
Numerical Simulation of Higher-Order Nonlinearity of Human Brain Functional Connectivity Using Hypergraph p-Laplacian


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Hypergraph Network Notation of the Human Brain
3. The Hypergraph p-Laplacian Diffusion Model
3.1. The Hypergraph Laplacian Diffusion Model
3.2. The Hypergraph p-Laplacian Diffusion Model
3.3. The Computation of Hypergraph p-Laplacian
| Algorithm 1 The estimation of hypergraph p-Laplacian |
| Input: The structural connectivity network and three parameters |
| Output: Hypergraph p-Laplacian: |
| Step 1: Initialize the hypergraph Laplacian matrix and adjacent matrix from the |
| structural connectivity network. |
| Step 2: Decomposition of the hypergraph Laplacian: . |
| Initialize: . |
| Step 3: While not converged do: |
| , where is given by Equation (10) |
| End |
| Step 4: |
| Return: |
4. Experiments
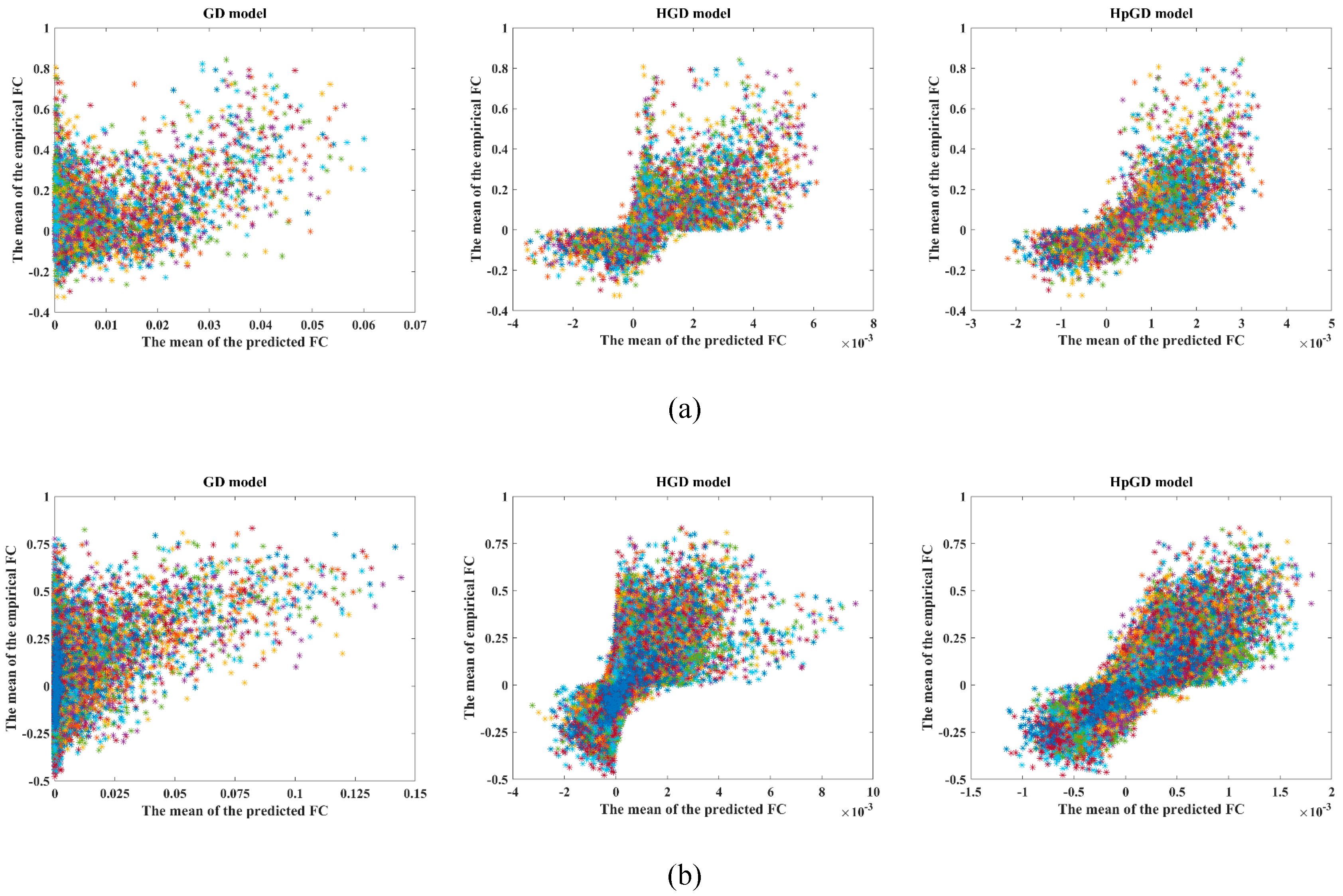
4.1. Correlations with the Experiment FC
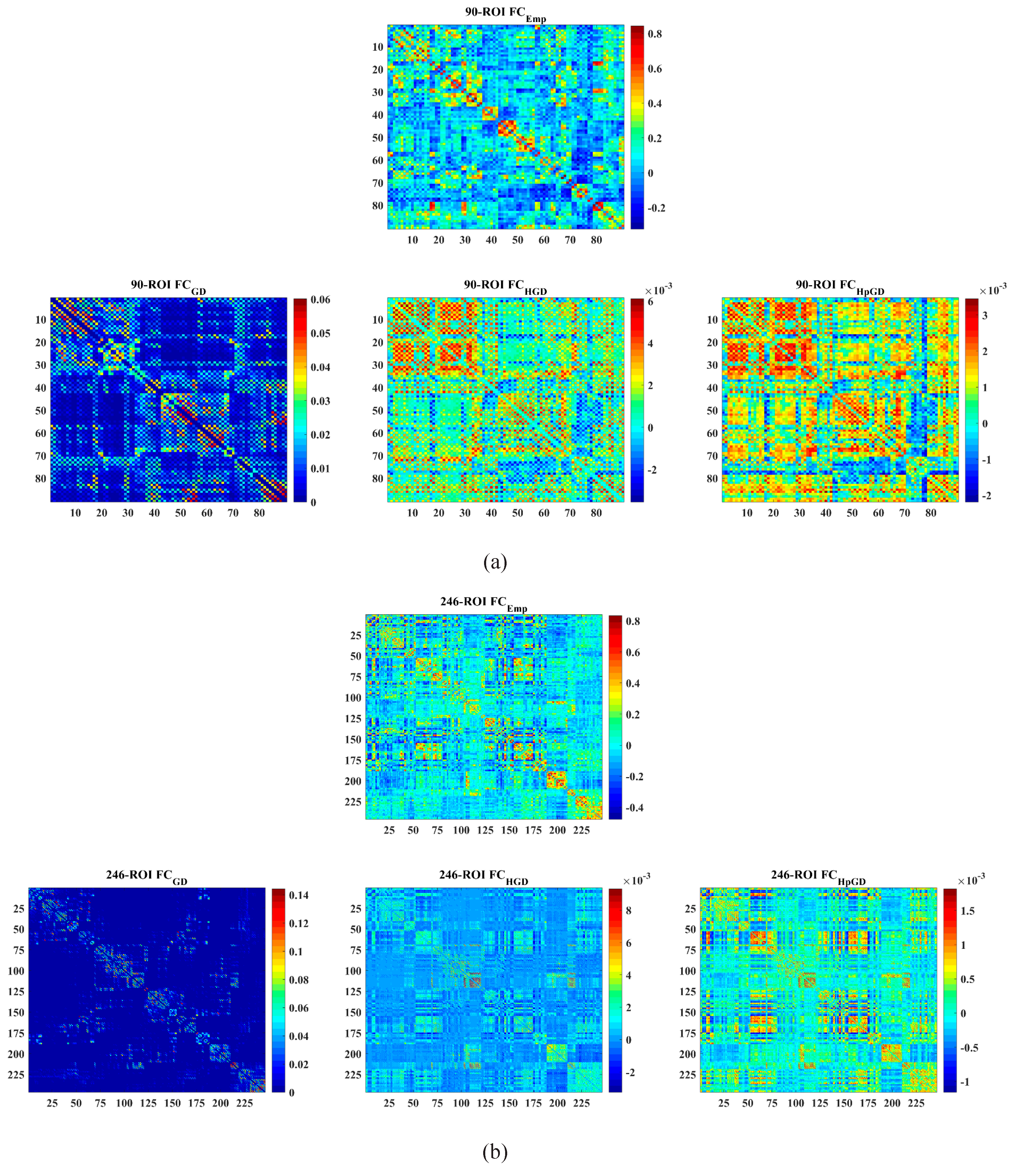
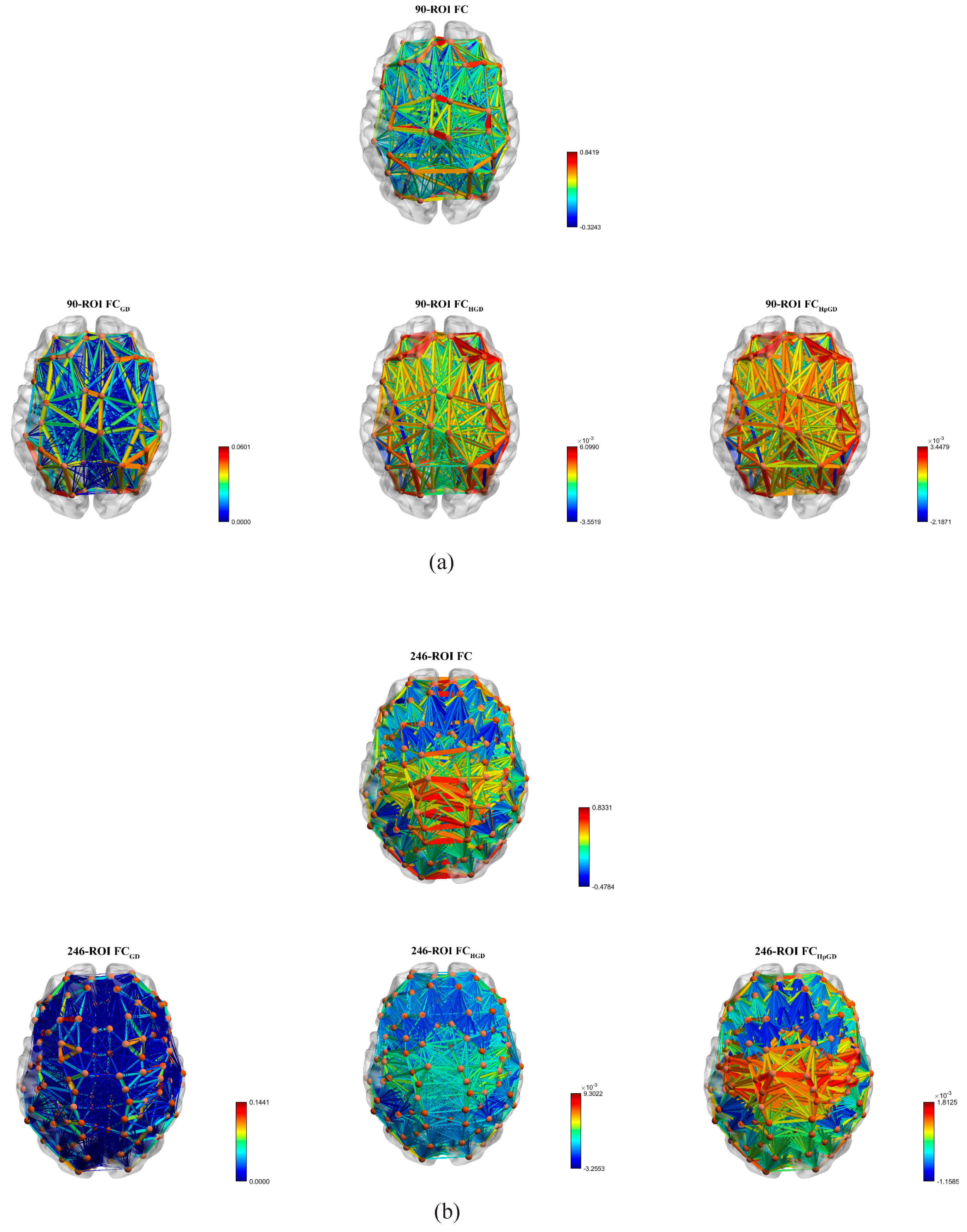
4.2. Mean FC Network
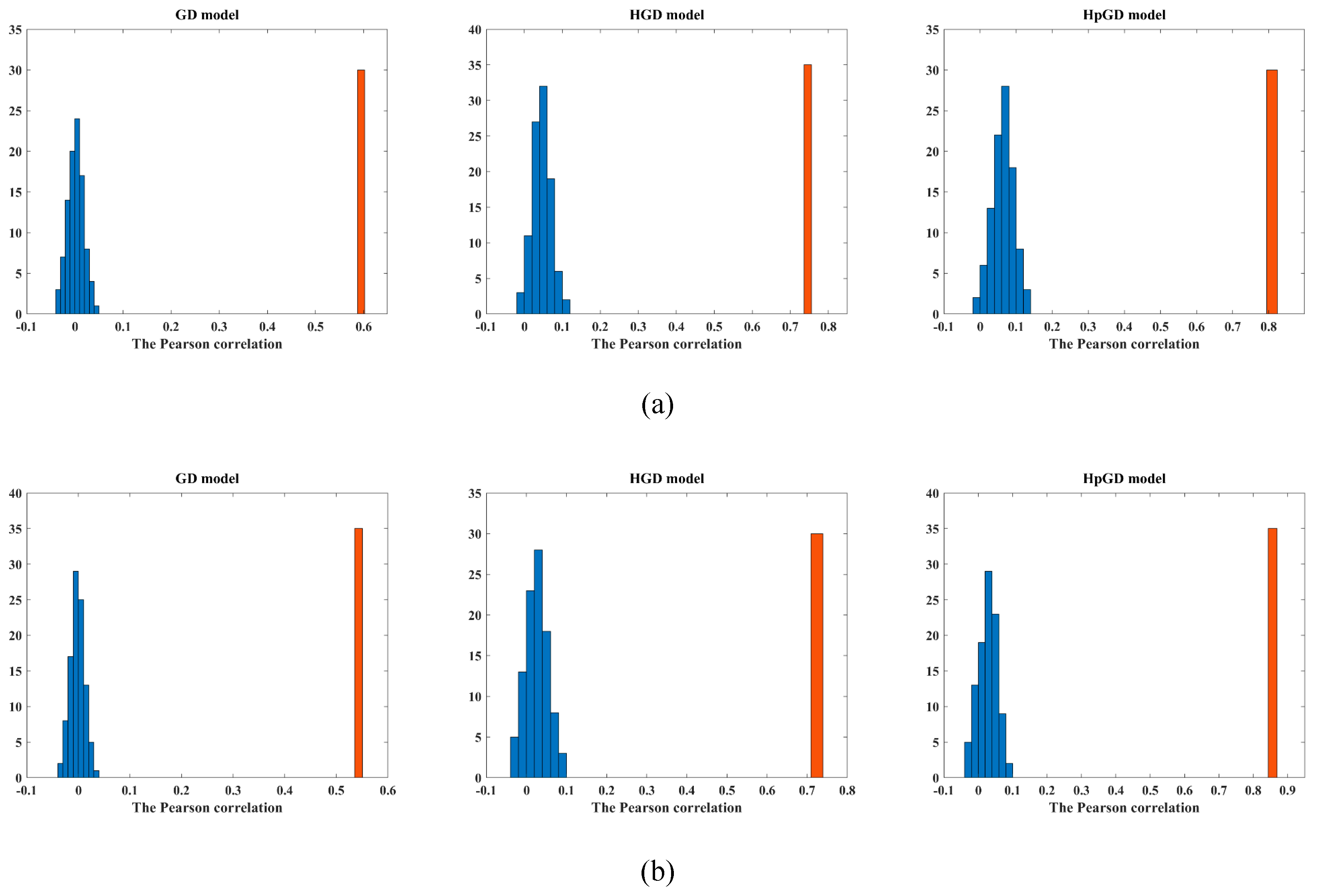
4.3. Stability and Robustness Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Uddin, L.Q. Complex relationships between structural and functional brain connectivity. Trends Cogn. Sci. 2013, 17, 600–602. [Google Scholar] [CrossRef] [Green Version]
