Effect of Cadmium Toxicity on Growth, Oxidative Damage, Antioxidant Defense System and Cadmium Accumulation in Two Sorghum Cultivars

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Planting Material and Treatments
2.2. Measurement of Growth Parameters
2.3. Estimation of Cd Accumulation
2.4. Determination of Cell Membrane Stability and Oxidative Damage
2.5. Estimation of Antioxidant Enzyme Activity
2.6. Statistical Analysis
3. Results
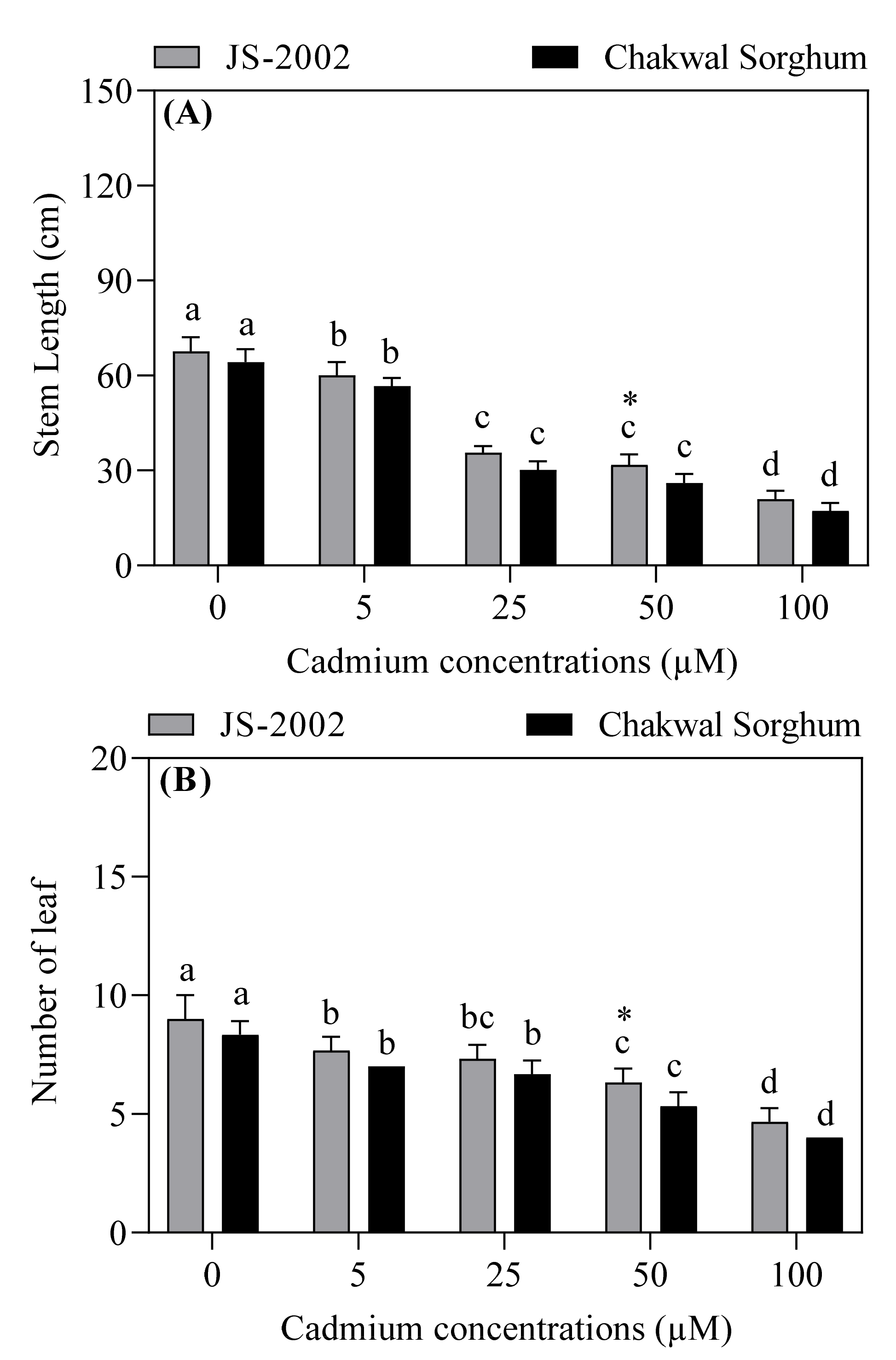
3.1. Effects of Exogenous Cadmium on Growth Parameters
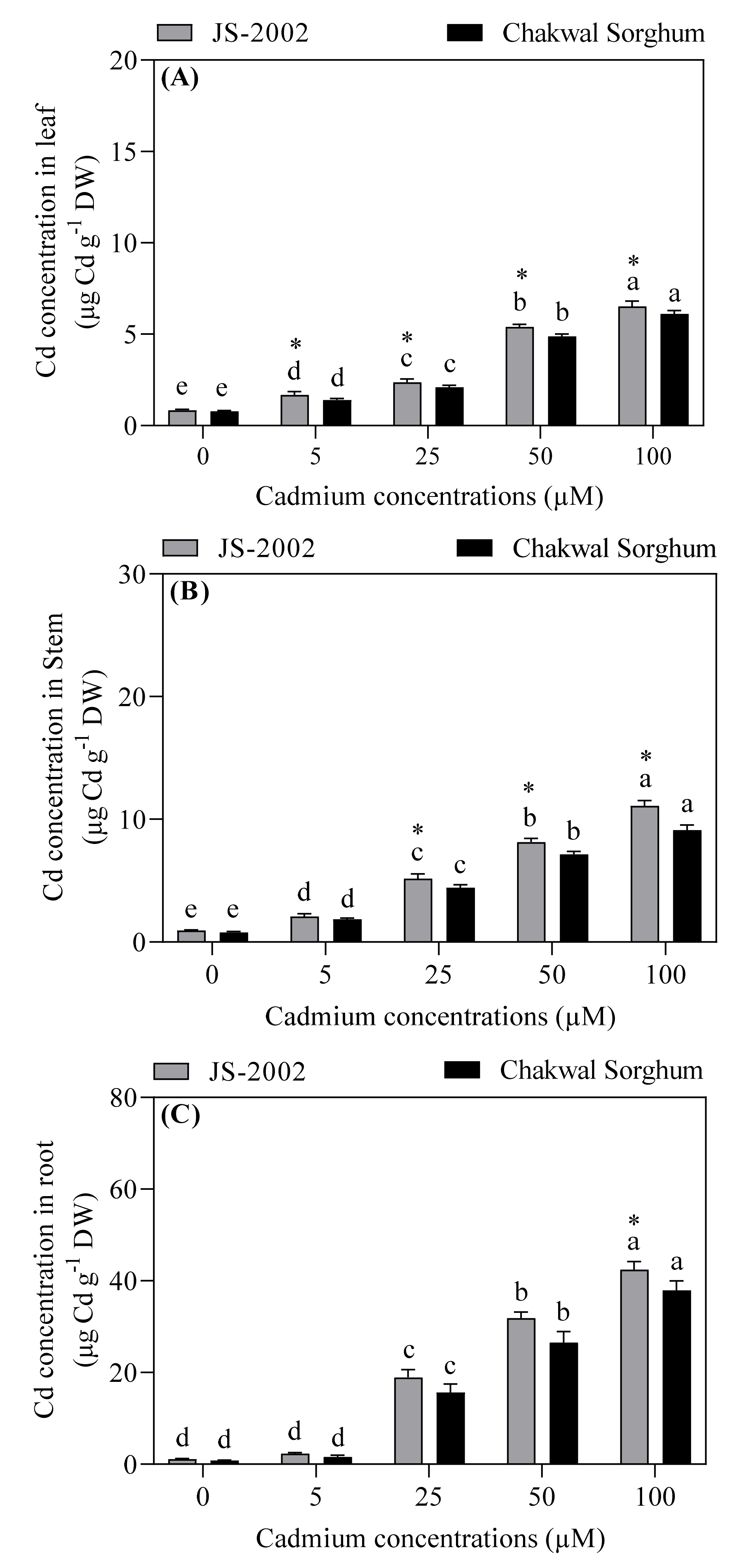
3.2. Effects of Exogenous Cadmium on Cadmium Accumulation
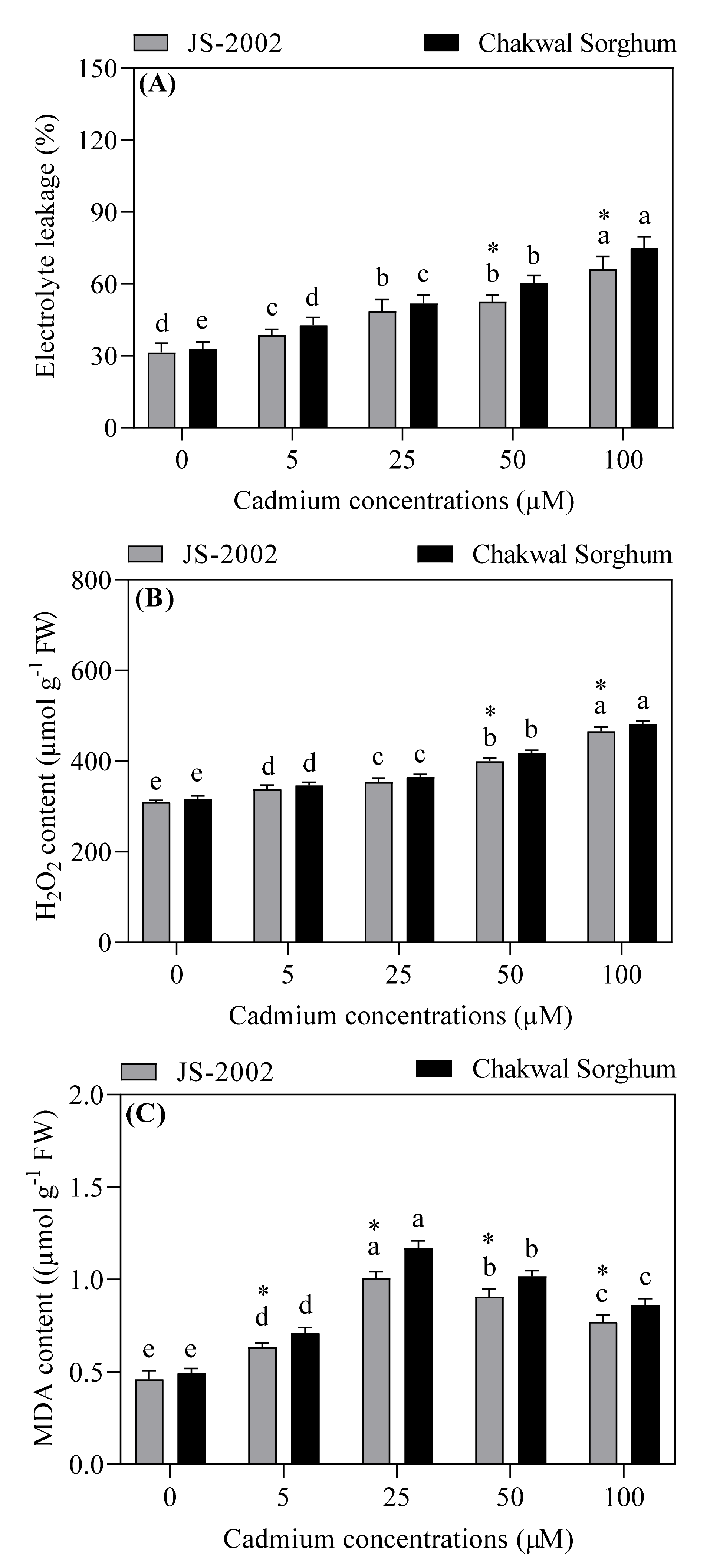
3.3. Effects of Exogenous Cadmium on Cell Membrane Stability and Oxidative Damage
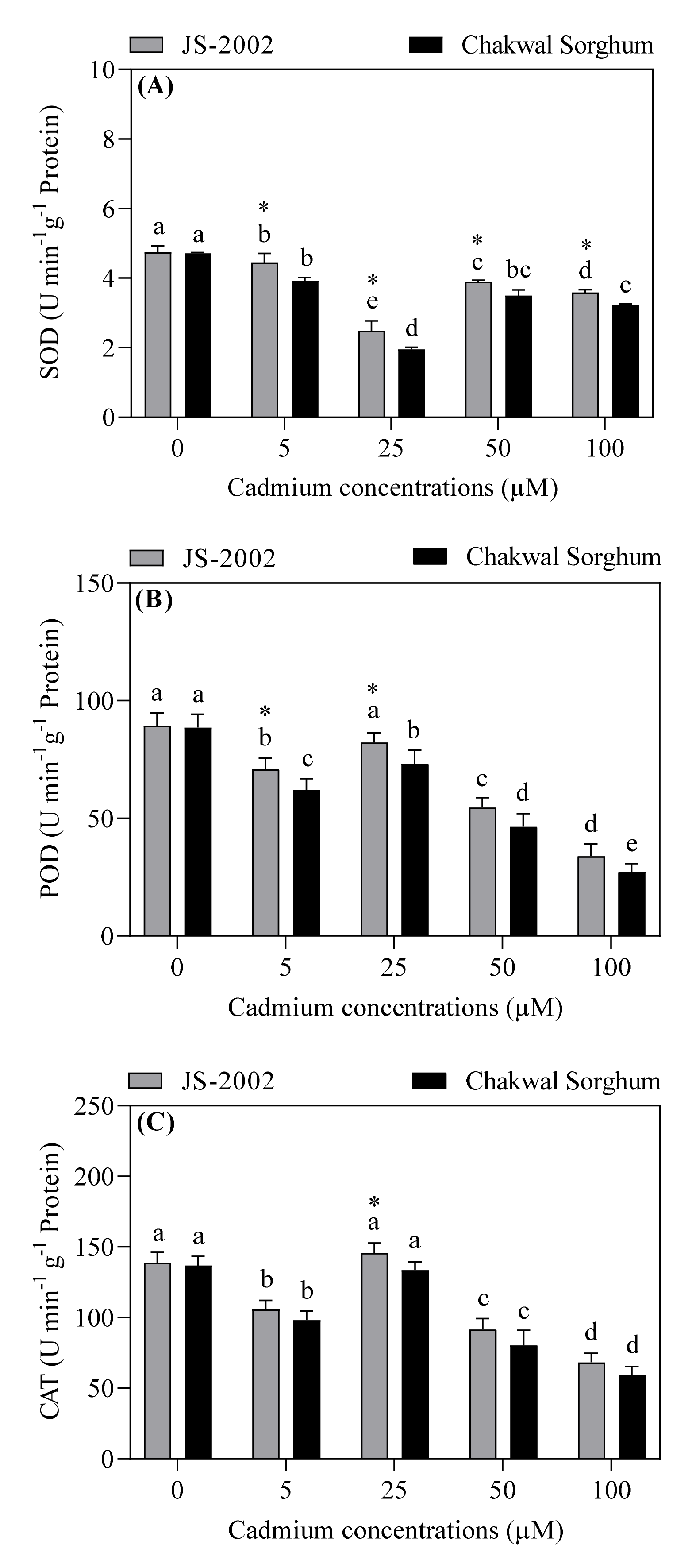
