Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture


Abstract
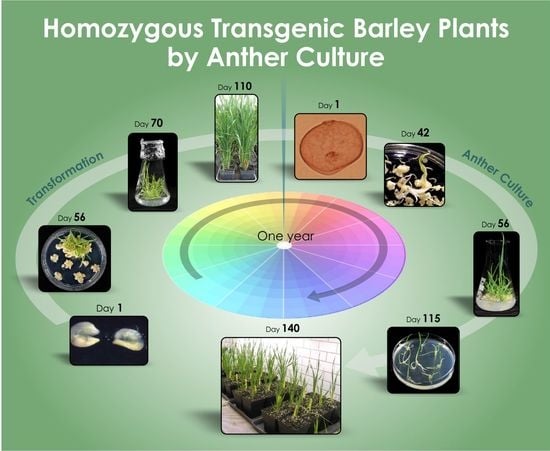
1. Introduction
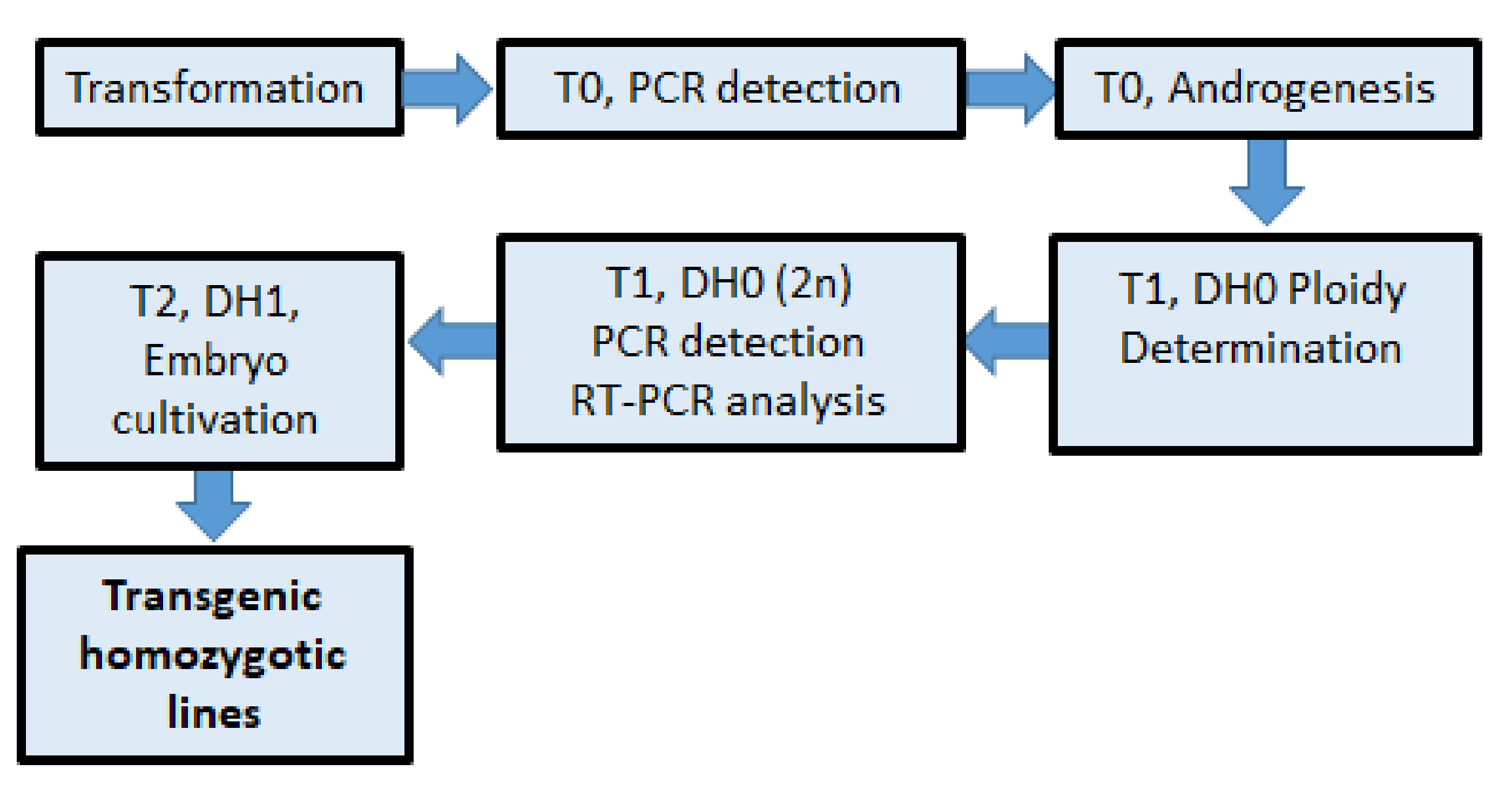
2. Results
2.1. Anther Cultures of Transgenic Plants
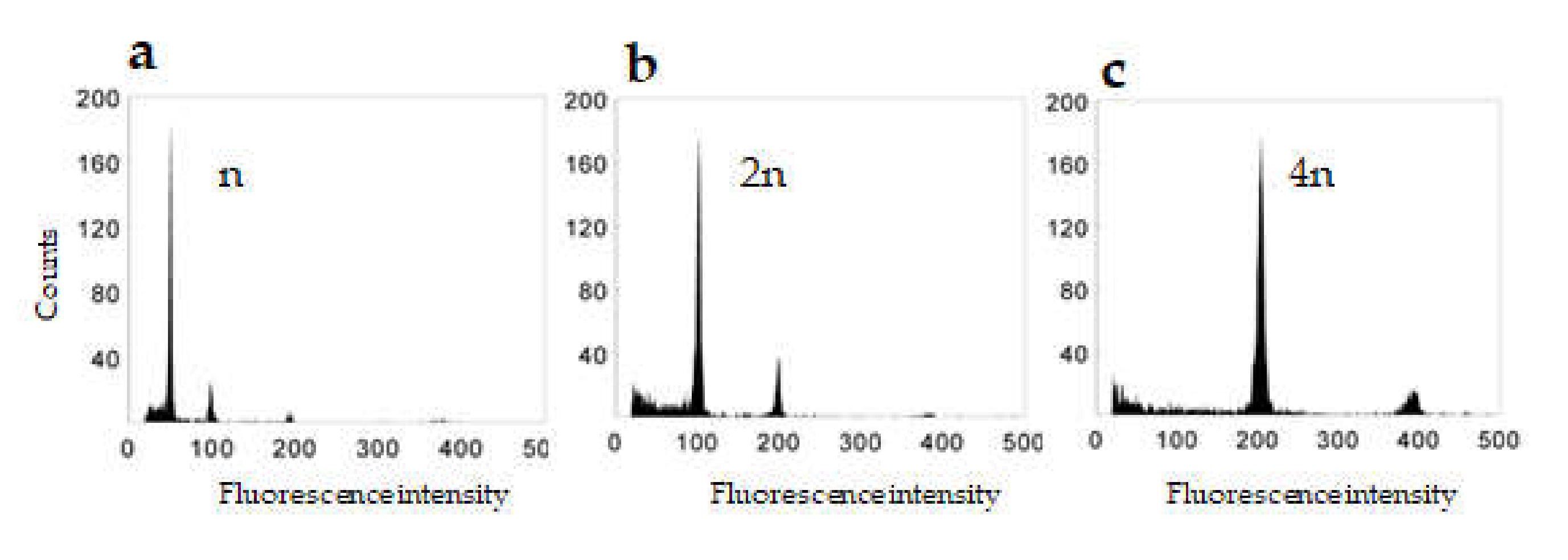
2.2. Ploidy Determination
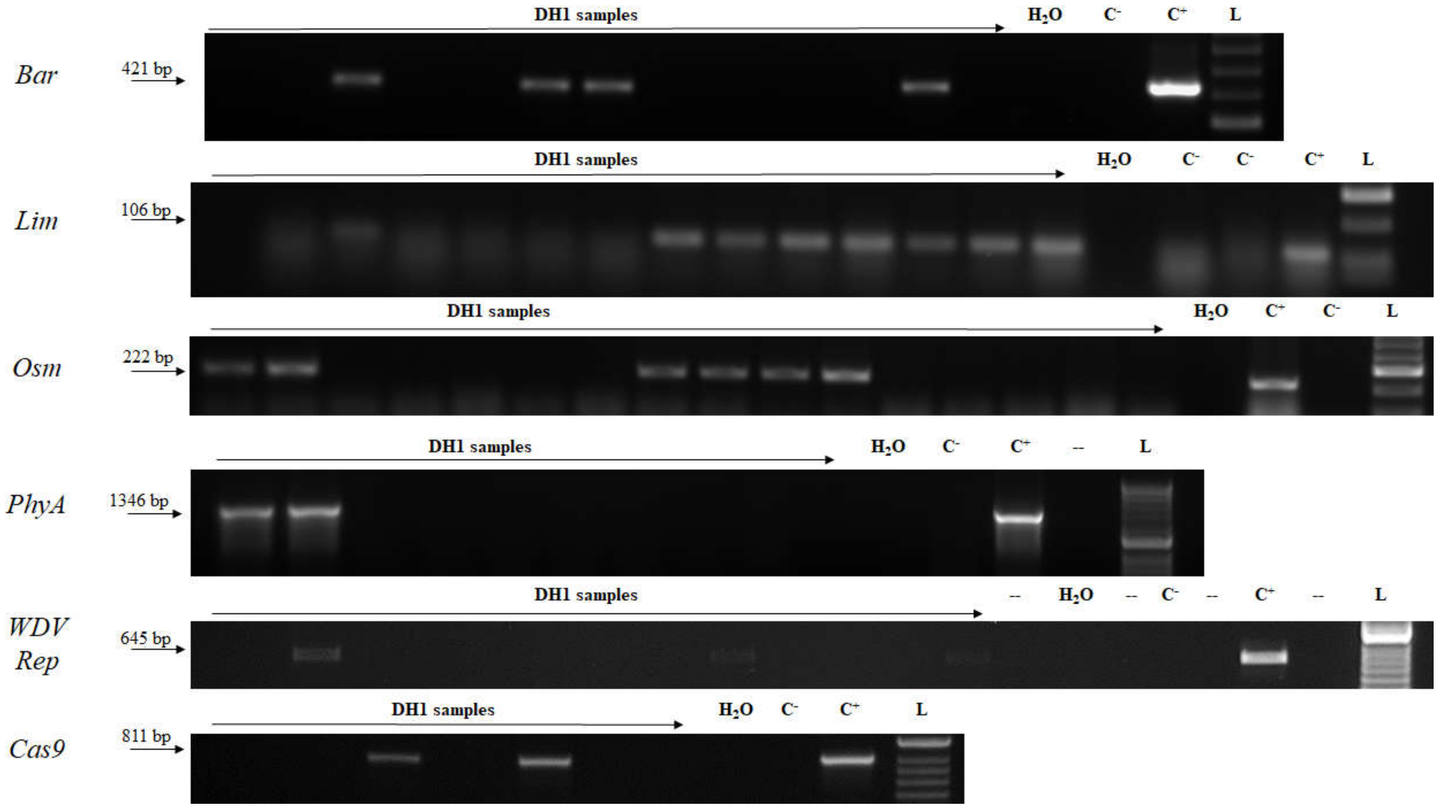
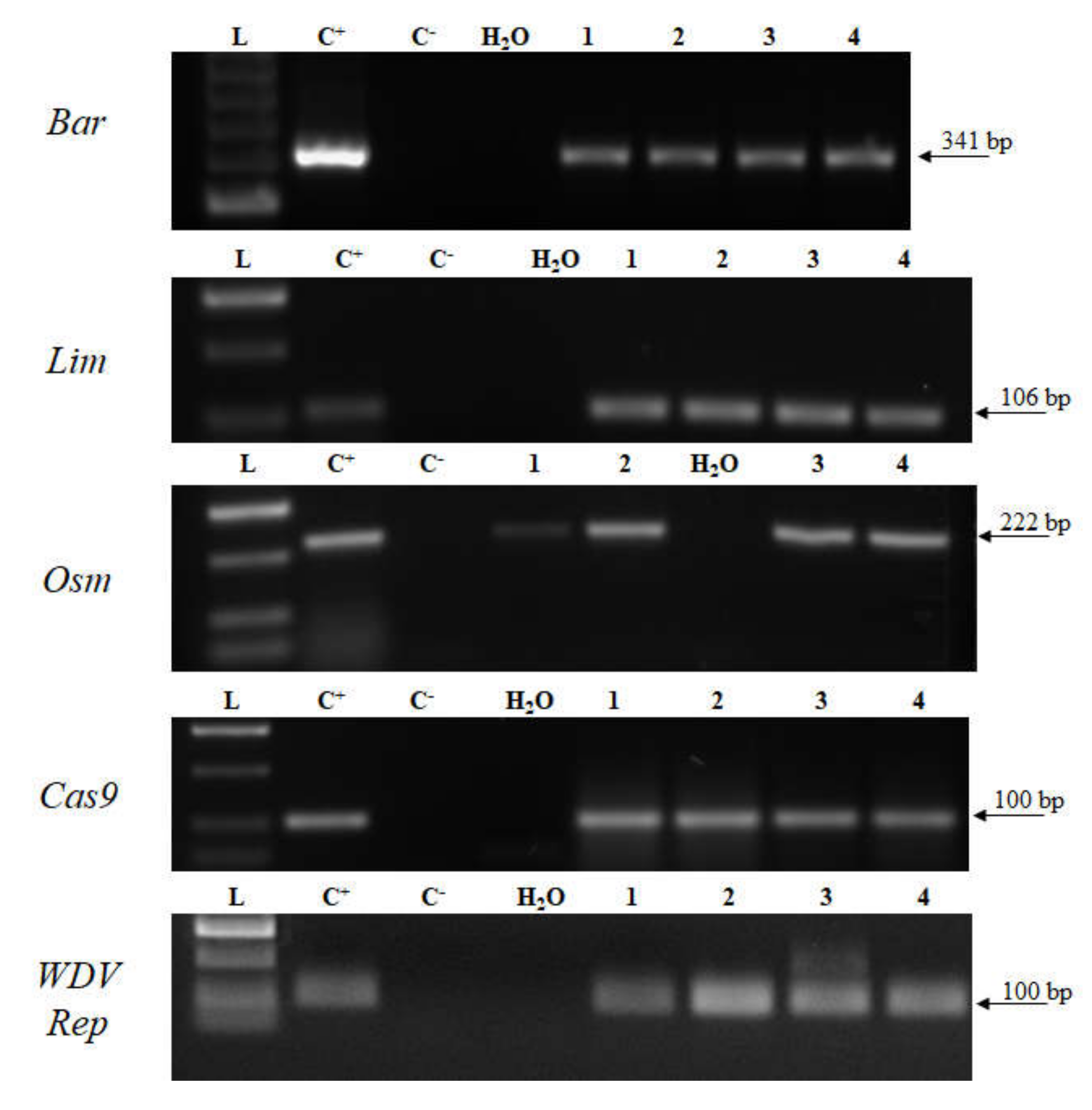
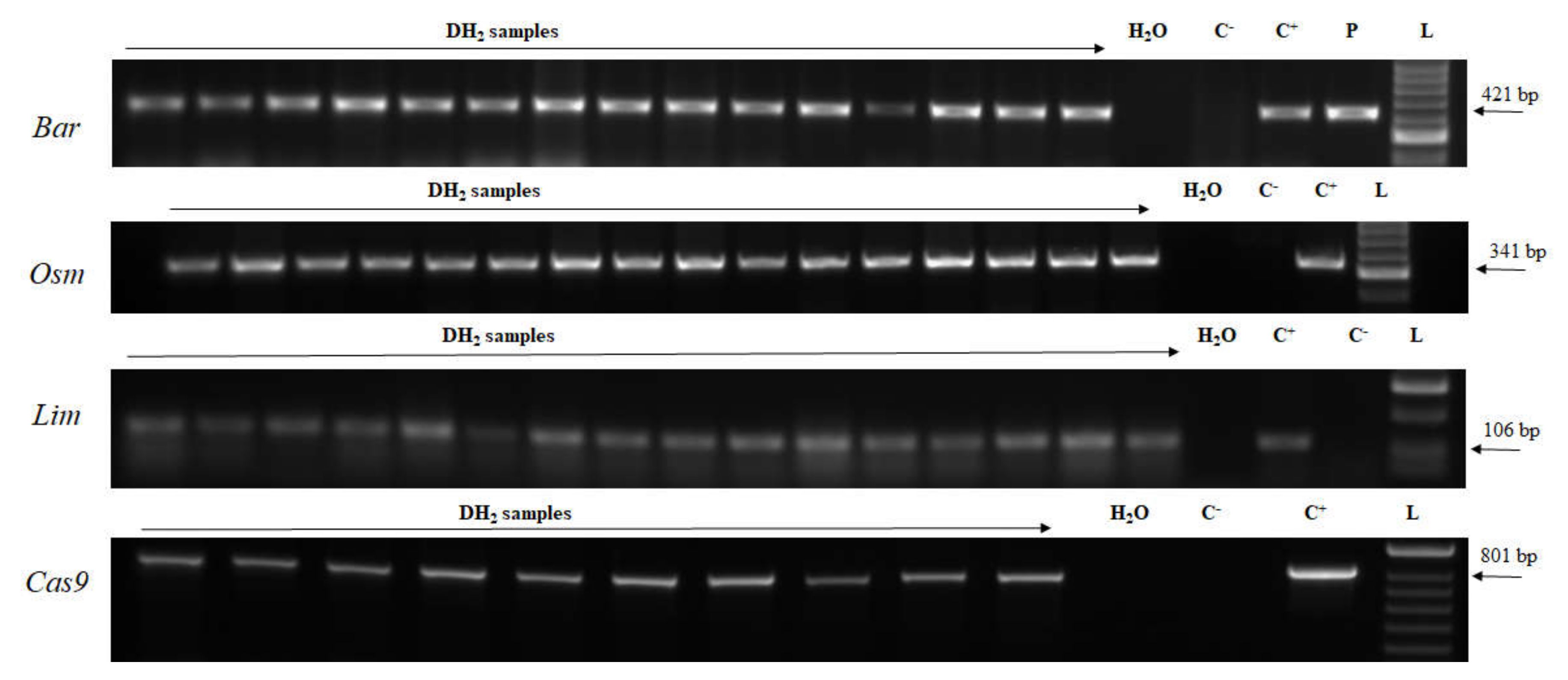
2.3. Genotyping
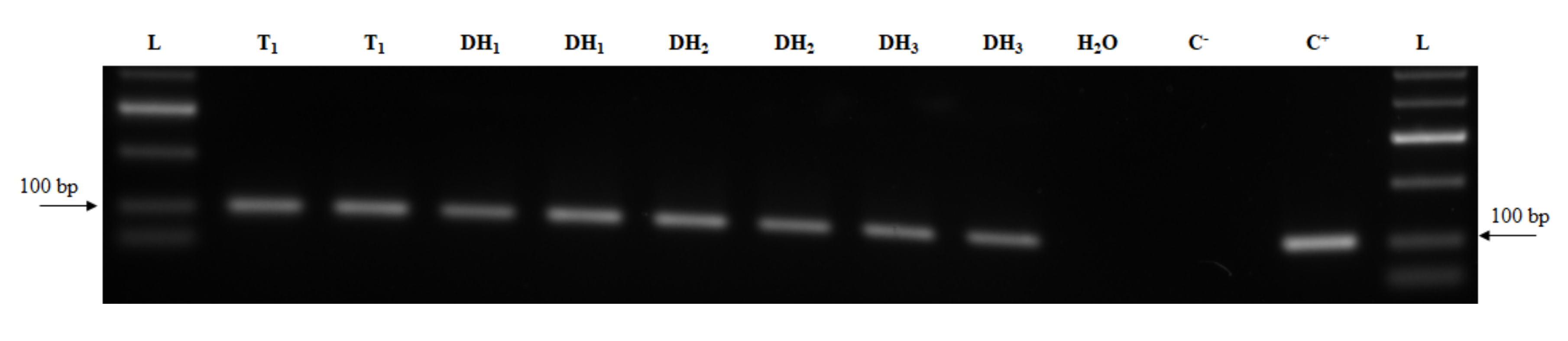
2.4. Transcript Detection
2.5. Embryo Cultures
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Anther Cultures
4.3. Detection of Transgenic Plants
4.4. Ploidy Level
4.5. Detection of Transcripts
4.6. Embryo Culture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, H. In vitro induced haploids in wheat. In In Vitro Haploid Production in Higher Plants: Volume 4—Cereals; Jain, S.M., Sopory, S.K., Veilleux, R.E., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 73–97. ISBN 978-94-017-1862-2. [Google Scholar] [CrossRef]
- Devaux, P.; Pickering, R. Haploids in the improvement of poaceae. In Haploids in Crop Improvement II. Biotechnology in Agriculture and Forestry; Don Palmer, C.E., Keller, W.A., Kasha, K.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 215–242. ISBN 978-3-540-26889-5. [Google Scholar] [CrossRef]
