
NADPH Oxidase Activity in Cerebral Arterioles Is a Key Mediator of Cerebral Small Vessel Disease—Implications for Prevention

{kind=link}
Abstract
:1. Functional Importance of Cerebral Small Vessel Disease in Brain Aging
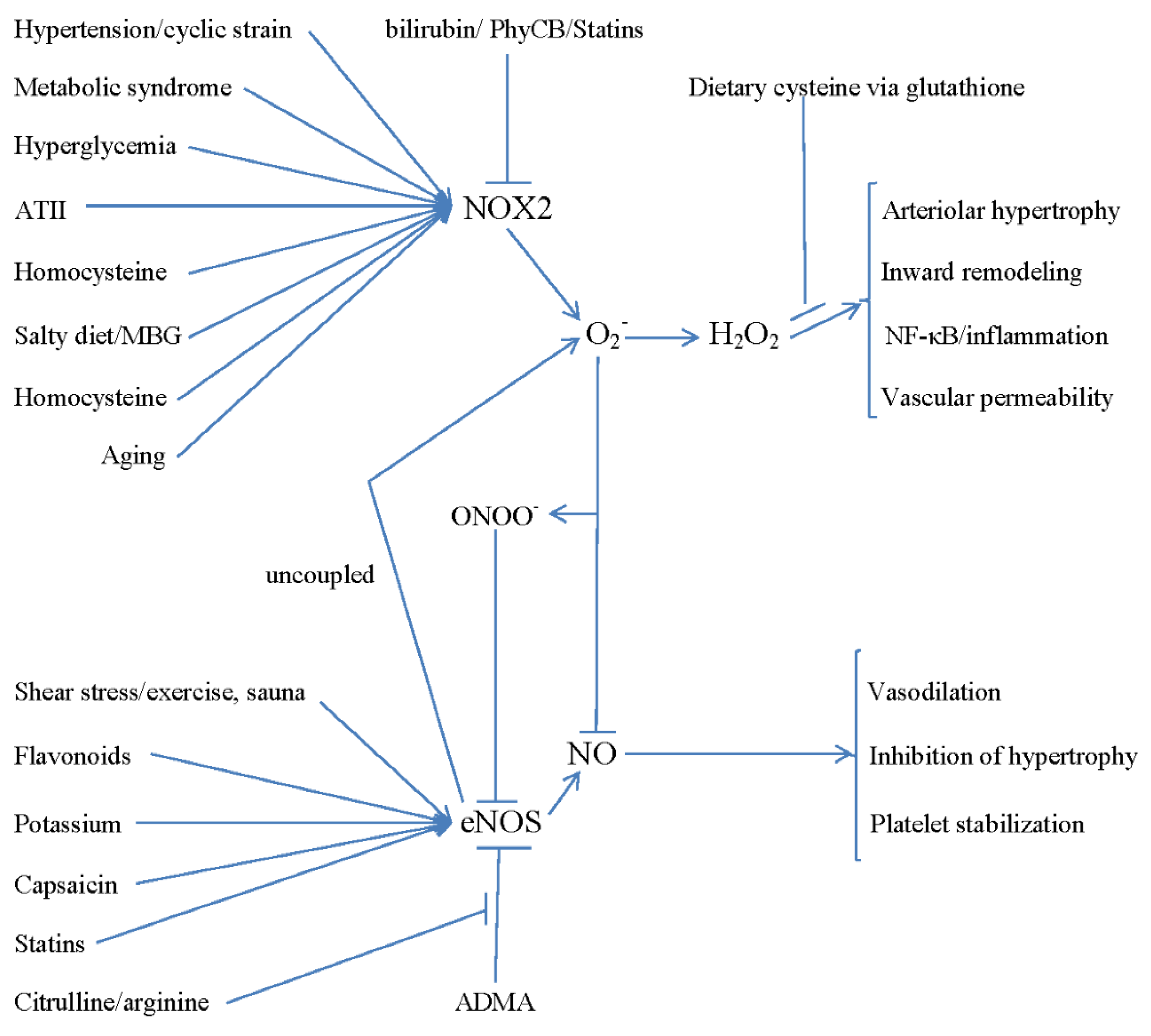
2. Activation of NAPDH Oxidase in Cerebral Arterioles Is Central to the Pathogenesis of SVD
3. What Factors Activate NADPH Oxidase in Cerebral Arterioles?
4. Practical Strategies for Preventing Cerebral Small Vessel Disease
5. Further Measures for Supporting eNOS Activity
6. The Impact of Dietary Protein
7. Conclusions
Acknowledgements
Conflicts of Interest
References
- Markus, H.S. Genes, endothelial function and cerebral small vessel disease in man. Exp. Physiol. 2008, 93, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Grueter, B.E.; Schulz, U.G. Age-related cerebral white matter disease (leukoaraiosis): A review. Postgrad. Med. J. 2012, 88, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Chutinet, A.; Rost, N.S. White matter disease as a biomarker for long-term cerebrovascular disease and dementia. Curr. Treat. Opt. Cardiovasc. Med. 2014. [Google Scholar] [CrossRef]
- Chrissobolis, S.; Faraci, F.M. The role of oxidative stress and NADPH oxidase in cerebrovascular disease. Trends Mol. Med. 2008, 14, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.A.; Drummond, G.R.; Schmidt, H.H.; Sobey, C.G. NADPH oxidase activity and function are profoundly greater in cerebral versus systemic arteries. Circ. Res. 2005, 97, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Ago, T.; Kitazono, T.; Kuroda, J.; Kumai, Y.; Kamouch, M.; Ooboshi, H.; Wakisaka, M.; Kawahara, T.; Rokutan, K.; Ibayashi, S.; et al. NAD(P)H oxidases in rat basilar arterial endothelial cells. Stroke 2005, 36, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.L.; Baumbach, G.L. Nox2 deficiency prevents hypertension-induced vascular dysfunction and hypertrophy in cerebral arterioles. Int. J. Hypertens. 2013. [Google Scholar] [CrossRef]
- Al Ghouleh, I.; Frazziano, G.; Rodriguez, A.I.; Csanyi, G.; Maniar, S.; St. Croix, C.M.; Kelley, E.E.; Egana, L.A.; Song, G.J.; Bisello, A.; et al. Aquaporin 1, Nox1, and Ask1 mediate oxidant-induced smooth muscle cell hypertrophy. Cardiovasc. Res. 2013, 97, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, H.; Lee, K.H.; Tsao, P.S. Cyclic strain induces reactive oxygen species production via an endothelial NAD(P)H oxidase. J. Cell Biochem. Suppl. 2001, S36, 99–106. [Google Scholar] [CrossRef]
- Chan, S.L.; Baumbach, G.L. Deficiency of Nox2 prevents angiotensin II-induced inward remodeling in cerebral arterioles. Front. Physiol. 2013. [Google Scholar] [CrossRef]
- Didion, S.P.; Faraci, F.M. Angiotensin II produces superoxide-mediated impairment of endothelial function in cerebral arterioles. Stroke 2003, 34, 2038–2042. [Google Scholar] [CrossRef] [PubMed]
- Kazama, K.; Anrather, J.; Zhou, P.; Girouard, H.; Frys, K.; Milner, T.A.; Iadecola, C. Angiotensin II impairs neurovascular coupling in neocortex through NADPH oxidase-derived radicals. Circ. Res. 2004, 95, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Mayhan, W.G.; Arrick, D.M.; Sharpe, G.M.; Patel, K.P.; Sun, H. Inhibition of NAD(P)H oxidase alleviates impaired NOS-dependent responses of pial arterioles in type 1 diabetes mellitus. Microcirculation 2006, 13, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zheng, H.; Molacek, E.; Fang, Q.; Patel, K.P.; Mayhan, W.G. Role of NAD(P)H oxidase in alcohol-induced impairment of endothelial nitric oxide synthase-dependent dilation of cerebral arterioles. Stroke 2006, 37, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Sun, H.; Arrick, D.M.; Mayhan, W.G. Inhibition of NADPH oxidase improves impaired reactivity of pial arterioles during chronic exposure to nicotine. J. Appl. Physiol. 2006, 100, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Iida, H.; Iida, M.; Takenaka, M.; Fukuoka, N.; Dohi, S. Rho-kinase inhibitor and nicotinamide adenine dinucleotide phosphate oxidase inhibitor prevent impairment of endothelium-dependent cerebral vasodilation by acute cigarette smoking in rats. J. Renin. Angiotensin. Aldosterone Syst. 2008, 9, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.M.; Kinzenbaw, D.A.; Chen, X.; Zhan, S.; Mezzeti, E.; Filosa, J.; Faulkner, J.L.; Faraci, F.M.; Didion, S.P. Nox2-derived superoxide contributes to cerebral vascular dysfunction in diet-induced obesity. Stroke 2013, 44, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
