Expression of Gibberellin Metabolism Genes and Signalling Components in Dwarf Phenotype of Breadfruit (Artocarpus altilis) Plants Growing on Marang (Artocarpus odoratissimus) Rootstocks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
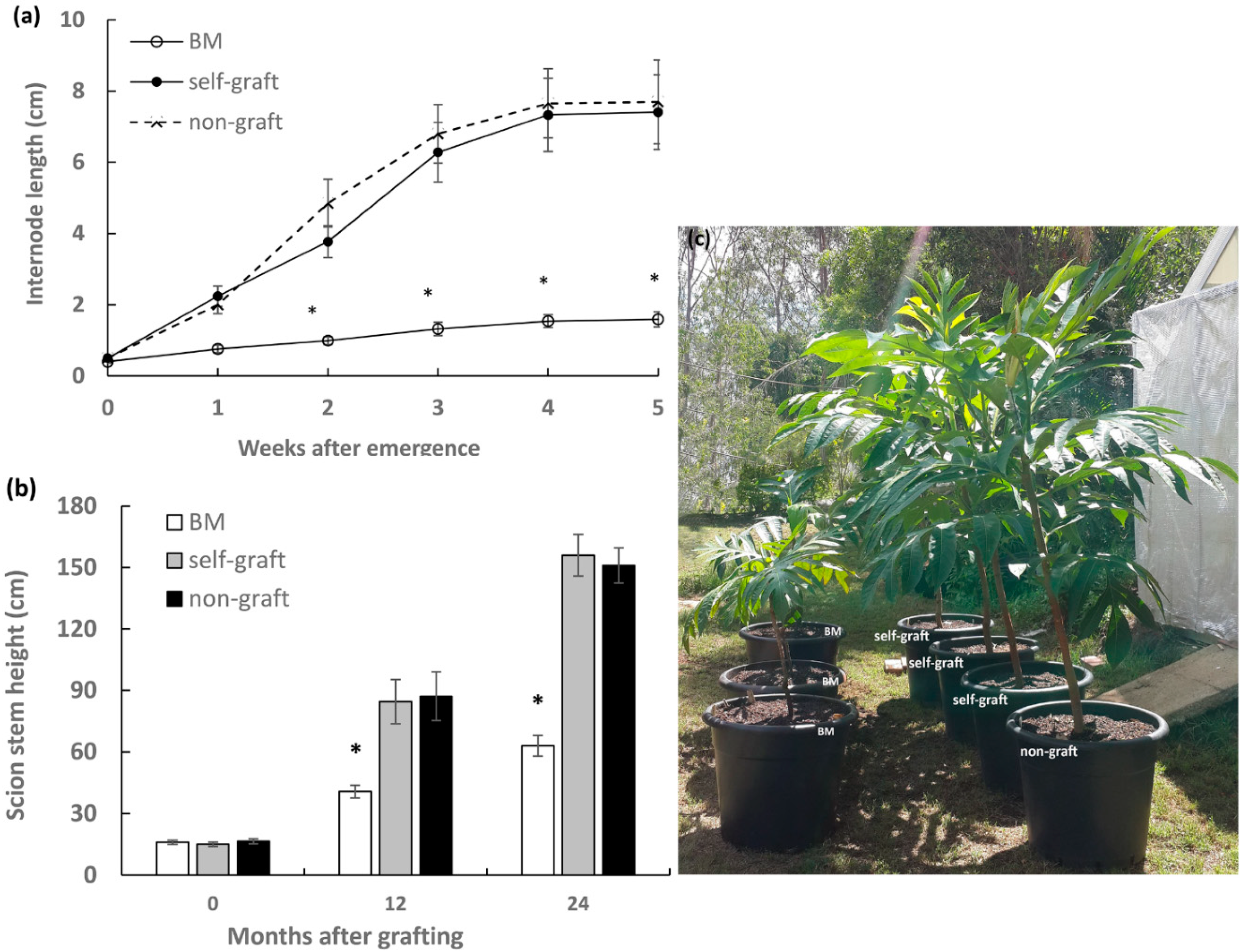
2.1. Effect of Rootstocks on Stem Elongation of Breadfruit Scions
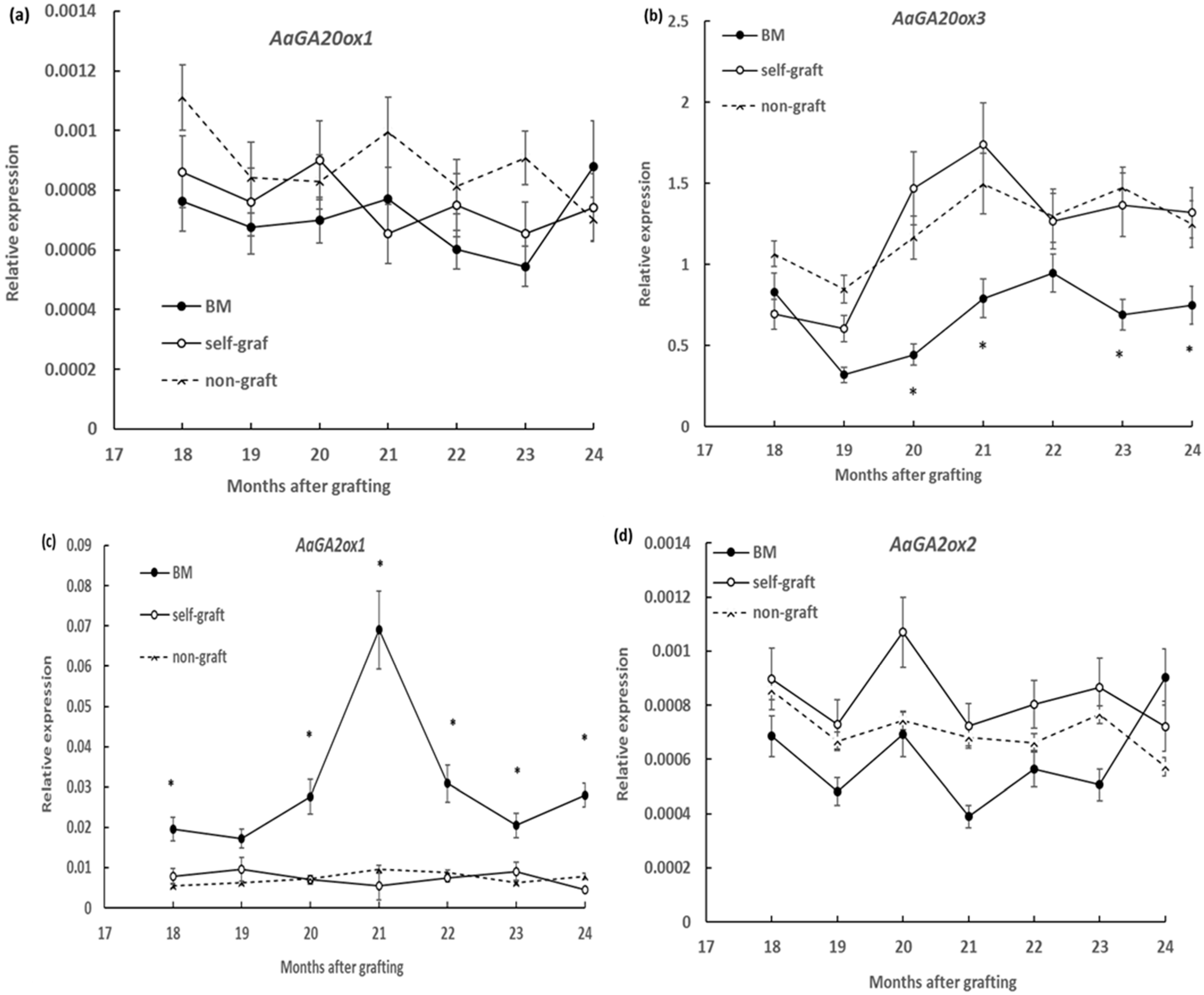
2.2. Effect of Rootstocks on the Expression of GA Metabolic Genes
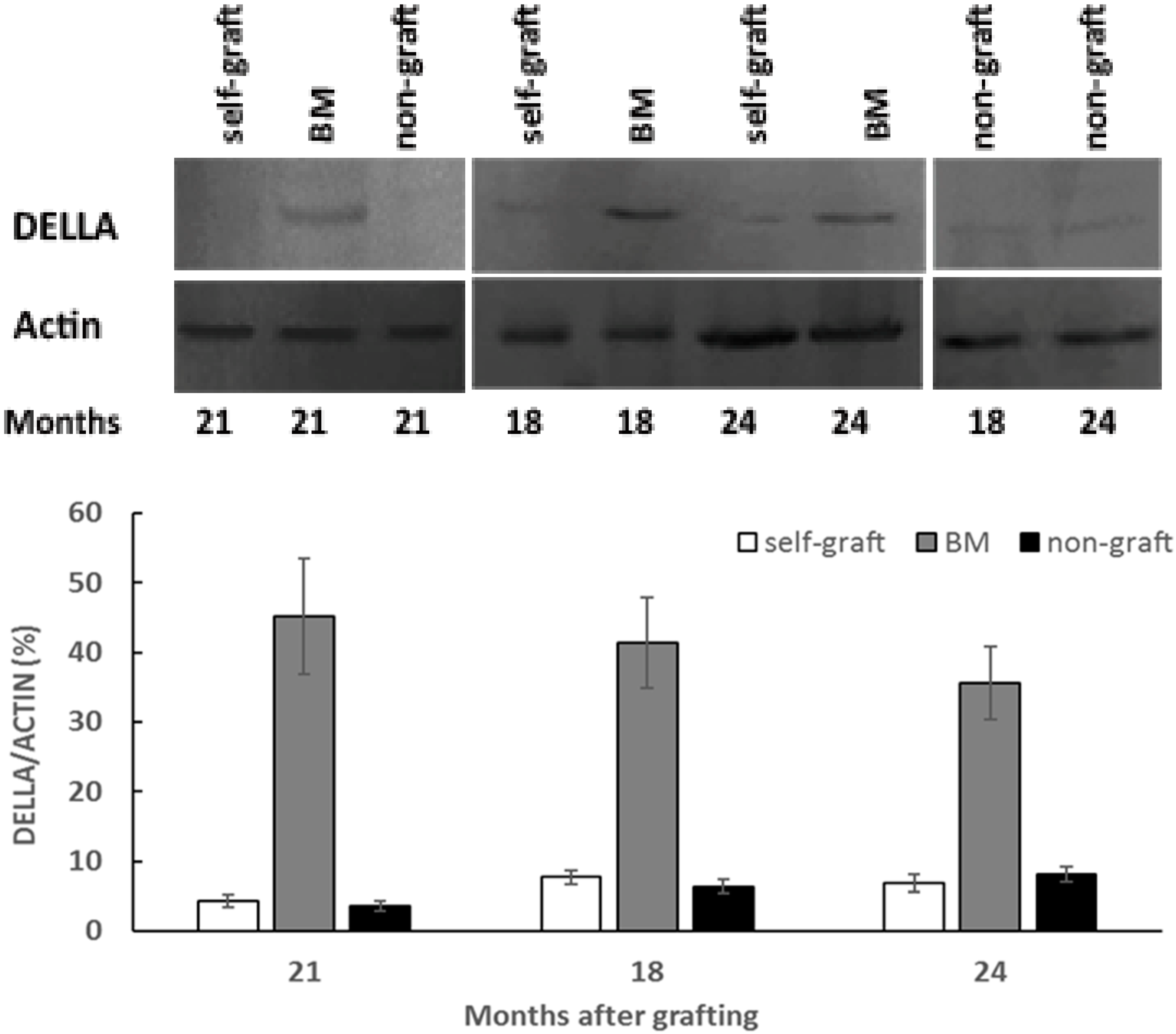
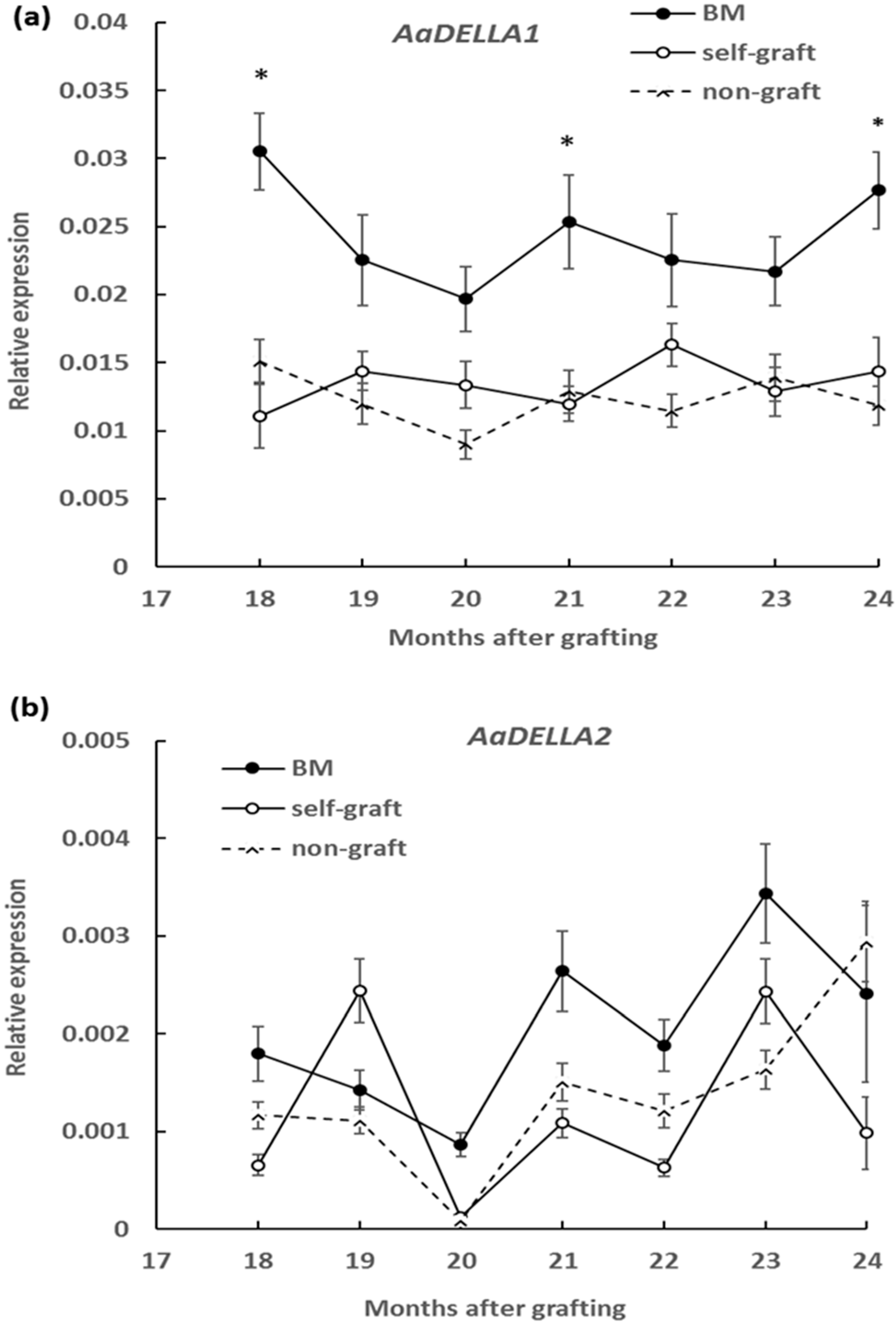
2.3. Effect of Rootstocks on DELLA Protein Abundance and Transcript Levels
2.4. Restoration of Stem Elongation by GA Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Quantitative Real-Time PCR
4.3. Cloning of DELLA cDNAs from “Gold Noli” Breadfruit
4.4. Western Blotting
4.5. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lochard, R.G.; Schneider, G.W. Stock and scion growth relationships and the dwarfing mechanism in apple. Hortic. Rev. 1981, 3, 315–375. [Google Scholar] [CrossRef]
- Soumelidou, K.; Battey, N.H.; John, P.; Barnett, J.R. The Anatomy of the developing bud union and its relationship to dwarfing in apple. Ann. Bot. 1994, 74, 605–611. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Else, M.A.; Taylor, L.; Dover, C.J. Root and stem hydraulic conductivity as determinants of growth potential in grafted trees of apple (Malus pumila Mill.). J. Exp. Biol. 2003, 54, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.; Atkinson, C.; Bengough, A.; Else, M.; Fernández-Fernández, F.; Harrison, R.; Schmidt, S. Contributions of roots and rootstocks to sustainable, intensified crop