Eco-Physiological Screening of Different Tomato Genotypes in Response to High Temperatures: A Combined Field-to-Laboratory Approach

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
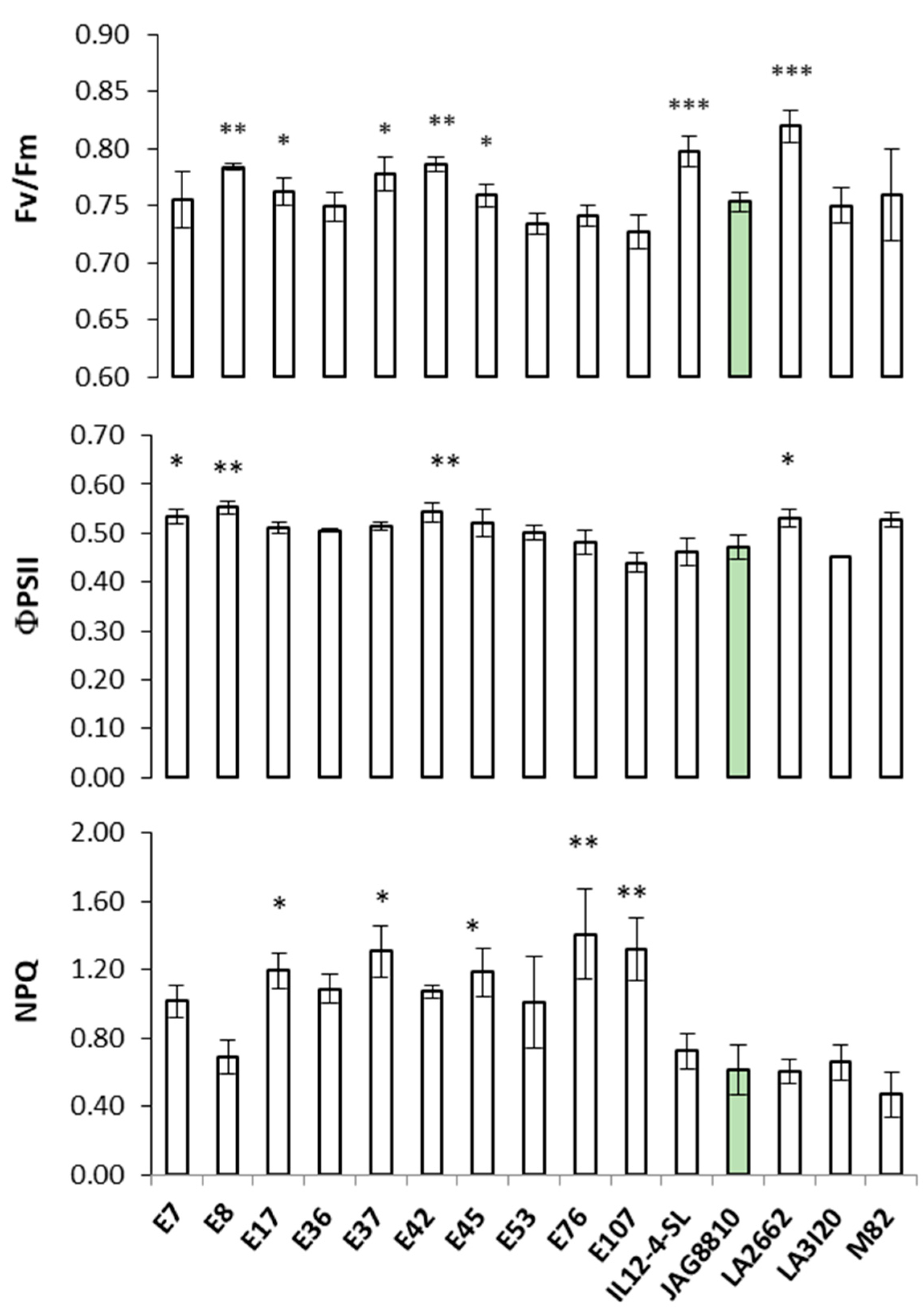
2.1. Greenhouse Trials: Correlations Between Physiological Parameters and Yields
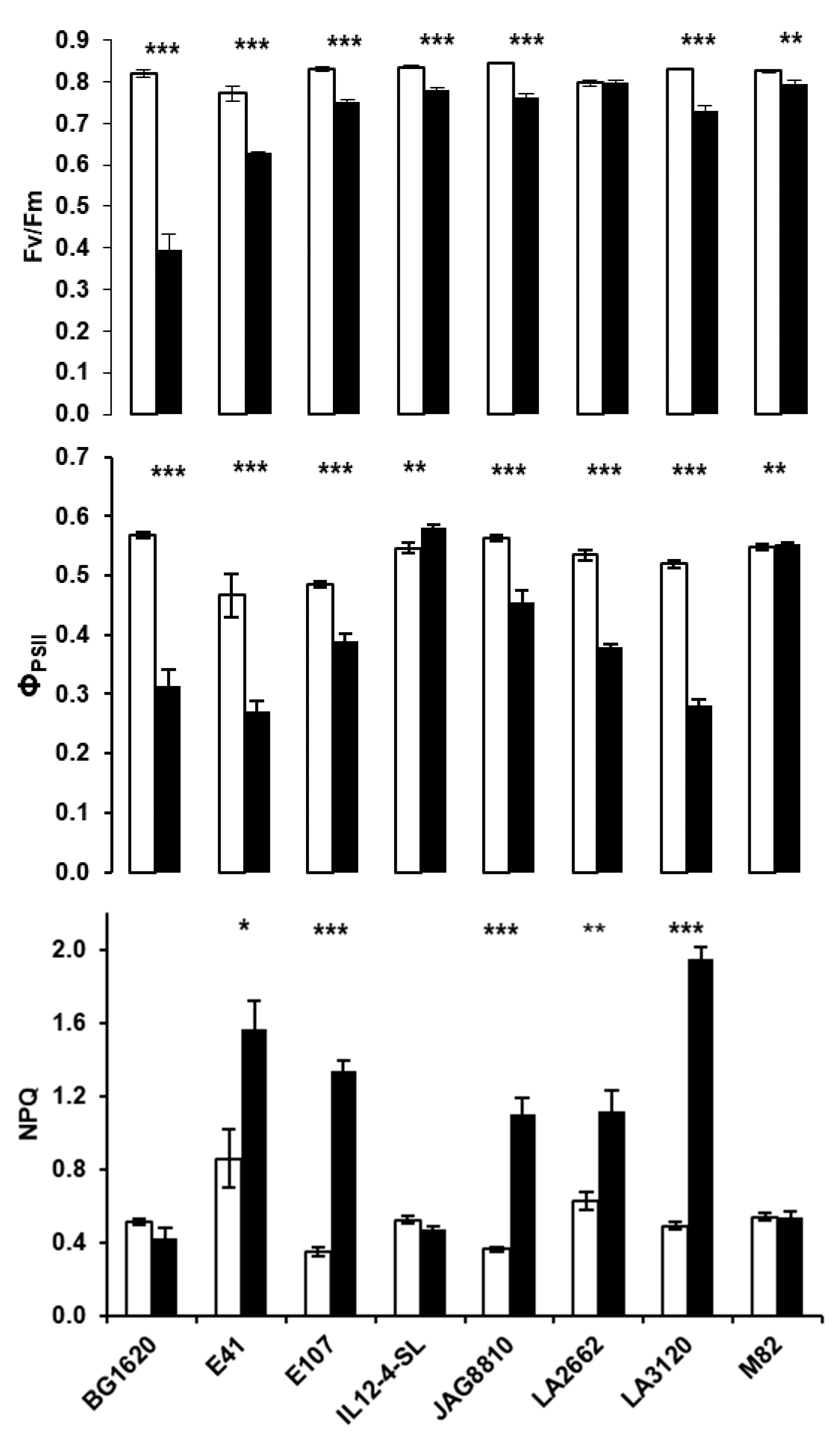
2.2. Chlorophyll a Fluorescence Measurements on Detached Leaves: Heat Shock Treatment at 35 °C and 45 °C
2.3. Heat-Induced Changes in Shape of Kautsky Kinetics
3. Discussion
4. Materials and Methods
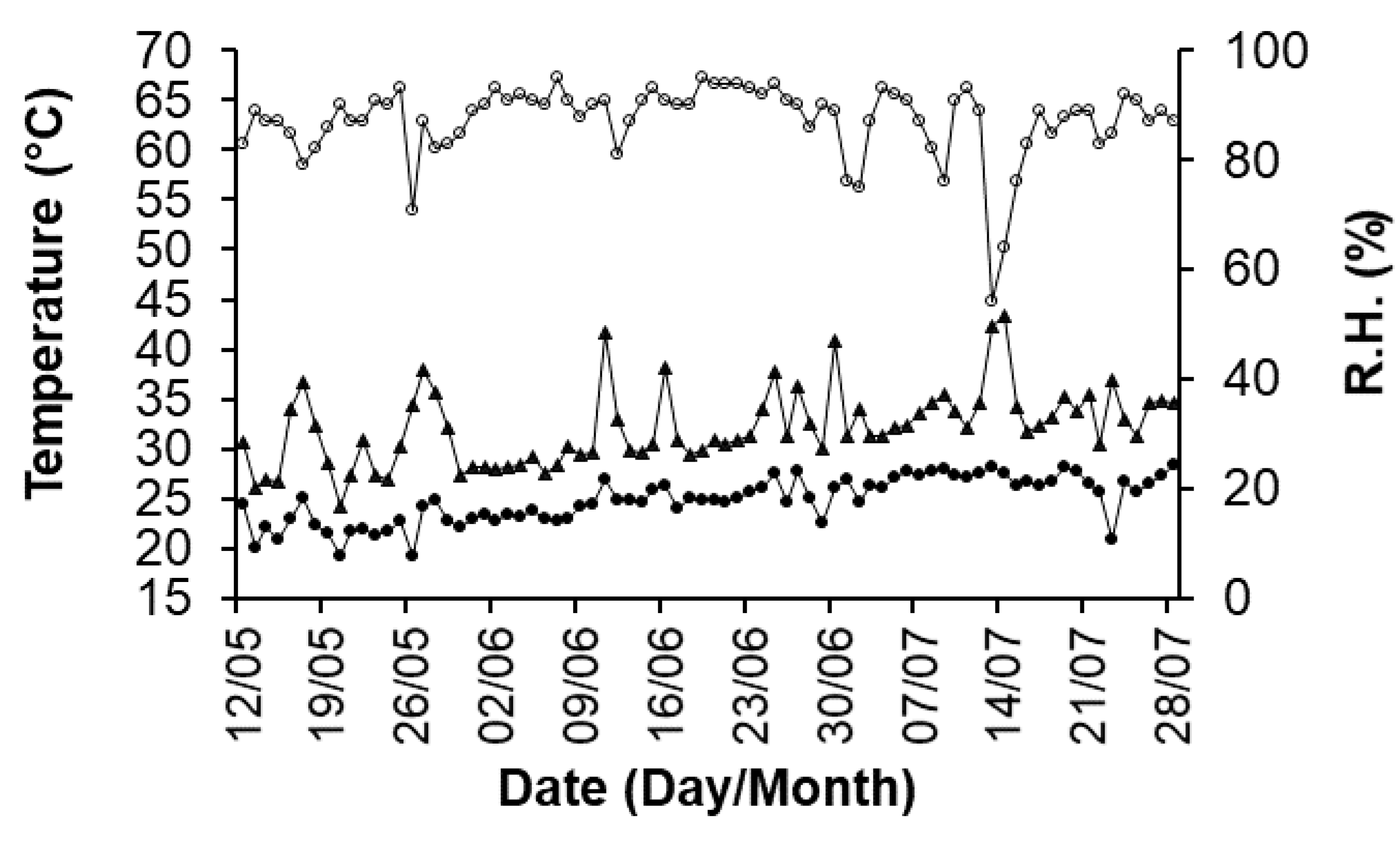
4.1. Plant Material and Growth Conditions
4.2. Functional Leaf Trait Analysis
4.3. Chlorophyll Fluorescence Emission Measurements in the Field
4.4. Determination of Total Chlorophylls and Carotenoids Content
4.5. Chlorophyll a Fluorescence—Heat Treatments and Laboratory Measurements
4.6. Analysis of Kautsky Kinetics Shape in Response to Heat Treatment
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M.M.B., Allen, S.K., Boschung, J., Nauels, A., Xia, W., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Vitale, L.; Arena, C.; Carillo, P.; Di Tommasi, P.; Mesolella, B.; Nacca, F.; De Santo, A.; Fuggi, A.; Magliulo, V. Gas exchange and leaf metabolism of irrigated maize at different growth stages. Plant. Biosyst. 2011, 145, 485–494. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Hanumanth Rao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant. Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wu, Z.; Wang, X.; Rosenqvist, E.; Wang, Y.; Zhao, T.; Ottosen, C.O. Evaluation of temperature stress tolerance in cultivated and wild tomatoes using photosynthesis and chlorophyll fluorescence. Hortic. Environ. Biotechnol. 2018, 59, 499–509. [Google Scholar] [CrossRef]