- Mayhan, W.G.; Arrick, D.M.; Sharpe, G.M.; Sun, H. Age-related alterations in reactivity of cerebral arterioles: Role of oxidative stress. Microcirculation 2008, 15, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Y.; Cao, Z.G.; Zhang, J.R.; Li, Y.; Wang, R.T. Decreased serum bilirubin is associated with silent cerebral infarction. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Shim, J.Y.; Lee, H.R.; Kang, H.T.; Lee, J.H.; Lee, Y.J. Association between serum total bilirubin level and leukoaraiosis in Korean adults. Clin. Biochem. 2012, 45, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Lanone, S.; Bloc, S.; Foresti, R.; Almolki, A.; Taille, C.; Callebert, J.; Conti, M.; Goven, D.; Aubier, M.; Dureuil, B.; et al. Bilirubin decreases nos2 expression via inhibition of NAD(P)H oxidase: Implications for protection against endotoxic shock in rats. FASEB J. 2005, 19, 1890–1892. [Google Scholar] [PubMed]
- Matsumoto, H.; Ishikawa, K.; Itabe, H.; Maruyama, Y. Carbon monoxide and bilirubin from heme oxygenase-1 suppresses reactive oxygen species generation and plasminogen activator inhibitor-1 induction. Mol. Cell Biochem. 2006, 291, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Roberts, S.J.; Datla, S.; Dusting, G.J. NO modulates NADPH oxidase function via heme oxygenase-1 in human endothelial cells. Hypertension 2006, 48, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Datla, S.R.; Dusting, G.J.; Mori, T.A.; Taylor, C.J.; Croft, K.D.; Jiang, F. Induction of heme oxygenase-1 in vivo suppresses NADPH oxidase derived oxidative stress. Hypertension 2007, 50, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.P.; Vitek, L.; Schwertner, H.A. Serum bilirubin and genes controlling bilirubin concentrations as biomarkers for cardiovascular disease. Clin. Chem. 2010, 56, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Vitek, L. The role of bilirubin in diabetes, metabolic syndrome, and cardiovascular diseases. Front. Pharmacol. 2012. [Google Scholar] [CrossRef]
- Zelenka, J.; Muchova, L.; Zelenkova, M.; Vanova, K.; Vreman, H.J.; Wong, R.J.; Vitek, L. Intracellular accumulation of bilirubin as a defense mechanism against increased oxidative stress. Biochimie 2012, 94, 1821–1827. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Serum bilirubin may serve as a marker for increased heme oxygenase activity and inducibility in tissues—A rationale for the versatile health protection associated with elevated plasma bilirubin. Med. Hypotheses. 2013, 81, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Inoguchi, T.; Li, P.; Umeda, F.; Kakimoto, M.; Imamura, M.; Aoki, T.; Hashimoto, T.; Naruse, M.; Sano, H.; Utsumi, H.; et al. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C—Dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 2000, 49, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Maloney, E.; Sweet, I.R.; Hockenbery, D.M.; Pham, M.; Rizzo, N.O.; Tateya, S.; Handa, P.; Schwartz, M.W.; Kim, F. Activation of NF-kappaB by palmitate in endothelial cells: A key role for NADPH oxidase-derived superoxide in response to TLR4 activation. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Edirimanne, V.E.; Woo, C.W.; Siow, Y.L.; Pierce, G.N.; Xie, J.Y. Homocysteine stimulates NADPH oxidase-mediated superoxide production leading to endothelial dysfunction in rats. Can. J. Physiol. Pharmacol. 2007, 85, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.M.; Wu, C.F.; Lu, G.P. Atorvastatin attenuates homocysteine-induced apoptosis in human umbilical vein endothelial cells via inhibiting NADPH oxidase-related oxidative stress-triggered p38MAPK signaling. Acta Pharmacol. Sin. 2009, 30, 1392–1398. [Google Scholar] [PubMed]
- Omae, T.; Nagaoka, T.; Tanano, I.; Yoshida, A. Homocysteine inhibition of endothelium-dependent nitric oxide-mediated dilation of porcine retinal arterioles via enhanced superoxide production. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2288–2295. [Google Scholar] [CrossRef]
- Jaimes, E.A.; DeMaster, E.G.; Tian, R.X.; Raij, L. Stable compounds of cigarette smoke induce endothelial superoxide anion production via NADPH oxidase activation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Orosz, Z.; Csiszar, A.; Labinskyy, N.; Smith, K.; Kaminski, P.M.; Fedinandy, P.; Wolin, M.S.; Rivera, A.; Ungvari, Z. Cigarette smoke-induced proinflammatory alterations in the endothelial phenotype: Role of NAD(P)H oxidase activation. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H130–H139. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Kaneda, Y.; Ogihara, T.; Morishita, R. Endothelial dysfunction in hyperglycemia as a trigger of atherosclerosis. Curr. Diabetes Rev. 2005, 1, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.P.; di Marco, E.; Okabe, J.; Szydralewiez, C.; Heitz, F.; Montezano, A.C.; de Haan, J.B.; Koulis, C.; El-Osta, A.; Andrews, K.L.; et al. NADPH oxidase 1 plays a key role in diabetes mellitus-accelerated atherosclerosis. Circulation 2013, 127, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Bayraktutan, U. Hyperglycaemia promotes cerebral barrier dysfunction through activation of protein kinase C-beta. Diabetes Obes. Metab. 