Heterologous Expression of Three Ammopiptanthus mongolicus Dehydrin Genes Confers Abiotic Stress Tolerance in Arabidopsis thaliana


Abstract
1. Introduction
2. Results
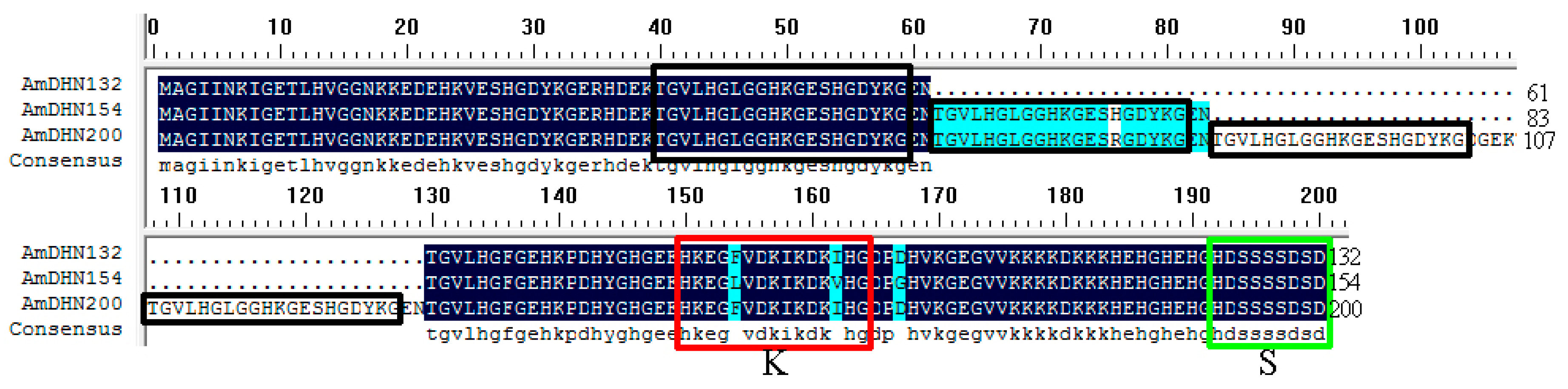
2.1. Sequence Characterization of the Dehydrin Gene Family
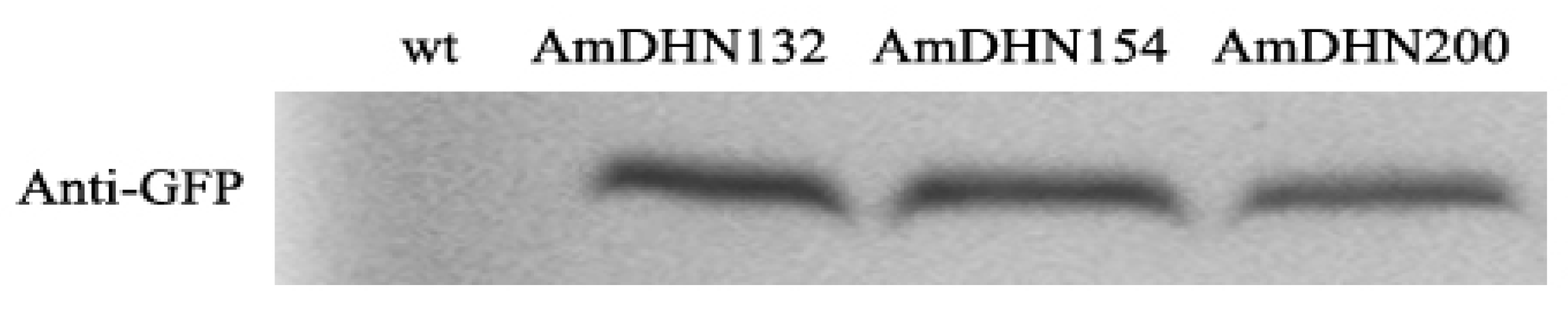
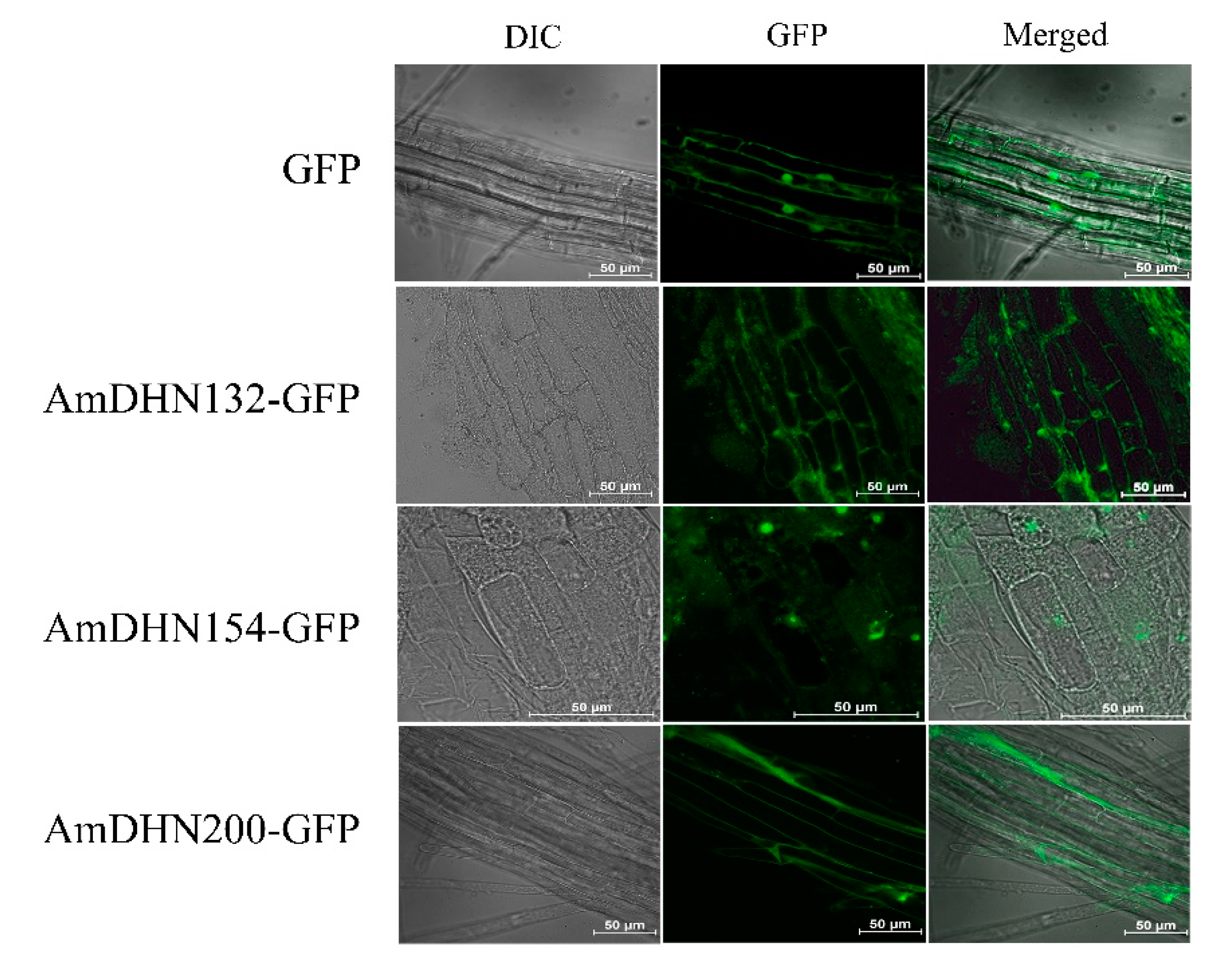
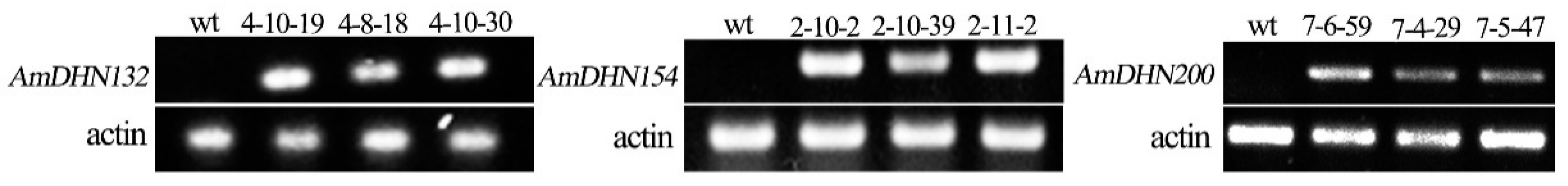
2.2. Creation of Transgenic Arabidopsis and the Subcellular Localization of AmDHNs
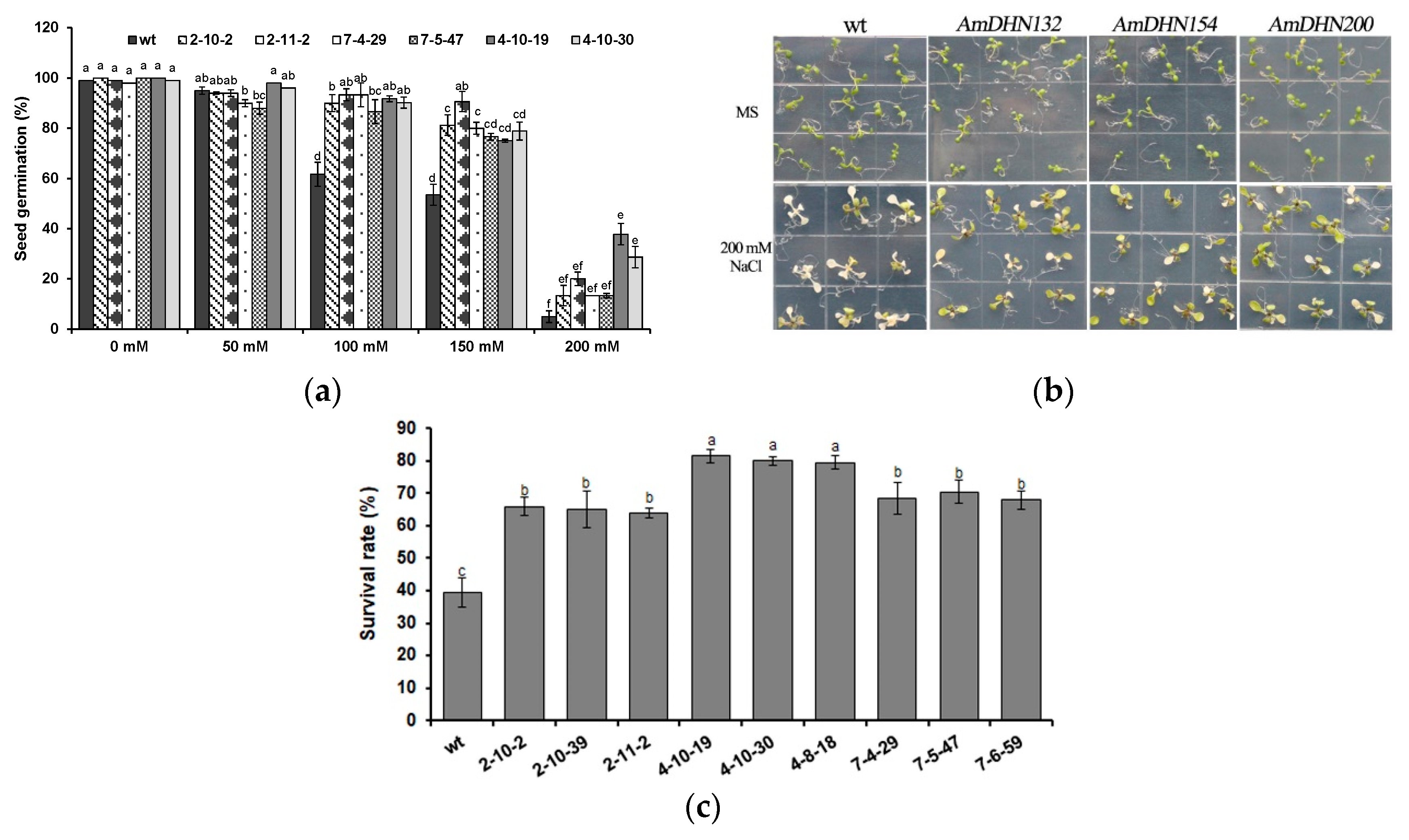
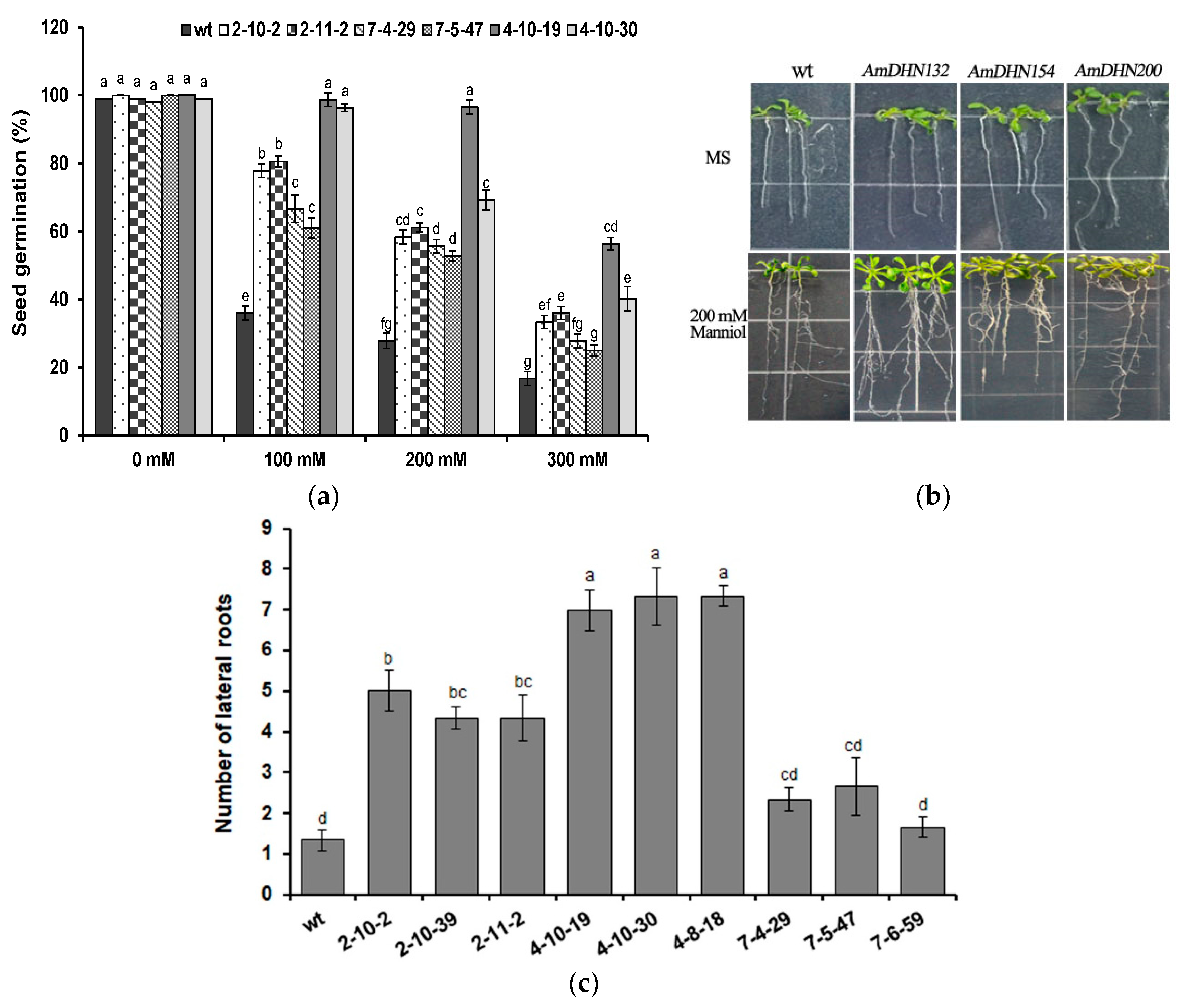
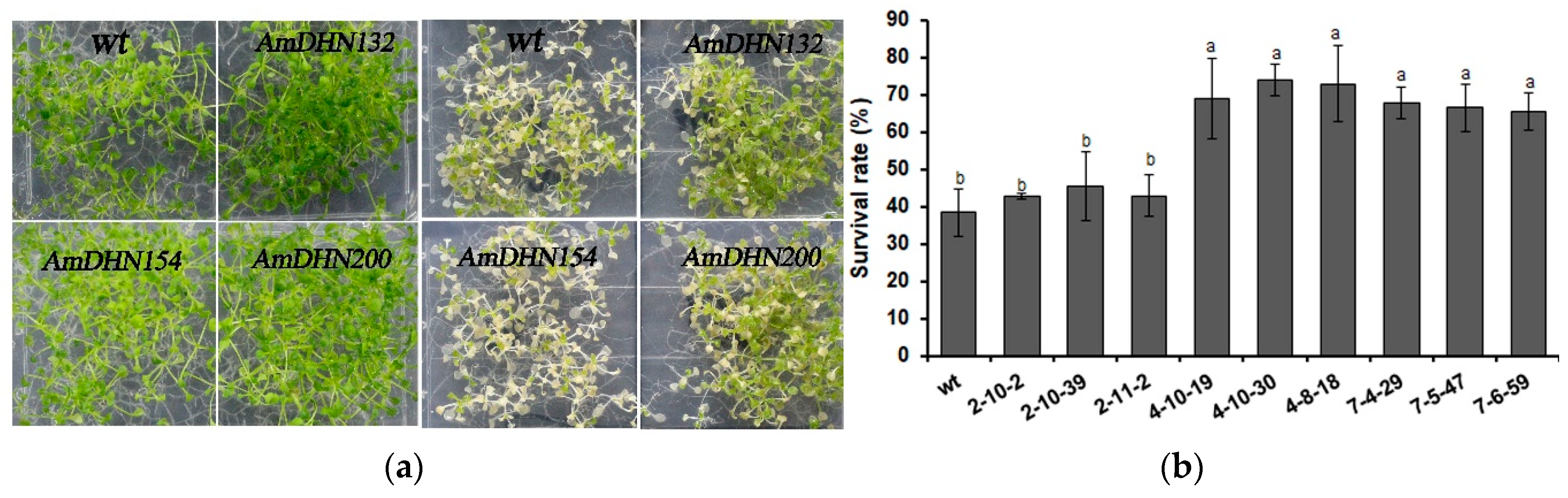
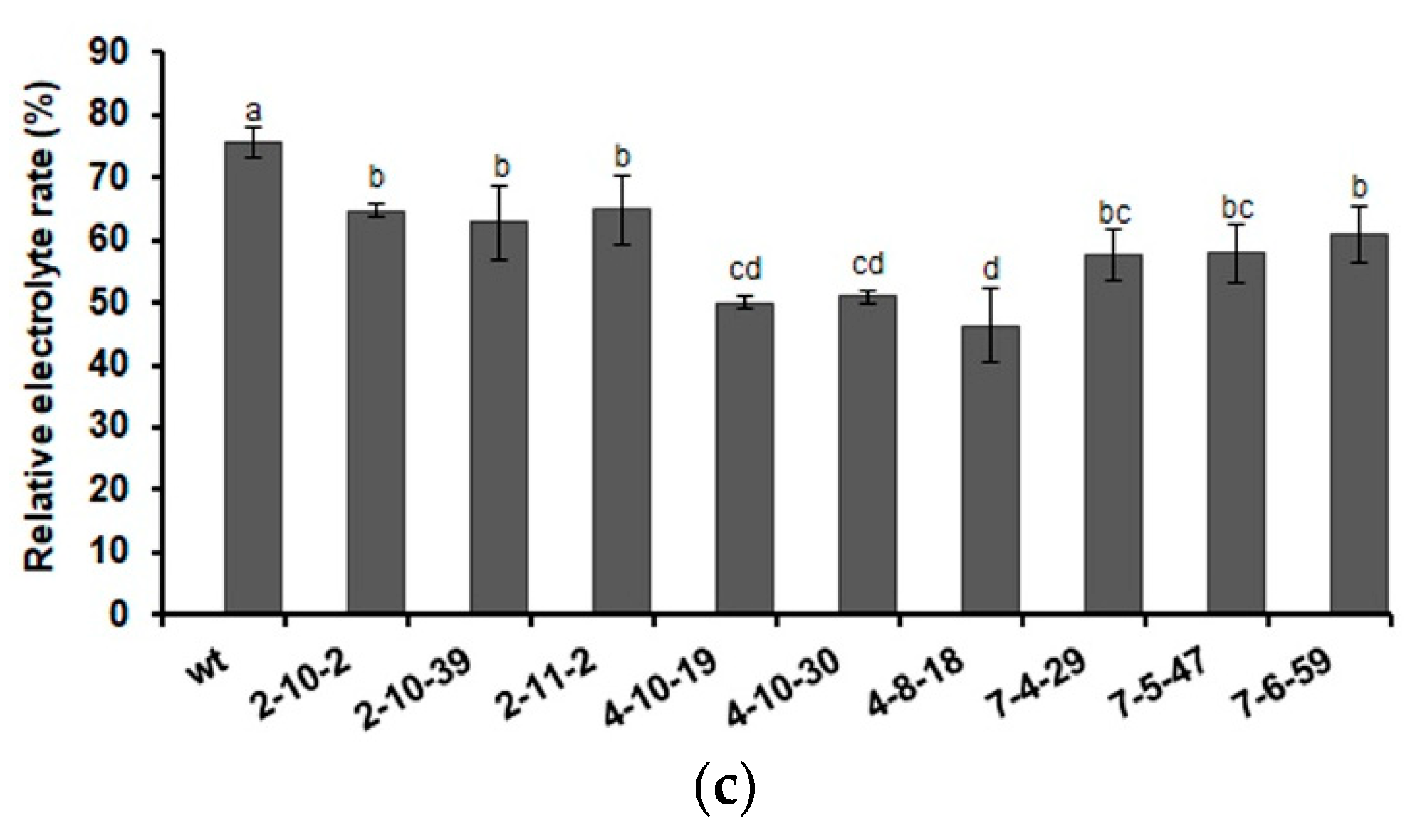
2.3. Heterologous Expression of AmDHNs in Arabidopsis Enhances Abiotic Stress Tolerance
3. Discussion
3.1. AmDHNs Are Conserved in A. mongolicus
3.2. Overexpression of AmDHNs Increases Abiotic Stress Tolerance of Arabidopsis
3.3. Potential Use of AmDHNs and Further Research
4. Materials and Methods
4.1. Plant Material and Normal and Stress Growth Conditions
4.2. Cloning A. mongolicus Dehydrin Genes
4.3. Construction and Transformation of Eukaryotic Expression Vectors
4.4. Screening of Homozygous Lines
4.5. RT-PCR Analysis
4.6. Western Blot Analysis
4.7. Subcellular Localization of AmDHN
4.8. Abiotic Stress Tolerance Analysis
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karlson, D.T.; Fujino, T.; Kimura, S.; Baba, K.; Itoh, T.; Ashworth, E.N. Novel plasmodesmata association of dehydrin-like proteins in cold-acclimated Red-osier dogwood (Cornussericea). Tree physiol. 2003, 23, 759–767. [Google Scholar] [CrossRef][Green Version]