- Rigano, M.M.; Arena, C.; Di Matteo, A.; Sellitto, S.; Frusciante, L.; Barone, A. Eco-physiological response to water stress of drought-tolerant and drought-sensitive tomato genotypes. Plant. Biosyst. 2016, 150, 682–691. [Google Scholar] [CrossRef]
- Zhou, R.; Kjaer, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.O. Physiological response to heat stress during seedling and anthesis stage in tomato genotypes differing in heat tolerance. J. Agron. Crop. Sci. 2016, 203, 68–80. [Google Scholar] [CrossRef]
- Francesca, S.; Arena, C.; Hay-Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The use of a plant-based biostimulant improves plant performances and fruit quality in tomato plants grown at elevated temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect overheat stress on three tomato cultivars subjected to combined stress. BMC Plant. Biol. 2017, 17, 24. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Responses to high temperature stress. J. Photochem. Photobiol. B 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Coito, J.L.; Goncalves, E.F.; Chaves, M.M.; Amancio, S. Differential physiological response of the grapevine varieties Touriga Nacional and Tricadeira to combined heat, drought and light stresses. Plant. Biol. 2016, 18, 101–111. [Google Scholar] [CrossRef]
- Giorio, P.; Guida, G.; Mistretta, C.; Sellami, H.M.; Oliva, M.; Punzo, P.; Iovieno, P.; Arena, C.; De Maio, A.; Grillo, S.; et al. Physiological, biochemical and molecular responses to water stress and rehydration in Mediterranean adapted tomato landraces. Plant. Biol. 2018, 20, 995–1004. [Google Scholar] [CrossRef]
- Patanè, C.; Scordia, D.; Testa, G.; Cosentino, L.S. Physiological screening for drought tolerance in Mediterranean long-storage tomato. Plant. Sci. 2016, 249, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Brestič, M.; Živčák, M.; Kunderliková, K.; Allakhverdiev, S.I. High temperature specifically