2013, 15, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, O.V.; Talan, M.I.; Agalakova, N.I.; Lakatta, E.G.; Bagrov, A.Y. Endogenous ligand of alpha(1) sodium pump, marinobufagenin, is a novel mediator of sodium chloride—Dependent hypertension. Circulation 2002, 105, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, K.L.; Fedorova, O.V.; Racine, M.L.; Geolfos, P.E.; Chonchol, M.; Fleenor, B.S.; Lakatta, E.G.; Bagrov, A.Y.; Seals, D.R. Dietary sodium restriction and association with urinary marinobufagenin, blood pressure, and aortic stiffness. Clin. J. Am. Soc. Nephrol. 2013, 8, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Marinobufagenin and cyclic strain may activate endothelial NADPH oxidase, contributing to the adverse impact of salty diets on vascular and cerebral health. Med. Hypotheses. 2012, 78, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rusch, N.J.; Lombard, J.H. Loss of endothelium and receptor-mediated dilation in pial arterioles of rats fed a short-term high salt diet. Hypertension 1999, 33, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Kuller, L.; Reisler, D.M. An explanation for variations in distribution of stroke and arteriosclerotic heart disease among populations and racial groups. Am. J. Epidemiol. 1971, 93, 1–9. [Google Scholar] [PubMed]
- Vartiainen, E.; Du, D.J.; Marks, J.S.; Korhonen, H.; Geng, G.Y.; Guo, Z.Y.; Koplan, J.P.; Pietinen, P.; We, G.L.; Williamson, D.; et al. Mortality, cardiovascular risk factors, and diet in China, Finland, and the United States. Public Health Rep. 1991, 106, 41–46. [Google Scholar] [PubMed]
- He, J.; Klag, M.J.; Wu, Z.; Whelton, P.K. Stroke in the People’s Republic of China. II. Meta-analysis of hypertension and risk of stroke. Stroke 1995, 26, 2228–2232. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.; Jacobs, D.R., Jr.; Hayashi, T.; Konishi, M.; Nelson, J.; Iso, H.; Strong, J. A comparison of lesions in small intracerebral arteries among Japanese men in Hawaii and Japan. Stroke 1994, 25, 60–65. [Google Scholar] [CrossRef]
- Topakian, R.; Barrick, T.R.; Howe, F.A.; Markus, H.S. Blood-brain barrier permeability is increased in normal-appearing white matter in patients with lacunar stroke and leucoaraiosis. J. Neurol. Neurosurg. Psychiatry 2010, 81, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Uh, J.; Yezhuvath, U.; Cheng, Y.; Lu, H. In vivo vascular hallmarks of diffuse leukoaraiosis. J. Magn. Reson. Imaging 2010, 32, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Gasparovic, C.; Huisa, B.N.; Adair, J.C.; Edmonds, E.; Prestopnik, J.; Grossetete, M.; Shah, N.J.; Wills, J.; Qualls, C.; et al. Blood-brain barrier permeability abnormalities in vascular cognitive impairment. Stroke 2011, 42, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Krizbai, I.A.; Bauer, H.; Bresgen, N.; Eckl, P.M.; Farkas, A.; Szatmari, E.; Traweger, A.; Wejksza, K.; Bauer, H.C. Effect of oxidative stress on the junctional proteins of cultured cerebral endothelial cells. Cell Mol. Neurobiol. 2005, 25, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gong, B.; Yang, Y.; Awasthi, Y.C.; Woods, M.; Boor, P.J. Glutathione-S-transferase protects against oxidative injury of endothelial cell tight junctions. Endothelium 2007, 14, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.H.; Potula, R.; Fan, S.; Eidem, T.; Papugani, A.; Reichenbach, N.; Dykstra, H.; Weksler, B.B.; Romero, I.A.; Couraud, P.O.; et al. Methamphetamine disrupts blood-brain barrier function by induction of oxidative stress in brain endothelial cells. J. Cereb. Blood Flow Metab. 2009, 29, 1933–1945. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wu, J.; Zou, M.H. Activation of AMP-activated protein kinase alleviates high-glucose-induced dysfunction of brain microvascular endothelial cell tight-junction dynamics. Free Radic. Biol. Med. 2012, 53, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Zehendner, C.M.; Librizzi, L.; Hedrich, J.; Bauer, N.M.; Angamo, E.A.; de Curtis, M.; Luhmann, H.J. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS One 2013, 8, e82823. [Google Scholar] [CrossRef] [PubMed]
- Ing, N.H.; Berghman, L.; Abi-Ghanem, D.; Abbas, K.; Kaushik, A.; Riggs, P.K.; Puschett, J.B. Marinobufagenin regulates permeability and gene expression of brain endothelial cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R918–R924. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.H.; Kohler, T.; Ruckschloss, U.; Just, I.; Hecker, M. Improvement of nitric oxide-dependent vasodilatation by HMG-CoA reductase inhibitors through attenuation of endothelial superoxide anion formation. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.; Laufs, U. Effects of statins on endothelium and signaling mechanisms. Stroke 2004, 35, 2708–2711. [Google Scholar] [CrossRef] [PubMed]
- Ebinger, M.; Brunecker, P.; Schultze-Amberger, J.; Gertz, K.; Muller, B.; Fiebach, J.B.; Dichganz, M.; Endres, M. Statins and cerebral perfusion in patients with leukoaraiosis—A translational proof-of-principal MRI study. Int. J. Stroke 2012. [Google Scholar] [CrossRef]