- Ismail, F.A.; Nitsch, L.M.; Wolters-Arts, M.M.; Mariani, C.; Derksen, J.W. Semi-viviparous embryo development and dehydrin expression in the mangrove Rhizophora mucronata Lam. Sex. Plant Reprod. 2010, 23, 95–103. [Google Scholar] [CrossRef][Green Version]
- Liu, Y.; Song, Q.; Li, D.; Yang, X.; Li, D. Multifunctional roles of plant dehydrins in response to environmental stresses. Front. Plant Sci. 2017, 8, 1018. [Google Scholar] [CrossRef]
- Qiu, H.; Zhang, L.; Liu, C.; He, L.; Wang, A.; Liu, H.L.; Zhu, J.B. Cloning and characterization of a novel dehydrin gene, SiDhn2, from Saussurea involucrata Kar et Kir. Plant Mol. Biol. 2014, 84, 707–718. [Google Scholar] [CrossRef]
- Remus-Borel, W.; Castonguay, Y.; Cloutier, J.; Michaud, R.; Bertrand, A.; Desgagnes, R.; Laberge, S. Dehydrin variants associated with superior freezing tolerance in alfalfa (Medicago sativa L.). Theor. Appl. Genet. 2010, 120, 1163–1174. [Google Scholar] [CrossRef]
- Shi, J.; Liu, M.; Chen, Y.; Wang, J.; Lu, C. Heterologous expression of the dehydrin-like protein gene AmDHN from Ammopiptanthus mongolicus enhances viability of Escherichia coli and tobacco under cold stress. Plant Growth Regul. 2016, 79, 71–80. [Google Scholar] [CrossRef]
- Brini, F.; Hanin, M.; Lumbreras, V.; Amara, I.; Khoudi, H.; Hassairi, A.; Pages, M.; Masmoudi, K. Overexpression of wheat dehydrin DHN-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 2007, 26, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, M.; Zhu, Z.; Li, S.; Xu, Y.; Zhang, C.; Singer, S.D.; Wang, Y. Identification