- Iturria-Medina, Y.; Sotero, R.C.; Canales-Rodrguez, E.J.; Aleman-Gomez, Y.; Melie-Garcia, L. Studying the human brain anatomical network via diffusion-weighted MRI and graph theory. NeuroImage 2008, 40, 1064–1076. [Google Scholar] [CrossRef]
- Zhu, D.J.; Zhang, T.; Jiang, X.; Hu, X.T.; Chen, H.B.; Yang, N.; Lv, J.L.; Han, J.W.; Guo, L.; Liu, T.M. Fusing DTI and fMRI data: A survey of methods and applications. NeuroImage 2014, 102, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eavani, H.; Satterthwaite, T.D.; Filipovych, R.; Gur, R.E.; Gur, R.C.; Davatzikos, C. Identifying sparse connectivity patterns in the brain using resting-state fMRI. NeuroImage 2015, 105, 286–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, L.Q.; Mooshagian, E.; Zaidel, E.; Scheres, A.; Margulies, D.S.; Kelly, A.C.; Shehzad, Z.; Adelstein, J.S.; Castellanos, F.X.; Biswal, B.B.; et al. Residual functional connectivity in the split-brain revealed with resting-state functional MRI. Neuroreport 2008, 19, 703–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewarie, P.; Hillebrand, A.; van Dellen, E.; Schoonheim, M.M.; Barkhof, F.; Polman, C.H.; Beaulieu, C.; Gong, G.; van Dijk, B.W.; Stam, C.J. Structural degree predicts functional network connectivity: A multimodal resting-state fMRI and MEG study. NeuroImage 2014, 97, 296–307. [Google Scholar] [CrossRef]