production. J. Exp. Bot. 2013, 64, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Ragone, D.; Leibniz Institute of Plant Genetics and Crop Plant Research (IPK). Breadfruit, Artocarpus altilis (Parkinson) Fosberg. Promot. Conserv. Use Underutilized Negl. Crop. 1997, 10, 77. [Google Scholar]
- Zhou, Y.; Underhill, S.J.R. A dwarf phenotype identified in breadfruit (Artocarpus altilis) plants growing on marang (A. odoratissimus) rootstocks. Horticulturae 2019, 5, 40. [Google Scholar] [CrossRef]
- Forster, J.R.; Forster, G. Characteres Generum Plantarum, Quas in Itinere Ad Insulas Maris Australis, Collegerunt, Descripserunt, Delinearunt, Annis 1772–1775; Missouri Botanical Garden: Saint Louis, MO, USA, 1776. [Google Scholar]
- Basile, B.; Marsal, J.; Solar, L.I.; Tyree, M.T.; Bryla, D.R.; Dejong, T.M. Hydraulic conductance of peach trees grafted on rootstocks with differing size-controlling potentials. J. Hortic. Sci. Biotech. 2003, 78, 768–774. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G. Hormone and growth interactions of scions and size-controlling rootstocks of young apple trees. Plant Growth Regul. 2016, 78, 105–119. [Google Scholar] [CrossRef]
- Richards, D.; Thompson, W.K.; Pharis, R.P. The influence of dwarfing interstocks on the distribution and metabolism of xylem-applied [3H]gibberellin A4 in apple. Plant Physiol. 1986, 82, 1090–1095. [Google Scholar] [CrossRef]
- van Hooijdonk, B.; Woolley, D.; Warrington, I.; Tustin, S. Rootstocks Modify Scion Architecture, Endogenous Hormones, and Root Growth of Newly Grafted ‘Royal Gala’ Apple Trees. J. Am. Soc. Hortic. Sci. 2011, 136, 93–102. [Google Scholar] [CrossRef]