of the dehydrin gene family from grapevine species and analysis of their responsiveness to various forms of abiotic and biotic stress. BMC Plant Biol. 2012, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, S.; Jiang, C.; Wang, Y.; Lv, B.; Shen, J.; Ming, F. RcLEA, a late embryogenesis abundant protein gene isolated from Rosa chinensis, confers tolerance to Escherichia coli and Arabidopsis thaliana and stabilizes enzyme activity under diverse stresses. Plant Mol. Biol. 2014, 85, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Dure, L.; Crouch, M.; Harada, J.; Ho, T.H.D.; Mundy, J.; Quatrano, R.; Thomas, T.; Sung, Z.R. Common amino acid sequence domains among the LEA proteins of higher plants. Plant Mol. Biol. 1989, 12, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Szalaine-Agoston, B.; Kovacs, D.; Tompa, P.; Perczel, A. Full backbone assignment and dynamics of the intrinsically disordered dehydrin ERD14. Biomol. NMR Assign. 2011, 5, 189–193. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plant. 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Amara, I.; Zaidi, I.; Masmoudi, K.; Ludevid, M.D.; Pagès, M.; Goday, A.; Brini, F. Insights into late embryogenesis abundant (LEA) proteins in plants: From Structure to the Functions. Am. J. Plant Sci. 2014, 5, 3440–3455. [Google Scholar] [CrossRef]
- Hughes, S.L.; Schart, V.; Malcolmson, J.; Hogarth, K.A.; Martynowicz, D.M.; Tralman-Baker, E.; Patel, S.N.; Graether, S.P. The importance of size and disorder in the cryoprotective effects of dehydrins. Plant Physiol. 2013, 163, 1376–1386. [Google Scholar] [CrossRef]