- Stam, C.J.; van Straaten, E.C.W.; van Dellen, E.; Tewarie, P.; Gong, G.; Hillebrand, A.; Meier, J.; Van Mieghem, P. The relation between structural and functional connectivity patterns in complex brain networks. Int. J. Psychophysiol. 2016, 103, 149–160. [Google Scholar] [CrossRef]
- Ventrella, A.V.; Piro, G.; Grieco, L.A. On modeling shortest path length distribution in scale-free network topologies. IEEE Syst. J. 2018, 12, 3869–3872. [Google Scholar] [CrossRef]
- Goni, J.; van den Heuvel, M.P.; Avena-Koenigsberger, A.; de Mendizabal, N.V.; Betzel, R.F.; Griffa, A.; Hagmann, P.; Corominas-Murtra, B.; Thiran, J.P.; Sporns, O. Resting-brain functional connectivity predicted by analytic measures of network communication. Proc. Natl. Acad. Sci. USA 2014, 111, 833–838. [Google Scholar] [CrossRef] [Green Version]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting human resting-state functional connectivity from structural connectivity. Proc. Natl. Acad. Sci. USA 2009, 106, 2035–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deco, G.; Ponce-Alvarez, A.; Mantini, D.; Romani, G.L.; Hagmann, P.; Corbetta, M. Resting-state functional connectivity emerges from structurally and dynamically shaped slow linear fluctuations. J. Neurosci. 2013, 33, 11239–11252. [Google Scholar] [CrossRef] [PubMed]