- Terry, M.J.; Maines, M.D.; Lagarias, J.C. Inactivation of phytochrome- and phycobiliprotein-chromophore precursors by rat liver biliverdin reductase. J. Biol. Chem. 1993, 268, 26099–26106. [Google Scholar] [PubMed]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from Spirulina platensis protect against diabetic nephropathy by inhibiting oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R110–R120. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Clinical potential of Spirulina as a source of phycocyanobilin. J. Med. Food 2007, 10, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Marin-Prida, J.; Pavon-Fuentes, N.; Llopiz-Arzuaga, A.; Fernandez-Masso, J.R.; Delgado-Roche, L.; Mendoza-Mari, Y.; Santana, S.P.; Cruz-Ramirez, A.; Valenzuela-Silva, C.; Nazabol-Galvez, M.; et al. Phycocyanobilin promotes PC12 cell survival and modulates immune and inflammatory genes and oxidative stress markers in acute cerebral hypoperfusion in rats. Toxicol. Appl. Pharmacol. 2013, 272, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Penton-Rol, G.; Marin-Prida, J.; Pardo-Andreu, G.; Martinez-Sanchez, G.; Acosta-Medina, E.F.; Valdivia-Acosta, A.; Lagumersindez-Denis, N.; Rodriguez-Jimenez, E.; Llopiz-Arzuaga, A.; Lopez-Saura, P.A.; et al. C-Phycocyanin is neuroprotective against global cerebral ischemia/reperfusion injury in gerbils. Brain Res. Bull. 2011, 86, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Thaakur, S.; Sravanthi, R. Neuroprotective effect of Spirulina in cerebral ischemia-reperfusion injury in rats. J. Neural. Transm. 2010, 117, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chang, C.F.; Chou, J.; Chen, H.L.; Deng, H.; Harvey, B.K.; Cadet, J.L.; Bickford, P. Dietary supplementation with blueberries, spinach, or spirulina reduces ischemic brain damage. Exp. Neurol. 2005, 193, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kahles, T.; Brandes, R.P. Which NADPH oxidase isoform is relevant for ischemic stroke? The case for Nox 2. Antioxid. Redox Signal. 2013, 18, 1400–1417. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; Camillo, T.P.; Braunersreuther, V.; Mach, F.; Vuilleumier, N.; Montecucco, F. Pathophysiology and treatments of oxidative injury in ischemic stroke: Focus on the phagocytic NADPH oxidase (NOX) 2. Antioxid. Redox Signal. 2014. [Google Scholar] [CrossRef]
- Miller, A.A.; Dusting, G.J.; Roulston, C.L.; Sobey, C.G. NADPH-oxidase activity is elevated in penumbral and non-ischemic cerebral arteries following stroke. Brain Res. 2006, 1111, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Tobian, L. High-potassium diets markedly protect against stroke deaths and kidney disease in hypertensive rats, an echo from prehistoric days. J. Hypertens. Suppl. 1986, 4, S67–S76. [Google Scholar] [PubMed]
- Khaw, K.T.; Barrett-Connor, E. Dietary potassium and stroke-associated mortality. A 12-year prospective population study. N. Engl. J. Med. 1987, 316, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Virtamo, J.; Wolk, A. Potassium, calcium, and magnesium intakes and risk of stroke in women. Am. J. Epidemiol. 2011, 174, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Aaron, K.J.; Sanders, P.W. Role of dietary salt and potassium intake in cardiovascular health and disease: A review of the evidence. Mayo Clin. Proc. 2013, 88, 987–995. [Google Scholar] [CrossRef] [PubMed]
- McCabe, R.D.; Bakarich, M.A.; Srivastava, K.; Young, D.B. Potassium inhibits free radical formation. Hypertension 1994, 24, 77–82. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Endothelial membrane potential regulates production of both nitric oxide and superoxide—A fundamental determinant of vascular health. Med. Hypotheses. 1999, 53, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Sohn, H.Y.; Keller, M.; Gloe, T.; Morawietz, H.; Rueckschloss, U.; Pohl, U. The small G-protein Rac mediates depolarization-induced superoxide formation in human endothelial cells. J. Biol. Chem. 2000, 275, 18745–18750. [Google Scholar] [CrossRef] [PubMed]
- Lindeberg, S.; Lundh, B. Apparent absence of stroke and ischaemic heart disease in a traditional Melanesian island: A clinical study in Kitava. J. Intern. Med. 1993, 233, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Lindeberg, S. Apparent absence of cerebrovascular disease in Melanesians. Ph.D Thesis, Lund University, Lund, Sweden, 1994. [Google Scholar]