- Perdiguero, P.; Collada, C.; Soto, A. Novel dehydrins lacking complete K-segments in Pinaceae. The exception rather than the rule. Front. Plant Sci. 2014, 5, 682. [Google Scholar] [CrossRef]
- Bao, F.; Du, D.; An, Y.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Overexpression of Prunus mume dehydrin genes in tobacco enhances tolerance to cold and drought. Front. Plant Sci. 2017, 8, 151. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiol. Plant. 1997, 100, 291–296. [Google Scholar] [CrossRef]
- Ochoa-Alfaro, A.E.; Rodriguez-Kessler, M.; Perez-Morales, M.B.; Delgado-Sanchez, P.; Cuevas-Velazquez, C.L.; Gomez-Anduro, G.; Jimenez-Bremont, J.F. Functional characterization of an acidic SK(3) dehydrin isolated from an Opuntia streptacantha cDNA library. Planta 2012, 235, 565–578. [Google Scholar] [CrossRef]
- Chiappetta, A.; Muto, A.; Bruno, L.; Woloszynska, M.; Van-Lijsebettens, M.; Bitonti, M.B. A dehydrin gene isolated from feral olive enhances drought tolerance in Arabidopsis transgenic plants. Front. Plant Sci. 2015, 6, 392. [Google Scholar] [CrossRef] [PubMed]
- Saibi, W.; Zouari, N.; Masmoudi, K.; Brini, F. Role of the durum wheat dehydrin in the function of proteases conferring salinity tolerance in Arabidopsis thaliana transgenic lines. Int. J. Biol. Macromol. 