affects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 2016, 130, 251–266. [Google Scholar] [CrossRef]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C. Phenotyping from lab to field – tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant. Biol. 2019, 46, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yu, X.; Kjaer, K.H.; Rosenqvist, E.; Ottosen, C. Screening and validation of tomato genotypes under heat stress using Fv/Fm to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Virzo De Santo, A. Photosynthesis and photoprotective strategies in Laurus nobilis L. and Quercus ilex L. under summer drought and winter cold. Plant. Biosyst. 2008, 142, 472–479. [Google Scholar] [CrossRef]
- Tyystjärvi, E.; Koski, A.; Keränen, M.K.; Nevalainen, O. The Kautsky Curve Is a Built-in Barcode. Biophys. J. 1999, 77, 1159–1167. [Google Scholar] [CrossRef]
- Sharma, D.K.; Fernandez, J.O.; Rosenqvist, E.; Ottosen, C.; Andersen, S.B. Genotypic response of detached leaves versus intact plants for chlorophyll fluorescence parameters under high temperature stress in wheat. J. Plant. Physiol. 2014, 171, 576–586. [Google Scholar] [CrossRef]
- Prasanth, V.V.; Babu, M.S.; Basava, R.K.; Tripura Venkata, V.G.N.; Mangrauthia, S.K.; Voleti, S.R.; Neelamraju, S. Trait and marker associations in Oryza nivara and O. rufipogon derived rice lines under two different heat stress conditions. Front. Plant. Sci. 2017, 8, 1819. [Google Scholar] [CrossRef]
- Stirbet, A.; Riznichenko, G.; Rubin, A.B. Modeling chlorophyll a fluorescence transient: Relation to photosynthesis. Biochemistry 2014, 79, 291–323. [Google Scholar] [CrossRef]