- Messe, A.; Rudrauf, D.; Giron, A.; Marrelec, G. Predicting functional connectivity from structural connectivity via computational models using MRI: An extensive comparison study. NeuroImage 2015, 111, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, G.; Vasa, F.; Griffa, A.; Hagmann, P.; Amico, E.; Goni, J.; Avidan, G.; Sporns, O. Mapping higher-order relations between brain structure and function with embedded vector representations of connectomes. Nat. Commun. 2018, 9, 2178. [Google Scholar] [CrossRef]
- Surampudi, S.G.; Misra, J.; Deco, G.; Bapi, R.S.; Sharma, A.; Roy, D. Resting state dynamics meets anatomical structure: Temporal multiple kernel learning (tmkl) model. NeuroImage 2019, 184, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Parhi, K.K.; Lenglet, C. Function-specific and enhanced brain structural connectivity mapping via joint modeling of diffusion and functional MRI. Sci. Rep. 2018, 8, 4741. [Google Scholar] [CrossRef] [Green Version]
- Abdelnour, F.; Voss, H.U.; Raj, A. Network diffusion accurately models the relationship between structural and functional brain connectivity networks. Neuroimage 2014, 90, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Abdelnour, F.; Dayan, M.; Devinsky, O.; Thesen, T.; Raj, A. Functional brain connectivity is predictable from anatomic network’s Laplacian eigen-structure. Neuroimage 2018, 172, 728–739. [Google Scholar] [CrossRef]