- Yan, G.; Liu, H.; Wang, H.; Lu, Z.; Wang, Y.; Mullan, D.; Hamblin, J.; Liu, C. Accelerated generation of selfed pure line plants for gene identification and crop breeding. Front. Plant Sci. 2017, 8, 1786. [Google Scholar] [CrossRef] [PubMed]
- Mullerova, E.; Novotny, J.; Vagera, J.; Harwood, W.A.; Ohnoutkova, L. Induction of androgenesis in transgenic barley plants. In Proceedings of the Induction of Androgenesis in Transgenic Barley Plants, Bled, Slovenia, 1–5 July 2000; pp. 29–32. [Google Scholar]
- Coronado, M.J.; Hensel, G.; Broeders, S.; Otto, I.; Kumlehn, J. Immature pollen-derived doubled haploid formation in barley cv. Golden Promise as a tool for transgene recombination. Acta Physiol. Plant. 2005, 27, 591–599. [Google Scholar] [CrossRef]
- Kapusi, E.; Hensel, G.; Coronado, M.J.; Broeders, S.; Marthe, C.; Otto, I.; Kumlehn, J. The elimination of a selectable marker gene in the doubled haploid progeny of co-transformed barley plants. Plant Mol. Biol. 2013, 81, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Kumlehn, J.; Serazetdinova, L.; Hensel, G.; Becker, D.; Loerz, H. Genetic transformation of barley (Hordeum vulgare L.) via infection of androgenetic pollen cultures with Agrobacterium tumefaciens. Plant Biotechnol. J. 2006, 4, 251–261. [Google Scholar] [CrossRef]