- Vitale, L.; Arena, C.; Virzo De Santo, A.; D’Ambrosio, N. Effects of heat stress on gas exchange and Photosystem II (PSII) photochemical activity of Phillyrea angustifolia exposed to elevated CO2 and subsaturating irradiance. Botany 2008, 86, 435–441. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Hückstädt, A.B.; Suthaparan, A.; Mortensen, L.M.; Gislerød, H.R. The effect of low night and high day temperatures on photosynthesis in tomato. Am. J. Plant. Sci. 2013, 4, 2323–2331. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant. Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Heckathorn, S.A.; Mainali, K.; Tripathee, R. Timing effects of heat-stress on plant ecophysiological characteristics and growth. Front. Plant. Sci. 2016, 7, 1629. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Xu, W.; Liu, A.; Chen, S. COMT1 silencing aggravates heat stress-induced reduction in photosynthesis by decreasing chlorophyll content, photosystem II activity and electron transport efficiency in tomato. Front. Plant. Sci. 2018, 9, 998. [Google Scholar] [CrossRef]
- Pan, C.; Ahammed, G.J.; Li, X.; Shi, K. Elevated CO2 improves photosynthesis under high temperature by attenuating the functional limitations to energy fluxes, electron transport and redox homeostasis in tomato leaves. Front. Plant. Sci. 2018, 9, 1739. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant. Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Effects of high temperatures on the photosynthetic systems in spinach: Oxygen-evolving activities, fluorescence characteristics and the denaturation process. Photosynth. Res. 1998, 57, 51–59. [Google Scholar] [CrossRef]
- Yamashita, A.; Nijo, N.; Pospíšil, P.; Morita, N.; Takenaka, D.; Aminaka, R.; Yamamoto, Y. Quality control of Photosystem II—Reactive oxygen species are responsible for the damage to Photosystem II under moderate heat stress. J. Biol. Chem. 2008, 283, 28380–28391. [Google Scholar] [CrossRef]
- Wang, Q.L.; Chen, J.H.; He, N.Y.; Guo, F.Q. Metabolic reprogramming in chloroplasts under heat stress in plants. Int. J. Mol. Sci. 2018, 19, 849. [Google Scholar] [CrossRef] [PubMed]