- Van Hooijdonk, B.M.; Woolley, D.J.; Warrington, I.J.; Tustin, D.S. Initial alteration of scion architecture by dwarfing apple rootstocks may involve shoot-root-shoot signalling by auxin, gibberellin, and cytokinin. J. Hortic. Sci. Biotech. 2010, 85, 59–65. [Google Scholar] [CrossRef]
- Yadava, U.L.; Lockard, R.G. Abscisic Acid and Gibberellin-in Three Ungrafted Apple (Malus sylvestris) Rootstock Clones. Physiol. Plant 1977, 40, 225–229. [Google Scholar] [CrossRef]
- Foster, T.; Kirk, C.; Jones, W.T.; Allan, A.C.; Espley, R.; Karunairetnam, S.; Rakonjac, J. Characterisation of the DELLA subfamily in apple (Malus x domestica Borkh.). Tree Genet. Genomes 2007, 3, 187–197. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Zhuang, W.B.; Tu, X.T.; Gao, Z.H.; Xiong, A.S.; Yu, X.Y.; Li, X.H.; Li, F.H.; Qu, S.C. Transcriptomic analysis of interstock-induced dwarfism in Sweet Persimmon (Diospyros kaki Thunb.). Hortic. Res. 2019, 6, 51. [Google Scholar] [CrossRef]
- El-Sharkawy, I.; El Kayal, W.; Prasath, D.; Fernandez, H.; Bouzayen, M.; Svircev, A.M.; Jayasankar, S. Identification and genetic characterization of a gibberellin 2-oxidase gene that controls tree stature and reproductive growth in plum. J. Exp. Bot. 2012, 63, 1225–1239. [Google Scholar] [CrossRef]
- Hedden, P.; Phillips, A. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Hedden, P.; Kamiya, Y. Gibberellin biosynthesis: Enzymes, genes and their regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 431–460. [Google Scholar] [CrossRef]