- Holme, I.B.; Brinch-Pedersen, H.; Lange, M.; Holm, P.B. Transformation of different barley (Hordeum vulgare L.) cultivars by Agrobacterium tumefaciens infection of in vitro cultured ovules. Plant Cell Rep. 2008, 27, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Brew-Appiah, R.A.T.; Ankrah, N.; Liu, W.; Konzak, C.F.; Von Wettstein, D.; Rustgi, S. Generation of doubled haploid transgenic wheat lines by microspore transformation. PLoS ONE 2013, 8, e80155. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, P.; Ellison, E.; Polley, B.; Bollina, V.; Kulkarni, M.; Ghanbarnia, K.; Song, H.; Gao, C.; Voytas, D.F.; Kagale, S. Targeted mutagenesis in wheat microspores using CRISPR/Cas9. Sci. Rep. 2018, 8, 6502. [Google Scholar] [CrossRef]
- Cegielska-Taras, T.; Pniewski, T.; Szała, L. Transformation of microspore-derived embryos of winter oilseed rape (Brassica napus L.) by using Agrobacterium tumefaciens. J. Appl. Genet. 2008, 49, 343–347. [Google Scholar] [CrossRef]
- Chen, C.; Xiao, H.; Zhang, W.; Wang, A.; Xia, Z.; Li, X.; Zhai, W.; Cheng, Z.; Zhu, L. Adapting rice anther culture to gene transformation and RNA interference. Sci. China Ser. C Life Sci. 2006, 49, 414–428. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Britt, A.B.; Tripathi, L.; Sharma, S.; Upadhyaya, H.D.; Ortiz, R. Haploids: Constraints and opportunities in plant breeding. Biotechnol. Adv. 2015, 33, 812–829. [Google Scholar] [CrossRef]