- Willits, D.H.; Peet, M.M. Measurement of chlorophyll fluorescence as a heat stress indicator in tomato: Laboratory and greenhouse comparisons. J. Am. Soc. Hortic. Sci. 2001, 126, 188–194. [Google Scholar] [CrossRef]
- Riznichenko, G.; Lebedeva, G.; Pogosyan, S.; Sivchenko, M.; Rubin, A. Fluorescence induction curves registered from individual microalgae cenobiums in the process of population growth. Photosynt. Res. 1996, 49, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Marutani, Y.; Yamauchi, Y.; Miyoshi, A.; Inoue, K.; Ikeda, K.; Mizutani, M.; Sugimoto, Y. Regulation of photochemical energy transfer accompanied by structural changes in thylakoid membranes of heat-stressed wheat. Int. J. Mol. Sci. 2014, 15, 23042–23058. [Google Scholar] [CrossRef]
- Papageorgiou, G.C. The non-photochemical quenching of the electronically excited state of chlorophyll a in plants: Definitions, timelines, viewpoints, open questions. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria; Demmig, B.A., Garab, G., Adams, W., III, Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2014; pp. 1–44. [Google Scholar]
- Nesterenko, T.V.; Tikhomirov, A.A.; Shikhov, V.N. Influence of excitation light intensity and leaf age on the slow chlorophyll fluorescence transient in radish. Biophysics 2012, 57, 464–468. [Google Scholar] [CrossRef]
- Brestič, M.; Živčák, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.H.; Li, X.N.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef]
- Ruggieri, V.; Calafiore, R.; Schettini, C.; Rigano, M.M.; Olivieri, F.; Frusciante, L.; Barone, A. Exploiting genetic and genomic resources to enhance heat-tolerance in tomatoes. Agronomy 2019, 9, 22. [Google Scholar] [CrossRef]
- Tomato Genetic Resources Center. Available online: https://tgrc.ucdavis.edu/Data/Acc/AccSearch.aspx (accessed on 14 January 2020).