- Bulley, S.M.; Wilson, F.M.; Hedden, P.; Phillips, A.L.; Croker, S.J.; James, D.J. Modification of gibberellin biosynthesis in the grafted apple scion allows control of tree height independent of the rootstock. Plant Biotechnol. J. 2005, 3, 215–223. [Google Scholar] [CrossRef]
- Fagoaga, C.; Tadeo, F.R.; Iglesias, D.J.; Huerta, L.; Lliso, I.; Vidal, A.M.; Talon, M.; Navarro, L.; Garcia-Martinez, J.L.; Pena, L. Engineering of gibberellin levels in citrus by sense and antisense overexpression of a GA 20-oxidase gene modifies plant architecture. J. Exp. Bot. 2007, 58, 1407–1420. [Google Scholar] [CrossRef]
- Busov, V.B.; Meilan, R.; Pearce, D.W.; Ma, C.; Rood, S.B.; Strauss, S.H. Activation tagging of a dominant gibberellin catabolism gene (GA 2-oxidase) from poplar that regulates tree stature. Plant Physiol. 2003, 132, 1283–1291. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Ariizumi, T.; Steber, C.M. Gibberellin signaling: A theme and variations on DELLA repression. Plant Physiol. 2012, 160, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Ueguchi-Tanaka, M.; Sata, Y.; Ashikari, M.; Matsuoka, M. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. Plant Cell 2002, 14, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Boss, P.K.; Thomas, M.R. Association of dwarfism and floral induction with a grape ‘green revolution’ mutation. Nature 2002, 416, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.Q.; Ma, C.; Kadmiel, M.; Gai, Y.; Strauss, S.; Jiang, X.N.; Busov, V. Tissue-specific expression of Populus C(19) GA 2-oxidases differentially regulate above- and below-ground biomass growth through control of bioactive GA concentrations. New Phytol. 