2016, 85, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Reyes, J.L.; Wei, H.; Yang, Y.; Karlson, D.; Covarrubias, A.A.; Krebs, S.L.; Fessehaie, A.; Arora, R. RcDhn5, a cold acclimation-responsive dehydrin from Rhododendron catawbiense rescues enzyme activity from dehydration effects in vitro and enhances freezing tolerance in RcDhn5—Overexpressing Arabidopsis plants. Physiol. Plant. 2008, 134, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J.; Lammers, P.J. An osmotic stress protein of cyanobacteria is immunologically related to plant dehydrins. Plant. Physiol. 1993, 101, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, L.; Svensson, J.; Carballo, V.; Izmendi, D.; Welin, B.; Vidal, S. A dehydrin gene in Physcomitrella patens is required for salt and osmotic stress tolerance. Plant J. 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Pochon, S.; Simoneau, P.; Pigné, S.; Balidas, S.; Bataillé-Simoneau, N.; Campion, C.; Jaspard, E.; Calmes, B.; Hamon, B.; Berruyer, R.; et al. Dehydrin-like proteins in the necrotrophic fungus Alternaria brassicicola have a role in plant pathogenesis and stress response. PLoS ONE 2013, 8, e75143. [Google Scholar] [CrossRef]
- Guo, L.; Yu, Y.; Xia, X.; Yin, W. Identification and functional characterisation of the promoter of the calcium sensor gene CBL1 from the xerophyte Ammopiptanthus mongolicus. BMC Plant Biol. 2010, 10, 18. [Google Scholar] [CrossRef]
- Liu, M.; Shi, J.; Lu, C. Identification of stress-responsive genes in Ammopiptanthus mongolicus using ESTs generated from cold and drought-stressed seedlings. BMC Plant Biol. 2013, 13, 88. [Google Scholar] [CrossRef]