- McCarty, M.F. Up-regulation of endothelial nitric oxide activity as a central strategy for prevention of ischemic stroke—Just say NO to stroke! Med. Hypotheses. 2000, 55, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.T.; Lee, M.; Hong, K.S.; Ovbiagele, B.; Saver, J.L. Efficacy of folic acid supplementation in cardiovascular disease prevention: An updated meta-analysis of randomized controlled trials. Eur. J. Intern. Med. 2012, 23, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Tan, S.; Xu, Y.; Chandra, A.; Shi, C.; Song, B.; Qin, J.; Gao, Y. Vitamin B supplementation, homocysteine levels, and the risk of cerebrovascular disease: A meta-analysis. Neurology 2013, 81, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Hunt, B.J.; O’Sullivan, M.; Bell, R.; D’Souza, R.; Jeffery, S.; Bamford, J.M.; Markus, H.S. Homocysteine is a risk factor for cerebral small vessel disease, acting via endothelial dysfunction. Brain 2004, 127, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Khan, U.; Hassan, A.; Vallance, P.; Markus, H.S. Asymmetric dimethylarginine in cerebral small vessel disease. Stroke 2007, 38, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Calabro, R.S.; Gervasi, G.; Baglieri, A.; Furnari, A.; Marino, S.; Bramanti, P. Is high oral dose L-arginine intake effective in leukoaraiosis? Preliminary data, study protocol and expert’s opinion. Curr. Aging Sci. 2013, 6, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Schwedhelm, E.; Maas, R.; Freese, R.; Jung, D.; Lukacs, Z.; Jambrecina, A.; Spickler, W.; Schulze, F.; Boger, R.H. Pharmacokinetic and pharmacodynamic properties of oral L-citrulline and L-arginine: Impact on nitric oxide metabolism. Br. J. Clin. Pharmacol. 2008, 65, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, M.C.; Wever, R.M.; Kastelein, J.J.; Van, D.T.; Koomans, H.A.; Rabelink, T.J. 5-methyltetrahydrofolate, the active form of folic acid, restores endothelial function in familial hypercholesterolemia. Circulation 1998, 97, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Stroes, E.S.; van Faassen, E.E.; Yo, M.; Martasek, P.; Boer, P.; Govers, R.; Rabelink, T.J. Folic acid reverts dysfunction of endothelial nitric oxide synthase. Circ. Res. 2000, 86, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Hyndman, M.E.; Verma, S.; Rosenfeld, R.J.; Anderson, T.J.; Parsons, H.G. Interaction of 5-methyltetrahydrofolate and tetrahydrobiopterin on endothelial function. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H2167–H2172. [Google Scholar] [CrossRef] [PubMed]
- Antoniades, C.; Shirodaria, C.; Warrick, N.; Cai, S.; de Bono, J.; Lee, J.; Leeson, P.; Neubauer, S.; Ratnatunga, C.; Pillai, R.; et al. 5-methyltetrahydrofolate rapidly improves endothelial function and decreases superoxide production in human vessels: Effects on vascular tetrahydrobiopterin availability and endothelial nitric oxide synthase coupling. Circulation 2006, 114, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Rezk, B.M.; Haenen, G.R.; van der Vijgh, W.J.; Bast, A. Tetrahydrofolate and 5-methyltetrahydrofolate are folates with high antioxidant activity. Identification of the antioxidant pharmacophore. FEBS Lett. 2003, 555, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Pardridge, W.M. Blood-brain barrier transport of reduced folic acid. Pharm. Res. 1999, 16, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Sabharanjak, S.; Mayor, S. Folate receptor endocytosis and trafficking. Adv .Drug Deliv. Rev. 2004, 56, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- McAllister, R.M.; Newcomer, S.C.; Laughlin, M.H. Vascular nitric oxide: Effects of exercise training in animals. Appl. Physiol. Nutr. Metab. 2008, 33, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Mayhan, W.G.; Arrick, D.M.; Sun, H.; Patel, K.P. Exercise training restores impaired dilator responses of cerebral arterioles during chronic exposure to nicotine. J. Appl. Physiol. (1985) 2010, 109, 1109–1114. [Google Scholar] [CrossRef]
- Mayhan, W.G.; Arrick, D.M.; Patel, K.P.; Sun, H. Exercise training normalizes impaired NOS-dependent responses of cerebral arterioles in type 1 diabetic rats. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1013–H1020. [Google Scholar] [CrossRef] [PubMed]
- Arrick, D.M.; Yang, S.; Li, C.; Cananzi, S.; Mayhan, W.G. Vigorous exercise training improves reactivity of cerebral arterioles and reduces brain injury following transient focal ischemia. Microcirculation 2014, 21, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Jefferis, B.J.; Whincup, P.H.; Papacosta, O.; Wannamethee, S.G. Protective effect of time spent walking on risk of stroke in older men. Stroke 2014, 45, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, H.; Heiss, C.; Balzer, J.; Kleinbongard, P.; Keen, C.L.; Hollenberg, N.K.; Sies, H.; Kwik-Uribe, C.; Schmitz, H.H.; Kelm, M. (−)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Hodgson, J.M.; Proudfoot, J.M.; McKinley, A.J.; Puddey, I.B.; Croft, K.D. Pure dietary flavonoids quercetin and (−)-epicatechin augment nitric oxide products and reduce endothelin-1 acutely in healthy men. Am. J. Clin. Nutr. 2008, 88, 1018–1025. [Google Scholar] [PubMed]
- Ramirez-Sanchez, I.; Maya, L.; Ceballos, G.; Villarreal, F. (−)-Epicatechin induces calcium and translocation independent eNOS activation in arterial endothelial cells. Am. J. Physiol. Cell Physiol. 2011, 300, C880–C887. [Google Scholar] [CrossRef] [PubMed]