2011, 192, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Eriksson, S.; Powers, S.J.; Gong, F.; Griffiths, J.; Woolley, L.; Benlloch, R.; Nilsson, O.; Thomas, S.G.; Hedden, P.; et al. Genetic analysis reveals that C(19)-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis. Plant Cell 2008, 20, 2420–2436. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S. Breadfruit (Artocarpus altilis) gibberellin 20-oxidase genes: Sequence variants, stem elongation and abiotic stress response. Tree Genet. Genomes 2015, 11, 1–13. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S. Breadfruit (Artocarpus altilis) gibberellin 2-oxidase genes in stem elongation and abiotic stress response. Plant Physiol. Biochem. 2016, 98, 81–88. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S. Breadfruit (Artocarpus altilis) DELLA genes: Gibberellin-regulated stem elongation and response to high salinity and drought. Plant Growth Regul. 2017, 83, 375–383. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomass production and xylem fiber length. Nat. Biotechnol. 2000, 18, 784–788. [Google Scholar] [CrossRef]
- Busov, V.; Meilan, R.; Pearce, D.W.; Rood, S.B.; Ma, C.P.; Tschaplinski, T.J.; Strauss, S.H. Transgenic modification of gai or rgl1 causes dwarfing and alters gibberellins, root growth, and metabolite profiles in Populus. Planta 2006, 224, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, H.T.; Li, J.J.; Wang, B.; Dai, C.; Wang, J.; Liu, K.D. Brassica napus DS-3, encoding a DELLA protein, negatively regulates stem elongation through gibberellin signaling pathway. Theor. Appl. Genet. 2017, 130, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.-Y.; Shang, J.-X.; Oh, E.; Fan, M.; Bai, Y.; Zentella, R.; Sun, T.-P.; Wang, Z.-Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in Arabidopsis. Nat. Cell Biol. 2012, 14, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Ma, N.; Pei, H.X.; Chen, J.W.; Li, J.; Gao, J.P. A DELLA gene, RhGAI1, is a direct target of EIN3 and mediates ethylene-regulated rose petal cell expansion via repressing the expression of RhCesA2. J. Exp. Bot. 2013, 64, 5075–5084. [Google Scholar] [CrossRef]
- Willige, B.C.; Isono, E.; Richter, R.; Zourelidou, M.; Schwechheimer, C. Gibberellin regulates PIN-FORMED abundance and is required for auxin transport-dependent growth and development in Arabidopsis thaliana. Plant Cell 2011, 23, 2184–2195. [Google Scholar] [CrossRef]
- Oh, E.; Yamaguchi, S.; Hu, J.; Yusuke, J.; Jung, B.; Paik, I.; Lee, H.-S.; Sun, T.-p.; Kamiya, Y.; Choi, G. PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds. Plant Cell 2007, 19, 1192–1208. [Google Scholar] [CrossRef]
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin localization and transport in plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef]
- Haywood, V.; Yu, T.-S.; Huang, N.-C.; Lucas, W.J. Phloem long-distance trafficking of GIBBERELLIC ACID-INSENSITIVE RNA regulates leaf development. Plant J. 2005, 42, 49–68. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S.J.R. Plasma membrane H-ATPase activity and graft success of breadfruit (Artocarpus altilis) onto interspecific rootstocks of marang (A. odoratissimus) and pedalai (A. sericicarpus). Plant Biol. 2018, 20, 978–985. [Google Scholar] [CrossRef]