- Rigano, M.M.; Lionetti, V.; Raiola, A.; Bellincampi, D.; Barone, A. Pectic enzymes as potential enhancers of ascorbic acid production through the D-galacturonate pathway in Solanaceae. Plant. Sci. 2018, 266, 55–63. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen–Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1716. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, A.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. Handbook of protocols for standardized and easy measurement of plant functional traits worldwide. Aus. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Sorrentino, M.C.; Capozzi, F.; Amitrano, C.; Giordano, S.; Arena, C.; Spagnuolo, V. Performance of three cardoon cultivars in an industrial heavy metal-contaminated soil: Effects on morphology, cytology and photosynthesis. J. Hazard. Mater. 2018, 351, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Roháček, K.; Soukupová, J.; Barták, M. Chlorophyll fluorescence: A wonderful tool to study plant physiology and plant stress. In Plant Cell Compartments; Schoefs, B., Ed.; Research Signpost: Kerala, India, 2008; pp. 41–104. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | DW (g) | LA (cm2) | SLA (cm2 g−1) | Carotenoids (mg 100 g−1) | Chl a (mg 100 g−1) | Chl b (mg 100 g−1) | YP (kg pt−1) |
|---|---|---|---|---|---|---|---|
| E7 | 0.04 ± 0.10 | 11.41 ± 1.71 | 278.17 ± 63.62 * | 28.76 ± 0.07 *** | 104.82 ± 0.13 *** | 36.53 ± 0.29 *** | 0.98 ± 0.00 ** |
| E8 | 0.10 ± 0.04 | 14.37 ± 2.86 | 155.73 ± 36.29 | 41.30 ± 0.25 *** | 169.30 ± 0.27 *** | 76.05 ± 0.46 *** | 0.77 ± 0.17 *** |
| E17 | 0.08 ± 0.02 | 17.76 ± 2.37 | 231.60 ± 45.53 | 44.67 ± 0.23 *** | 177.92 ± 0.29 *** | 75.18 ± 0.77 *** | 1.26 ± 0.17 ** |
| E36 | 0.14 ± 0.03 *** | 20.09 ± 3.53 ** | 146.83 ± 15.50 | 39.94 ± 0.10 *** | 159.32 ± 0.37 *** | 67.97 ± 0.17 *** | 0.94 ± 0.22 ** |
| E37 | 0.15 ± 0.04 *** | 13.12 ± 2.52 | 92.48 ± 17.82** | 34.40 ± 0.03 *** | 133.61 ± 0.64 *** | 54.61 ± 0.64 *** | 0.88 ± 0.17 *** |
| E42 | 0.10 ± 0.03 | 18.02 ± 3.11 | 188.57 ± 37.70 | 43.02 ± 0.54 *** | 174.35 ± 1.62 *** | 76.43 ± 0.83 *** | 2.25 ± 1.04 |
| E45 | 0.16 ± 0.02 *** | 27.58 ± 4.78 *** | 171.16 ± 26.94 | 36.61 ± 0.04 * | 142.81 ± 0.36 *** | 59.02 ± 0.35 ** | 2.24 ± 1.04 |
| E53 | 0.07 ± 0.02 | 12.50 ± 2.19 | 210.79 ± 62.56 | 34.37 ± 0.06 *** | 132.38 ± 0.10 *** | 53.01 ± 0.26 *** | 1.32 ± 0.28 ** |
| E76 | 0.11 ± 0.01** | 21.17 ± 2.25 ** | 210.79 ± 62.56 | 44.63 ± 0.12 *** | 171.90 ± 0.06 *** | 68.26 ± 0.37 *** | 0.64 ± 0.01 *** |
| E107 | 0.07 ± 0.01 | 13.87 ± 2.95 | 194.36 ± 30.79 | 36.43 ± 0.04 ** | 134.20 ± 0.28 *** | 48.49 ± 0.44 *** | 0.42 ± 0.12 *** |
