.; Miller, J.C.B.; Petocz, P.; Farmakalidis, E. A satiety index of common foods. Eur. J. Clin. Nutr. 1995, 49, 675–690. [Google Scholar] [PubMed]
- Rebello, C.J.; Chu, Y.-F.; Johnson, W.D.; Martin, C.K.; Han, H.; Bordenave, N.; Shi, Y.; OʼShea, M.; Greenway, F.L. The role of meal viscosity and oat β-glucan characteristics in human appetite control: A randomized crossover trial. Nutr. J. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Begg, D.P.; Woods, S.C. The endocrinology of food intake. Nat. Rev. Endocrinol. 2013, 9, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Chapelot, D. Quantifying Satiation and Satiety. In Satiation, Satiety and the Control of Food Intake: Theory and Practice; Woodhead Publishing: Cambridge, UK, 2013; pp. 12–39. [Google Scholar]
- Blundell, J.E.; Goodson, S.; Halford, J.C.G. Regulation of appetite: Role of leptin in signalling systems for drive and satiety. Int. J. Obes. 2001, 25, S29–S34. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Jensen, M.G.; Riboldi, G.; Petronio, M.; Bugel, S.; Toubro, S.; Tetens, I.; Astrup, A. Wholegrain vs. Refined wheat bread and pasta. Effect on postprandial glycemia, appetite, and subsequent ad libitum energy intake in young healthy adults. Appetite 2010, 54, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Bodinham, C.L.; Hitchen, K.L.; Youngman, P.J.; Frost, G.S.; Robertson, M.D. Short-term effects of whole-grain wheat on appetite and food intake in healthy adults: A pilot study. Br. J. Nutr. 2011, 106, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Rebello, C.J.; Johnson, W.D.; Martin, C.K.; Xie, W.; O'Shea, M.; Kurilich, A.; Bordenave, N.; Andler, S.; van Klinken, B.J.W.; Chu, Y.-F.; et al. Acute effect of oatmeal on subjective measures of appetite and satiety compared to a ready-to-eat breakfast cereal: A randomized crossover trial. J. Am. Coll. Nutr. 2013, 32, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.V.; Nilsson, A.C.; Ostman, E.M.; Bjorck, I.M.E. Effects of indigestible carbohydrates in barley on glucose metabolism, appetite and voluntary food intake over 16 h in healthy adults. Nutr. J. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, H.; Tillander, I.; Andersson, R.; Olsson, J.; Fredriksson, H.; Webb, D.-L.; Aman, P. Whole grain rye breakfast—Sustained satiety during three weeks of regular consumption. Physiol. Behav. 2012, 105, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.A.; Ostman, E.M.; Bjorck, I.M.E. Effects of cereal breakfasts on postprandial glucose, appetite regulation and voluntary energy intake at a subsequent standardized lunch; focusing on rye products. Nutr. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Luhovyy, B.L.; Mollard, R.C.; Yurchenko, S.; Nunez, M.F.; Berengut, S.; Liu, T.T.; Smith, C.E.; Pelkman, C.L.; Anderson, G.H. The effects of whole grain high-amylose maize flour as a source of resistant starch on blood glucose, satiety, and food intake in young men. J. Food Sci. 2014, 79, H2550–H2556. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, N.; Gallaher, D.D.; Arndt, E.A.; Marquart, L. Influence of whole grain barley, whole grain wheat, and refined rice-based foods on short-term satiety and energy intake. Appetite 2009, 53, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Lange, E. Oats products as functional food. Zywnosc-Nauka Technol. Jakosc 2010, 17, 7–24. [Google Scholar] [CrossRef]
- Isaksson, H.; Rakha, A.; Andersson, R.; Fredriksson, H.; Olsson, J.; Aman, P. Rye kernel breakfast increases satiety in the afternoon—An effect of food structure. Nutr. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, H.; Sundberg, B.; Aman, P.; Fredriksson, H.; Olsson, J. Whole grain rye porridge breakfast improves satiety compared to refined wheat bread breakfast. Food Nutr. Res. 2008, 52. [Google Scholar] [CrossRef]
- Xu, Z.J.; Knight, R. Dietary effects on human gut microbiome diversity. Br. J. Nutr. 2015, 113, S1–S5. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Febbraio, M.A.; Whitham, M. From cytokine to myokine: The emerging role of interleukin-6 in metabolic regulation. Immunol. Cell Biol. 2014, 92, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, H.; Achremowicz, B.; Przygodzka, M. Antioxidants in cereal grains. Zywnosc-Nauka Technol. Jakosc 2012, 19, 5–26. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooper, D.N.; Martin, R.J.; Keim, N.L. Does Whole Grain Consumption Alter Gut Microbiota and Satiety? Healthcare 2015, 3, 364-392. https://doi.org/10.3390/healthcare3020364
Cooper DN, Martin RJ, Keim NL. Does Whole Grain Consumption Alter Gut Microbiota and Satiety? Healthcare. 2015; 3(2):364-392. https://doi.org/10.3390/healthcare3020364
Chicago/Turabian StyleCooper, Danielle N., Roy J. Martin, and Nancy L. Keim. 2015. "Does Whole Grain Consumption Alter Gut Microbiota and Satiety?" Healthcare 3, no. 2: 364-392. https://doi.org/10.3390/healthcare3020364
APA StyleCooper, D. N., Martin, R. J., & Keim, N. L. (2015). Does Whole Grain Consumption Alter Gut Microbiota and Satiety? Healthcare, 3(2), 364-392. https://doi.org/10.3390/healthcare3020364