- Lantos, C.; Purgel, S.; Ács, K.; Langó, B.; Bóna, L.; Boda, K.; Békés, F.; Pauk, J. Utilization of in vitro anther culture in spelt wheat breeding. Plants 2019, 8, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Lantos, C.; Pauk, J. Anther culture as an effective tool in winter wheat (Triticum aestivum L.) breeding. Russ. J. Genet. 2016, 52, 794–801. [Google Scholar] [CrossRef]
- Shen, Y.; Pan, G.; Lubberstedt, T. Haploid strategies for functional validation of plant genes. Trends Biotechnol. 2015, 33, 611–620. [Google Scholar] [CrossRef]
- Murovec, J.; Bohanec, B. Haploids and doubled haploids in plant breeding. In Plant Breeding; Abdurakhmonov, I.Y., Ed.; IntechOpen: Rijeka, Croatia, 2012; ISBN 978-953-307-932-5. [Google Scholar] [CrossRef]
- Mohammadi, P.P.; Moieni, A.; Ebrahimi, A.; Javidfar, F. Doubled haploid plants following colchicine treatment of microspore-derived embryos of oilseed rape (Brassica napus L.). Plant Cell. Tissue Organ Cult. 2012, 108, 251–256. [Google Scholar] [CrossRef]
- Soriano, M.; Cistué, L.; Vallés, M.P.; Castillo, A.M. Effects of colchicine on anther and microspore culture of bread wheat (Triticum aestivum L.). Plant Cell. Tissue Organ Cult. 2007, 91, 225–234. [Google Scholar] [CrossRef]
- Ohnoutkova, L.; Vlcko, T.; Mentewab, A. Barley anther culture. In Barley: Methods and Protocols; Harwood, W.A., Ed.; Springer Science & Business Media: New York, NY, USA, 2019; pp. 37–52. ISBN 978-1-4939-8942-3. [Google Scholar] [CrossRef]