- Ma, J.C.; Wang, Y.J.; Liu, B.D.; Liu, W.F. Accurately modeling the human brain functional correlations with hypergraph Laplacian. Neurocomputing 2021, 428, 239–247. [Google Scholar] [CrossRef]
- Raj, A.; Kuceyeski, A.; Weiner, M. A network diffusion model of disease progression in dementia. Neuron 2012, 73, 1204–1215. [Google Scholar] [CrossRef] [Green Version]
- Buhler, T.; Hein, M. Spectral clustering based on the graph p-Laplacian. In Proceedings of the 26th Annual International Conference on Machine Learning, Montreal, QC, Canada, 14–18 June 2009; pp. 81–88. [Google Scholar]
- Ma, X.Q.; Liu, W.F.; Li, S.Y.; Tao, D.P.; Zhou, Y.C. Hypergraph p-Laplacian regularization for remotely sensed image recognition. IEEE T. Geosci. Remote. 2019, 57, 1585–1595. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.C.; Liu, W.F.; Zhou, Y.C.; Nie, L.Q. HplapGCN: Hypergraph p-Laplacian graph convolutional networks. Neurocomputing 2019, 362, 166–174. [Google Scholar] [CrossRef]
- Zhou, D.Y.; Huang, J.Y.; Schölkopf, B. Learning with hypergraphs: Clustering, classification, and embedding. In Proceedings of the 19th International Conference on Neural Information Processing Systems, Vancouver, BC, Canada, 4–7 December 2006; pp. 1601–1608. [Google Scholar]