- Liu, M.; Lu, C.; Shen, X.; Yin, W. Characterization and function analysis of a cold-induced AmCIP gene encoding a dehydrin-like protein in Ammopiptanthus mongolicus. DNA Seq. 2006, 17, 342–349. [Google Scholar] [CrossRef]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef]
- Zhou, Y.; Hu, L.; Xu, S.; Jiang, L.; Liu, S. Identification and transcriptional Sequence analysis of dehydrin gene family in cucumber (Cucumis sativus). Acta Physiol. Plant. 2018, 40, 144. [Google Scholar] [CrossRef]
- Hara, M.; Monnaa, S.; Murataa, T.; Nakanoa, T.; Amano, S.; Nachbarb, M.; Wätzigb, H. The Arabidopsis KS-type dehydrin recovers lactate dehydrogenase activity inhibited by copper with the contribution of His residues. Plant Sci. 2016, 245, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Niu, Q.; Qian, M.; Bai, S.; Teng, Y. Genome-wide identification, characterization, and expression analysis of the dehydrin gene family in Asian pear (Pyrus pyrifolia). Tree Genet. Gen 2015, 11, 110. [Google Scholar] [CrossRef]
- Tanmoy, H.; Gouranga, U.; Sudipta, R. YSK2 type dehydrin (SbDhn1) from Sorghum bicolor showed improved protection under high temperature and osmotic stress condition. Front. Plant Sci. 2017, 8, 918. [Google Scholar]
- Shekhawat, U.K.S.; Srinivas, L.; Ganapathi, T.R. MusaDHN-1, a novel multiple stress-inducible SK3-type dehydrin gene, contributes affirmatively to drought- and salt-stress tolerance in banana. Planta 2011, 234, 915. [Google Scholar] [CrossRef]