- Melander, M.; Kamnert, I.; Happstadius, I.; Liljeroth, E.; Bryngelsson, T. Stability of transgene integration and expression in subsequent generations of doubled haploid oilseed rape transformed with chitinase and β-1,3-glucanase genes in a double-gene construct. Plant Cell Rep. 2006, 25, 942–952. [Google Scholar] [CrossRef]
- Kim, K.M.; Baenziger, P.S.; Rybczynski, J.J.; Arumuganathan, K. Characterization of ploidy levels of wheat microspore-derived plants using laser flow cytometry. Vitr. Cell. Dev. Biol.-Plant 2003, 39, 663–668. [Google Scholar] [CrossRef]
- Makowska, K.; Oleszczuk, S.; Zimny, A.; Czaplicki, A.; Zimny, J. Androgenic capability among genotypes of winter and spring barley. Plant Breed. 2015, 134, 668–674. [Google Scholar] [CrossRef]
- Sriskandarajah, S.; Sameri, M.; Lerceteau-Köhler, E.; Westerbergh, A. Increased recovery of green doubled haploid plants from barley anther culture. Crop Sci. 2015, 55, 2806–2812. [Google Scholar] [CrossRef]
- Gajecka, M.; Marzec, M.; Chmielewska, B.; Jelonek, J.; Zbieszczyk, J.; Szarejko, I. Plastid differentiation during microgametogenesis determines green plant regeneration in barley microspore culture. Plant Sci. 2020, 291, 110321. [Google Scholar] [CrossRef]