- Lin, Q.X.; Dai, Z.J.; Xia, M.R.; Han, Z.Z.; Huang, R.W.; Gong, G.L.; Liu, C.; Bi, Y.C.; He, Y. A connectivity-based test-retest dataset of multi-modal magnetic resonance imaging in young healthy adults. Sci. Data 2015, 2, 150056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.Z.; Li, H.; Zhuo, J.J.; Zhang, Y.; Wang, J.J.; Chen, L.F.; Yang, Z.Y.; Chu, C.Y.; Xie, S.M.; Laird, A.R.; et al. The human brainnetome atlas: A new brain atlas based on connectional architecture. Cereb. Cortex 2016, 26, 3508–3526. [Google Scholar] [CrossRef]
- Xia, M.R.; Wang, J.H.; He, Y. Brainnet viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Du, C.; Liu, W.; Wang, Y. Numerical Simulation of Higher-Order Nonlinearity of Human Brain Functional Connectivity Using Hypergraph p-Laplacian. Mathematics 2021, 9, 2345. https://doi.org/10.3390/math9182345
Ma J, Du C, Liu W, Wang Y. Numerical Simulation of Higher-Order Nonlinearity of Human Brain Functional Connectivity Using Hypergraph p-Laplacian. Mathematics. 2021; 9(18):2345. https://doi.org/10.3390/math9182345
Chicago/Turabian StyleMa, Jichao, Chunyu Du, Weifeng Liu, and Yanjiang Wang. 2021. "Numerical Simulation of Higher-Order Nonlinearity of Human Brain Functional Connectivity Using Hypergraph p-Laplacian" Mathematics 9, no. 18: 2345. https://doi.org/10.3390/math9182345
APA StyleMa, J., Du, C., Liu, W., & Wang, Y. (2021). Numerical Simulation of Higher-Order Nonlinearity of Human Brain Functional Connectivity Using Hypergraph p-Laplacian. Mathematics, 9(18), 2345. https://doi.org/10.3390/math9182345