- Kumar, M.; Lee, S.C.; Kim, J.Y.; Kim, S.J.; Kim, S.R. Over-expression of dehydrin gene, OsDhn1, improves drought and salt stress tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). J. Plant Biol. 2014, 57, 383–393. [Google Scholar] [CrossRef]
- Falavigna, V.D.; Miotto, Y.E.; Porto, D.D.; Anzanello, R.; Santos, H.P.; Bello, F.F.; Margispinheiro, M.; Pasquali, G.; Revers, L.F. Functional diversification of the dehydrin gene family in apple and its contribution to cold acclimation during dormancy. Physiol. Plant. 2015, 155, 315–329. [Google Scholar] [CrossRef]
- Rorat, T. Pland dehydrins-tissue location, structure and function. Cell Mol. Biol. Lett. 2006, 11, 536–556. [Google Scholar] [CrossRef]
- Puhakainen, T.; Hess, M.W.; Makela, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol. 2004, 54, 743–753. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, L.; Zhu, J.; Liu, H.; Wang, A. Cloning and characterization of SiDHN, a novel dehydrin gene from Saussurea involucrata Kar. et Kir. that enhances cold and drought tolerance in tobacco. Plant Sci. 2016, 256, 160. [Google Scholar] [CrossRef]
- Ruibal, C.; Salamo, I.P.; Carballo, V.; Castro, A.; Bentancor, M.; Borsani, O.; Szabados, L.; Vidal, S. Differential contribution of individual dehydrin genes from Physcomitrella patens to salt and osmotic stress tolerance. Plant Sci. 2012, 190, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, T.; Upadhyaya, G.; Halder, T.; Mukherjee, A.; Majumder, A.L.; Ray, S. Different dehydrins perform separate functions in Physcomitrella patens. Planta 2016, 245, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fan, P.; Song, H.; Chen, X.; Li, X.; Li, Y. Comparative proteomic analysis of differentially expressed proteins in shoots of Salicornia europaea under different salinity. J. Proteome Res. 2009, 8, 3331–3345. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Size (bp) | Protein Length (aa) | Molecular Weight (kD) | Gene Accession Number |
|---|---|---|---|---|
| AmDHN132 | 399 | 132 | 14.41 | MH512938 |
| AmDHN154 | 465 | 154 | 16.65 | MH512939 |
| AmDHN200 | 603 | 200 | 21.42 | MH512941 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, H.; Wang, Y.; Yu, T.; Chen, S.; Chen, Y.; Lu, C. Heterologous Expression of Three Ammopiptanthus mongolicus Dehydrin Genes Confers Abiotic Stress Tolerance in Arabidopsis thaliana. Plants 2020, 9, 193. https://doi.org/10.3390/plants9020193
Cui H, Wang Y, Yu T, Chen S, Chen Y, Lu C. Heterologous Expression of Three Ammopiptanthus mongolicus Dehydrin Genes Confers Abiotic Stress Tolerance in Arabidopsis thaliana. Plants. 2020; 9(2):193. https://doi.org/10.3390/plants9020193
Chicago/Turabian StyleCui, Hongwei, Yang Wang, Tingqiao Yu, Shaoliang Chen, Yuzhen Chen, and Cunfu Lu. 2020. "Heterologous Expression of Three Ammopiptanthus mongolicus Dehydrin Genes Confers Abiotic Stress Tolerance in Arabidopsis thaliana" Plants 9, no. 2: 193. https://doi.org/10.3390/plants9020193
APA StyleCui, H., Wang, Y., Yu, T., Chen, S., Chen, Y., & Lu, C. (2020). Heterologous Expression of Three Ammopiptanthus mongolicus Dehydrin Genes Confers Abiotic Stress Tolerance in Arabidopsis thaliana. Plants, 9(2), 193. https://doi.org/10.3390/plants9020193