- Cejnar, P.; Ohnoutková, L.; Ripl, J.; Vlčko, T.; Kundu, J.K. Two mutations in the truncated Rep gene RBR domain delayed the Wheat dwarf virus infection in transgenic barley plants. J. Integr. Agric. 2018, 17, 2492–2500. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Patat-Ochatt, E.M.; Moessner, A. Ploidy level determination within the context of in vitro breeding. Plant Cell. Tissue Organ Cult. 2011, 104, 329–341. [Google Scholar] [CrossRef]
- Davies, G.J.; Sheikh, M.A.; Ratcliffe, O.J.; Coupland, G.; Furner, I.J. Genetics of homology-dependent gene silencing in Arabidopsis; a role for methylation. Plant J. 1997, 12, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Elmayan, T.; Vaucheret, H. Expression of single copies of a strongly expressed 35S transgene can be silenced post-transcriptionally. Plant J. 1996, 9, 787–797. [Google Scholar] [CrossRef]
- Viktorova, J.; Klcova, B.; Rehorova, K.; Vlcko, T.; Stankova, L.; Jelenova, N.; Cejnar, P.; Kundu, J.K.; Ohnoutkova, L.; Macek, T. Recombinant expression of osmotin in barley improves stress resistance and food safety during adverse growing conditions. PLoS ONE 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Rehorova, K.; Viktorova, J.; Macuchova, B.; Vlcko, T.; Stankova, L.; Jelenova, N.; Ohnoutkova, L.; Macek, T. Limen, non-toxic recombinant plant defensin and its effect against pathogenic yeast and fungi. J. Pharm. Pharmacol. 2018, 6, 1–11. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Dolezel, J.; Gohde, W. Sex determination in dioecious plants melandrium album and m. rubrum using high-resolution flow cytometry. Cytometry 1995, 19, 103–106. [Google Scholar] [CrossRef]