- Fisher, N.D.; Sorond, F.A.; Hollenberg, N.K. Cocoa flavanols and brain perfusion. J. Cardiovasc. Pharmacol. 2006, 47, S210–S214. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Bernatova, I.; Puzserova, A.; Balis, P.; Sestakova, N.; Pechanova, O.; Fraga, C.G. (−)‑Epicatechin reduces blood pressure and improves vasorelaxation in spontaneously hypertensive rats by NO-mediated mechanism. IUBMB Life 2013, 65, 710–715. [Google Scholar] [CrossRef]
- Osakabe, N.; Shibata, M. Ingestion of cocoa ameliorates endothelial dysfunction in mesentery arterioles induced by high fat diet in rats: An in vivo intravital microscopy study. Life Sci. 2012, 91, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Farouque, H.M.; Leung, M.; Hope, S.A.; Baldi, M.; Schechter, C.; Cameron, J.D.; Meredith, I.T. Acute and chronic effects of flavanol-rich cocoa on vascular function in subjects with coronary artery disease: A randomized double-blind placebo-controlled study. Clin. Sci. (Lond.) 2006, 111, 71–80. [Google Scholar] [CrossRef]
- Benito, S.; Lopez, D.; Saiz, M.P.; Buxaderas, S.; Sanchez, J.; Puig-Parellada, P.; Mitjavila, M.T. A flavonoid-rich diet increases nitric oxide production in rat aorta. Br. J. Pharmacol. 2002, 135, 910–916. [Google Scholar] [CrossRef]
- Monahan, K.D.; Feehan, R.P.; Kunselman, A.R.; Preston, A.G.; Miller, D.L.; Lott, M.E. Dose-dependent increases in flow-mediated dilation following acute cocoa ingestion in healthy older adults. J. Appl. Physiol. (1985) 2011, 111, 1568–1574. [Google Scholar] [CrossRef]
- Drouin, A.; Bolduc, V.; Thorin-Trescases, N.; Belanger, E.; Fernandez, P.; Baraghis, E.; Lesage, F.; Gillis, M.A.; Villeneuve, L.; Hamel, E.; et al. Catechin treatment improves cerebrovascular flow-mediated dilation and learning abilities in atherosclerotic mice. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1032–H1043. [Google Scholar] [CrossRef] [PubMed]
- Widlansky, M.E.; Hamburg, N.M.; Anter, E.; Holbrook, M.; Kahn, D.F.; Elliott, J.G.; Keaney, J.F., Jr.; Vita, J.A. Acute EGCG supplementation reverses endothelial dysfunction in patients with coronary artery disease. J. Am. Coll. Nutr. 2007, 26, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Zsombok, A. Vanilloid receptors—Do they have a role in whole body metabolism? Evidence from TRPV1. J. Diabetes Complications 2013, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.C.; Hsiao, H.C.; Wu, D.C.; Lin, R.J.; Liang, J.C.; Yeh, J.L.; Chen, I.J. A novel capsaicin derivative VOA induced relaxation in rat mesenteric and aortic arteries: Involvement of CGRP, NO, cGMP, and endothelium-dependent activities. J. Cardiovasc Pharmacol 2003, 42, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Ching, L.C.; Kou, Y.R.; Shyue, S.K.; Su, K.H.; Wei, J.; Chen, L.C.; Yu, Y.B.; Pan, C.C.; Lee, T.S. Molecular mechanisms of activation of endothelial nitric oxide synthase mediated by transient receptor potential vanilloid type 1. Cardiovasc. Res. 2011, 91, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Luo, Z.; Ma, S.; Wong, W.T.; Ma, L.; Zhong, J.; He, H.; Zhao, Z.; Cao, T.; Yan, Z.; et al. Activation of TRPV1 by dietary capsaicin improves endothelium-dependent vasorelaxation and prevents hypertension. Cell Metab. 2010, 12, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, P.; Zhao, Z.; Cao, T.; He, H.; Luo, Z.; Zhong, J.; Gao, F.; Zhu, Z.; Li, L.; et al. Activation of transient receptor potential vanilloid 1 by dietary capsaicin delays the onset of stroke in stroke-prone spontaneously hypertensive rats. Stroke 2011, 42, 3245–3251. [Google Scholar] [CrossRef] [PubMed]
- Fan, N.C.; Tsai, C.M.; Hsu, C.N.; Huang, L.T.; Tain, Y.L. N-acetylcysteine prevents hypertension via regulation of the ADMA-DDAH pathway in young spontaneously hypertensive rats. Biomed. Res. Int. 2013, 2013, 696317. [Google Scholar] [PubMed]
- Vasdev, S.; Singal, P.; Gill, V. The antihypertensive effect of cysteine. Int. J. Angiol. 2009, 18, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, R.V.; Patel, S.G.; Guthikonda, A.P.; Reid, M.; Balasubramanyam, A.; Taffet, G.E.; Jahoor, F. Deficient synthesis of glutathione underlies oxidative stress in aging and can be corrected by dietary cysteine and glycine supplementation. Am. J. Clin. Nutr. 2011, 94, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.; del, A.C.; Lahera, V.; Ruilope, L.M. N-acetylcysteine potentiates the antihypertensive effect of angiotensin converting enzyme inhibitors. Am. J. Hypertens. 1995. [Google Scholar] [CrossRef]