| IL12-4-SL | 0.10 ± 0.02 | 21.00 ± 1.83 ** | 217.16 ± 48.16 | 39.71 ± 0.07 *** | 149.58 ± 0.11 *** | 56.68 ± 0.20 *** | 2.77 ± 0.24 * |
| JAG8810 | 0.07 ± 0.01 | 13.46 ± 1.86 | 203.15 ± 55.39 | 42.96 ± 0.29 | 161.63 ± 1.51 | 60.49 ± 1.25 | 2.99 ± 0.5 |
| LA2662 | 0.12 ± 0.01 ** | 21.59 ± 2.07 ** | 190.13 ± 24.57 | 36.98 ± 0.03 *** | 143.90 ± 0.42 *** | 58.78 ± 0.47 ** | 1.97 ± 0.66 |
| LA3120 | 0.13 ± 0.03 * | 20.57 ± 1.14 *** | 166.53 ± 36.45 | 39.30 ± 0.13 *** | 155.43 ± 0.23 *** | 65.78 ± 0.60 *** | 1.91 ± 0.98 |
| M82 | 0.15 ± 0.03 *** | 24.37 ± 4.93 *** | 163.26 ± 12.14 | 32.87 ± 0.14 *** | 121.82 ± 0.23 *** | 44.88 ± 0.20 *** | 3.25 ± 0.6 * |
| YP | Fv/Fm | ΦPSII | NPQ | DW | SLA | LA | Chl a | Chl b | Car | |
|---|---|---|---|---|---|---|---|---|---|---|
| YP | 1 | 0.254 | −0.010 | −0.647 ** | 0.192 | 0.026 | 0.376 | −0.017 | 0.036 | −0.048 |
| Fv/Fm | 1 | 0.506 | −0.266 | 0.304 | −0.405 | 0.256 | 0.219 | 0.281 | 0.167 | |
| ΦPSII | 1 | −0.115 | 0.161 | −0.124 | 0.002 | −0.022 | 0.139 | −0.147 | ||
| NPQ | 1 | −0.068 | −0.049 | −0.183 | 0.106 | 0.035 | 0.154 | |||
| DW | 1 | −0.650 ** | 0.744 ** | 0.155 | 0.221 | 0.110 | ||||
| SLA | 1 | −0.149 | −0.226 | −0.307 | −0.167 | |||||
| LA | 1 | 0.251 | 0.212 | 0.281 | ||||||
| Chl a | 1 | 0.970 ** | 0.983 ** | |||||||
| Chl b | 1 | 0.909 ** | ||||||||
| Car | 1 |
| Control | ||||||||
| BG1620 | E41 | E107 | IL12-4-SL | JAG8810 | LA2662 | LA3120 | M82 | |
| O/P | 0.229 | 0.242 | 0.212 | 0.220 | 0.201 | 0.239 | 0.194 | 0.207 |
| P/S | 1.810 | 1.167 | 1.065 | 1.517 | 1.405 | 1.242 | 1.193 | 1.673 |
| P/M | 1.669 | - | 1.059 | 1.389 | 1.318 | - | 1.163 | 1.486 |
| S/M | 0.922 | - | 0.994 | 0.916 | 0.938 | - | 0.975 | 0.888 |
| P/T (steady state) | 2.665 | 3.101 | 2.101 | 2.482 | 2.428 | 2.887 | 2.694 | 2.801 |
| M/T (steady state) | 1.597 | - | 1.985 | 1.787 | 1.842 | - | 2.316 | 1.886 |
| Rfd = (Fp−Fs)/Fs | 1.665 | 2.101 | 1.101 | 1.482 | 1.428 | 1.887 | 1.694 | 1.801 |
| tP | 1.120 | 1.120 | 0.960 | 1.200 | 1.040 | 1.040 | 1.280 | 1.040 |
| tS | 6.080 | 3.360 | 4.080 | 4.080 | 4.080 | 4.080 | −29.940 | 6.080 |
| tM | 10.080 | - | 2.720 | 10.080 | 8.080 | - | 6.080 | 12.080 |
| tT | 270.060 | 270.060 | 270.060 | 270.060 | 270.060 | 270.060 | 270.060 | 270.060 |
| Dip at | 26.080 | - | 52.080 | 42.080 | - | 48.080 | 86.080 | 38.080 |
| Heat Treated (60 min at 45 °C) | ||||||||
| BG1620 | E41 | E107 | IL12-4-SL | JAG8810 | LA2662 | LA3120 | M82 | |
| O/P | 0.713 *** | 0.483 ** | 0.341 ** | 0.344 ** | 0.366 ** | 0.252 * | 0.379 ** | 0.316 ** |
| P/S | - | - | - | 1.280 * | 0.936 * | - | - | 0.855 * |
| P/M | - | - | - | 1.166 * | 0.929 * | - | - | 0.853 * |