- Nicot, N.; Hausman, J.-F.; Hoffmann, L.; Evers, D.l. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- Wang, W.; Vignani, R.; Scali, M.; Cresti, M. A universal and rapid protocol for protein extraction from recalcitrant plant tissues for proteomic analysis. Electrophoresis 2006, 27, 2782–2786. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Thomas, M.; Hetherington, L.; Wang, X.-D.; Offler, C.; Patrick, J. Genotypic differences in seed growth rates of Phaseolus vulgaris L. Factors contributing to cotyledon sink activity and sink size. Aust. J. Plant Physiol. 2000, 27, 119–128. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Underhill, S.J.R. Expression of Gibberellin Metabolism Genes and Signalling Components in Dwarf Phenotype of Breadfruit (Artocarpus altilis) Plants Growing on Marang (Artocarpus odoratissimus) Rootstocks. Plants 2020, 9, 634. https://doi.org/10.3390/plants9050634
Zhou Y, Underhill SJR. Expression of Gibberellin Metabolism Genes and Signalling Components in Dwarf Phenotype of Breadfruit (Artocarpus altilis) Plants Growing on Marang (Artocarpus odoratissimus) Rootstocks. Plants. 2020; 9(5):634. https://doi.org/10.3390/plants9050634
Chicago/Turabian StyleZhou, Yuchan, and Steven J. R. Underhill. 2020. "Expression of Gibberellin Metabolism Genes and Signalling Components in Dwarf Phenotype of Breadfruit (Artocarpus altilis) Plants Growing on Marang (Artocarpus odoratissimus) Rootstocks" Plants 9, no. 5: 634. https://doi.org/10.3390/plants9050634
APA StyleZhou, Y., & Underhill, S. J. R. (2020). Expression of Gibberellin Metabolism Genes and Signalling Components in Dwarf Phenotype of Breadfruit (Artocarpus altilis) Plants Growing on Marang (Artocarpus odoratissimus) Rootstocks. Plants, 9(5), 634. https://doi.org/10.3390/plants9050634