- Barrios, V.; Calderon, A.; Navarro-Cid, J.; Lahera, V.; Ruilope, L.M. N-acetylcysteine potentiates the antihypertensive effect of ACE inhibitors in hypertensive patients. Blood Press. 2002, 11, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Martina, V.; Masha, A.; Gigliardi, V.R.; Brocata, L.; Manzato, E.; Berchio, A.; Massarenti, P.; Settanni, F.; Della Casa, L.; Bergamini, S.; et al. Long-term N-acetylcysteine and L-arginine administration reduces endothelial activation and systolic blood pressure in hypertensive patients with type 2 diabetes. Diabetes Care 2008, 31, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Vasdev, S.; Stuckless, J. Antihypertensive effects of dietary protein and its mechanism. Int. J. Angiol. 2010, 19, e7–e20. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.; Stamler, J.; Dyer, A.R.; Appel, L.; Dennis, B.; Kesteloot, H.; Ueshima, H.; Okayama, A.; Garside, D.B.; Zhou, B. Association between protein intake and blood pressure: The INTERMAP Study. Arch. Intern. Med. 2006, 166, 79–87. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Gu, D.; Wu, X.; Chen, J.; Duan, X.; Chan, J.; Whelton, P.K. Effect of soybean protein on blood pressure: A randomized, controlled trial. Ann. Intern. Med. 2005, 143, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bindoli, A.; Rigobello, M.P. Principles in redox signaling: From chemistry to functional significance. Antioxid. Redox Signal. 2013, 18, 1557–1593. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.M.; Carroll, K.S. The redox biochemistry of protein sulfenylation and sulfinylation. J. Biol. Chem. 2013, 288, 26480–26408. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, D.A.; Forman, H.J. Glutathione in defense and signaling: Lessons from a small thiol. Ann. NY Acad. Sci. 2002, 973, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Shelton, M.D.; Chock, P.B.; Mieyal, J.J. Glutaredoxin: Role in reversible protein s-glutathionylation and regulation of redox signal transduction and protein translocation. Antioxid. Redox Signal. 2005, 7, 348–366. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.M. The paradox of high risk of stroke in populations with low risk of coronary heart disease. Am. J. Epidemiol. 1990, 131, 579–588. [Google Scholar] [PubMed]
- Bailey, E.L.; Smith, C.; Sudlow, C.L.; Wardlaw, J.M. Is the spontaneously hypertensive stroke prone rat a pertinent model of sub cortical ischemic stroke? A systematic review. Int. J. Stroke 2011, 6, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Itoh, T.; Tabuchi, M.; Satou, T.; Ezaki, O. Dietary protein, but not carbohydrate, is a primary determinant of the onset of stroke in stroke-prone spontaneously hypertensive rats. Stroke 2009, 40, 2828–2835. [Google Scholar] [CrossRef] [PubMed]
- Ueshima, H.; Iida, M.; Shimamoto, T.; Konishi, M.; Tsujioka, K.; Tanigaka, M.; Nakanishi, N.; Ozawa, H.; Kojima, S.; Komachi, Y. Multivariate analysis of risk factors for stroke. Eight-year follow-up study of farming villages in Akita, Japan. Prev. Med. 1980, 9, 722–740. [Google Scholar] [CrossRef] [PubMed]
- Yamori, Y.; Kihara, M.; Fujikawa, J.; Soh, Y.; Ohtaka, M.; Horie, R.; Tsunematsu, T.; Note, S.; Fukase, M. Dietary risk factors of stroke and hypertension in Japan—Part 3: Comparative study on risk factors between farming and fishing villages in japan. Jpn. Circ. J. 1982, 46, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Kagan, A.; Popper, J.S.; Rhoads, G.G.; Yano, K. Dietary and other risk factors for stroke in Hawaiian Japanese men. Stroke 1985, 16, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Sato, S.; Kitamura, A.; Naito, Y.; Shimamoto, T.; Komachi, Y. Fat and protein intakes and risk of intraparenchymal hemorrhage among middle-aged Japanese. Am. J. Epidemiol. 2003, 157, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Sauvaget, C.; Nagano, J.; Hayashi, M.; Yamada, M. Animal protein, animal fat, and cholesterol intakes and risk of cerebral infarction mortality in the adult health study. Stroke 2004, 35, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Virtamo, J.; Wolk, A. Dietary protein intake and risk of stroke in women. Atherosclerosis 2012, 224, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Virtamo, J.; Wolk, A. Red meat consumption and risk of stroke in Swedish women. Stroke 2011, 42, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, A.M.; Pan, A.; Rexrode, K.M.; Stampfer, M.; Hu, F.B.; Mozaffarian, D.; Willett, W.C. Dietary protein sources and the risk of stroke in men and women. Stroke 2012, 43, 637–644. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. IGF-I activity may be a key determinant of stroke risk—A cautionary lesson for vegans. Med. Hypotheses. 2003, 61, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Michell, B.J.; Griffiths, J.E.; Mitchelhill, K.I.; Rodriguez-Crespo, I.; Tiganis, T.; Bozinovski, S.; de Montellano, P.R.; Kemp, B.E.; Pearson, R.B. The Akt kinase signals directly to endothelial nitric oxide synthase. Curr. Biol. 1999, 9, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.T.; Krueger, K.D.; Dhume, A.; Agrawal, D.K. Sustained Akt/PKB activation and transient attenuation of c-jun N-terminal kinase in the inhibition of apoptosis by IGF-1 in vascular smooth muscle cells. Apoptosis 2005, 10, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Conn, K.J.; Rich, C.B.; Jensen, D.E.; Fontanilla, M.R.; Bashir, M.M.; Rosenbloom, J.; Foster, J.A. Insulin-like growth factor-I regulates transcription of the elastin gene through a putative retinoblastoma control element. A role for Sp3 acting as a repressor of elastin gene transcription. J. Biol. Chem. 1996, 271, 28853–28860. [Google Scholar] [CrossRef] [PubMed]