- Masek, T.; Vopalensky, V.; Suchomelova, P.; Pospisek, M. Denaturing RNA electrophoresis in TAE agarose gels. Anal. Biochem. 2005, 336, 46–50. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transgene | No. of Cultivated Anthers | Regenerated Plants % (No.) | Ploidy Level Green Plants % (No.) | Transgenic Plants 2n % (No.) | ||||
|---|---|---|---|---|---|---|---|---|
| Green | Albino | n | 2n | 4n | Aneupl | |||
| Bar | 3547 | 1.3 (46) | 5.2 (184) | 39.1 (18) | 52.2 (24) | 6.5 (3) | 2.1(1) | 58.3 (14) |
| Lim | 2428 | 1.3 (31) | 4.4 (109) | 38.7 (12) | 45.2(14) | 6.6 (2) | 9.7(3) | 50.0 (7) |
| Osm | 1209 | 2.8 (35) | 3.0 (36) | 42.8 (15) | 54.3 (19) | 0 | 2.8 (1) | 73.7 (14) |
| PhyA | 612 | 2.6 (16) | 5.3 (33) | 50.0 (8) | 43.8 (7) | 6.3 (1) | 0 | 85.7 (6) |
| WDV Rep | 797 | 4.6 (37) | 27.8 (222) | 70.0 (26) | 30.0 (11) | 0 | 0 | 54.5 (6) |
| Cas9 | 1288 | 4.0 (52) | 1.2 (15) | 55.8 (29) | 44.2 (23) | 0 | 0 | 47.8 (11) |
| Transgene | Primer Pair 5′–3′ Oriented | PCR Amplicon | TA [°C] |
|---|---|---|---|
| Bar | GGT CTG CAC CAT CGT CAA CC | 421 bp | 65 |
| GTC ATG CCA GTT CCC GTG CT | |||
| Lim | AGC TTG GTA CCA CCA TGC AC | 106 bp | 57 |
| CAG TGG TCG TCG CAG TTA GA | |||
| Osm | CTC CTC GAC GGC TTC AAC AT | 341 bp | 57 |
| TCG AGT GGG AAG TTT GGG TG | |||
| GCC CTG CCT TCA TAC GCT AT | 222 bp | 58 | |