| S/M | - | - | - | 0.911 * | 0.993 * | - | - | 0.997 * |
| P/T (steady state) | 1.658 * | 2.729 * | 2.680 * | 2.250 * | 2.407 * | 2.779 * | 2.881 * | 2.302 * |
| M/T (steady state) | - | - | - | 1.929 * | 2.591 * | - | - | 2.700 * |
| Rfd = (Fp-Fs)/Fs | 0.658 ** | 1.729 * | 1.680 ** | 1.250 * | 1.407 * | 1.779 * | 1.881 * | 1.302 * |
| tP | 1.280 * | 1.280 * | 1.760 ** | 1.120 * | 1.280 * | 1.040 | 1.280 | 1.040 |
| tS | - | - | - | 4.080 | 2.240 *** | - | - | 3.360 |
| tM | - | - | - | 8.080 | 2.400 *** | - | - | 4.080 |
| tT | 255.060 * | 270.060 * | 245.060 * | 260.060 * | 245.060 * | 260.060 * | 270.060 * | 250.06 * |
| Dip at | - | - | - | 34.080 * | 40.080 | - | - | 40.08 * |
| No. | Genotype | Origin | Common Name |
|---|---|---|---|
| 1 | E7 | Italy | Corbarino PC04 |
| 2 | E8 | Italy | Corbarino PC05 |
| 3 | E17 | Italy | Pantano Romanesco |
| 4 | E36 | Italy | Riccia San Vito |
| 5 | E37 | Italy | Siccagno |
| 6 | E41 | Italy | Parmitanella |
| 7 | E42 | Italy | PI15250 |
| 8 | E45 | Italy | SM246 |
| 9 | E53 | South America | Latin American cultivar (Honduras) |
| 10 | E76 | URSS | Black Plum |
| 11 | E107 | Europe | E-L-19, Spain |
| 12 | JAG8810 | - | Monsanto F1 hybrid |
| 13 | M82 | California | M82 |
| 14 | IL12-4-SL | Italy | IL12-4-SL |
| 15 | LA2662 | - | Saladette |
| 16 | LA3120 | - | Malintka |
| 17 | BG1620 | Bulgary | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arena, C.; Conti, S.; Francesca, S.; Melchionna, G.; Hájek, J.; Barták, M.; Barone, A.; Rigano, M.M. Eco-Physiological Screening of Different Tomato Genotypes in Response to High Temperatures: A Combined Field-to-Laboratory Approach. Plants 2020, 9, 508. https://doi.org/10.3390/plants9040508
Arena C, Conti S, Francesca S, Melchionna G, Hájek J, Barták M, Barone A, Rigano MM. Eco-Physiological Screening of Different Tomato Genotypes in Response to High Temperatures: A Combined Field-to-Laboratory Approach. Plants. 2020; 9(4):508. https://doi.org/10.3390/plants9040508
Chicago/Turabian StyleArena, Carmen, Stefano Conti, Silvana Francesca, Giuseppe Melchionna, Josef Hájek, Miloš Barták, Amalia Barone, and Maria Manuela Rigano. 2020. "Eco-Physiological Screening of Different Tomato Genotypes in Response to High Temperatures: A Combined Field-to-Laboratory Approach" Plants 9, no. 4: 508. https://doi.org/10.3390/plants9040508
APA StyleArena, C., Conti, S., Francesca, S., Melchionna, G., Hájek, J., Barták, M., Barone, A., & Rigano, M. M. (2020). Eco-Physiological Screening of Different Tomato Genotypes in Response to High Temperatures: A Combined Field-to-Laboratory Approach. Plants, 9(4), 508. https://doi.org/10.3390/plants9040508