- Mauro, L.; Bartucci, M.; Morelli, C.; Ando, S.; Surmacz, E. IGF-I receptor-induced cell-cell adhesion of MCF-7 breast cancer cells requires the expression of junction protein ZO-1. J. Biol. Chem. 2001, 276, 39892–39897. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.A.; Murata, S.; Nishida, T. Up-regulation of the tight-junction protein ZO-1 by substance P and IGF-1 in A431 cells. Cell Biochem. Funct. 2009, 27, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, S.P.; Hundborg, H.H.; Sorensen, H.T.; Orskov, H.; Tjonnedland, A.; Overvad, K.; Jorgensen, J.O. Insulin-like growth factor (IGF) I, -II, and IGF binding protein-3 and risk of ischemic stroke. J. Clin. Endocrinol. Metab. 2005, 90, 5937–5941. [Google Scholar] [CrossRef] [PubMed]
- Aoi, N.; Nakayama, T.; Soma, M.; Kosuge, K.; Haketa, A.; Sato, M.; Hinohara, S.; Doba, N.; Asai, S. The insulin-like growth factor-1 gene is associated with cerebral infarction in Japanese subjects. Hereditas 2012, 149, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Liu, J.; Li, X.; Peng, J.; Han, S.; Zhang, R.; Xu, Y.; Nie, S. Insulin-like growth factor-1 receptor polymorphism and ischemic stroke: A case-control study in Chinese population. Acta Neurol. Scand. 2008, 118, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Klein, S.; Holloszy, J.O. Long-term low-protein, low-calorie diet and endurance exercise modulate metabolic factors associated with cancer risk. Am. J. Clin. Nutr. 2006, 84, 1456–1462. [Google Scholar] [PubMed]
- McCarty, M.F. Insulin and IGF-I as determinants of low “Western” cancer rates in the rural third world. Int. J. Epidemiol. 2004, 33, 908–910. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Mortality from Western cancers rose dramatically among African-Americans during the 20th century: Are dietary animal products to blame? Med. Hypotheses. 2001, 57, 169–174. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. A low-fat, whole-food vegan diet, as well as other strategies that down-regulate IGF-I activity, may slow the human aging process. Med. Hypotheses. 2003, 60, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A. Impact of reduced insulin-like growth factor-1/insulin signaling on aging in mammals: Novel findings. Aging Cell 2008, 7, 285–290. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; Barroso-Aranda, J.; Contreras, F. The low-methionine content of vegan diets may make methionine restriction feasible as a life extension strategy. Med. Hypotheses. 2009, 72, 125–128. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Vegan proteins may reduce risk of cancer, obesity, and cardiovascular disease by promoting increased glucagon activity. Med. Hypotheses. 1999, 53, 459–485. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Upregulation of lymphocyte apoptosis as a strategy for preventing and treating autoimmune disorders: A role for whole-food vegan diets, fish oil and dopamine agonists. Med. Hypotheses. 2001, 57, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Barnard, N.D.; Cohen, J.; Jenkins, D.J.; Turener-McGrievy, G.; Gloede, L.; Jaster, B.; Seidl, K.; Green, A.A.; Talpers, S. A low-fat vegan diet improves glycemic control and cardiovascular risk factors in a randomized clinical trial in individuals with type 2 diabetes. Diabetes Care 2006, 29, 1777–1783. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.C.; Junshi, C. Diet and chronic degenerative diseases: Perspectives from China. Am. J. Clin. Nutr. 1994, 59, 1153S–1161S. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCarty, M.F. NADPH Oxidase Activity in Cerebral Arterioles Is a Key Mediator of Cerebral Small Vessel Disease—Implications for Prevention. Healthcare 2015, 3, 233-251. https://doi.org/10.3390/healthcare3020233
McCarty MF. NADPH Oxidase Activity in Cerebral Arterioles Is a Key Mediator of Cerebral Small Vessel Disease—Implications for Prevention. Healthcare. 2015; 3(2):233-251. https://doi.org/10.3390/healthcare3020233
Chicago/Turabian StyleMcCarty, Mark F. 2015. "NADPH Oxidase Activity in Cerebral Arterioles Is a Key Mediator of Cerebral Small Vessel Disease—Implications for Prevention" Healthcare 3, no. 2: 233-251. https://doi.org/10.3390/healthcare3020233
APA StyleMcCarty, M. F. (2015). NADPH Oxidase Activity in Cerebral Arterioles Is a Key Mediator of Cerebral Small Vessel Disease—Implications for Prevention. Healthcare, 3(2), 233-251. https://doi.org/10.3390/healthcare3020233