| TAC GGG CAG TTG TTC CTC AC | |||
| PhyA | GGC AGT CCC CGC CTC GAG AAA | 1346 bp | 55 |
| AAA CAC TCC GCC CAA TCA CCC | |||
| WDV Rep | ATGGCCTCTTCATCTGCACC | 645 bp | 63 |
| TGATTCGAGGCTTACGGAGT | |||
| Cas9 | TTC GCT ACT GTT CGC AAG GT | 811 bp | 58 |
| Transgene | Primer Pair 5′–3′ Oriented | PCR Amplicon |
|---|---|---|
| Bar | GGT CTG CAC CAT CGT CAA CC | 421 bp |
| GTC ATG CCA GTT CCC GTG CT | ||
| Lim | AGC TTG GTA CCA CCA TGC AC | 106 bp |
| CAG TGG TCG TCG CAG TTA GA | ||
| Osm | GCC CTG CCT TCA TAC GCT AT | 222 bp |
| TAC GGG CAG TTG TTC CTC AC | ||
| Cas9 | CGA CGC TAC TCT CAT CCA CC | 100 bp |
| CTT TTT GGT GGC AGC AGG AC | ||
| WDV Rep | TTC ATC TGC ACC CAG GTT CC | 100 bp |
| GTG CGA AGT GAA TCC AAG GC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohnoutková, L.; Vlčko, T. Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants 2020, 9, 918. https://doi.org/10.3390/plants9070918
Ohnoutková L, Vlčko T. Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants. 2020; 9(7):918. https://doi.org/10.3390/plants9070918
Chicago/Turabian StyleOhnoutková, Ludmila, and Tomáš Vlčko. 2020. "Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture" Plants 9, no. 7: 918. https://doi.org/10.3390/plants9070918
APA StyleOhnoutková, L., & Vlčko, T. (2020). Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants, 9(7), 918. https://doi.org/10.3390/plants9070918