3.4. Effect of Exogenous Cadmium on Antioxidant Enzymes Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawkes, S.J. What is a “heavy metal”? J. Chem. Educ. 1997, 74, 1374. [Google Scholar] [CrossRef]
- De Caroli, M.; Furini, A.; Dalcorso, G.; Rojas, M.; Sansebastiano, G.P.D. Endomembrane Reorganization Induced by Heavy Metals. Plants 2020, 9, 482. [Google Scholar] [CrossRef] [PubMed]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Kim, D.-H.; Lee, S.-Y.; Kim, K.-M.; Waqas, M.; Jung, H.-Y.; Shin, J.-H.; Kim, J.-G.; Lee, I.-J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Gomes-Junior, R.A.; Moldes, C.A.; Delite, F.S.; Pompeu, G.B.; Gratao, P.L.; Mazzafera, P.; Lea, P.J.; Azevedo, R.A. Antioxidant metabolism of coffee cell suspension cultures in response to cadmium. Chemosphere 2006, 65, 1330–1337. [Google Scholar] [CrossRef]
- Liu, J.; Qian, M.; Cai, G.; Yang, J.; Zhu, Q. Uptake and translocation of Cd in different rice cultivars and the relation with Cd accumulation in rice grain. J. Hazard. Mater. 2007, 143, 443–447. [Google Scholar] [CrossRef]
- Vergine, M.; Aprile, A.; Sabella, E.; Genga, A.; Bellis, L.D. Cadmium Concentration in Grains of Durum Wheat (Triticum turgidum L. subsp. durum). J. Agric. Food Chem. 2017, 65, 6240–6246. [Google Scholar] [CrossRef]
- Grant, C.; Buckley, W.; Bailey, L.; Selles, F. Cadmium accumulation in crops. Can. J. Plant Sci. 1998, 78, 1–17. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Sullivan, L.A.; Kochian, L.V. Characterization of cadmium binding, uptake, and translocation in intact seedlings of bread and durum wheat cultivars. Plant Physiol. 1998, 116, 1413–1420. [Google Scholar] [CrossRef]
- Hall, J.Á.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Roohi, M.; Iqbal, Z.; Ashraf, M.; Öztürk, M.; Gücel, S. Cadmium (Cd) and lead (Pb) induced changes in growth, some biochemical attributes, and mineral accumulation in two cultivars of mung bean [Vigna radiata (L.) Wilczek]. Commun. Soil Sci. Plant Anal. 2016, 47, 405–413. [Google Scholar]
- Doganlar, Z.B.P.; Yurekli, F. Interactions between cadmium and phytochelatin accumulation in two different sunflower cultivars. Fresenius Environ. Bull. 2009, 18, 304–310. [Google Scholar]
- Michel-López, C.Y.; Espadas y Gil, F.; Fuentes Ortíz, G.; Santamaría, J.M.; González-Mendoza, D.; Ceceña-Duran, C.; Grimaldo Juarez, O. Bioaccumulation and effect of cadmium in the photosynthetic apparatus of Prosopis juliflora. Chem. Speciat. Bioavailab. 2016, 28, 1–6. [Google Scholar] [CrossRef]
- Okem, A.; Moyo, M.; Stirk, W.; Finnie, J.; Van Staden, J. Investigating the effect of cadmium and aluminium on growth and stress-induced responses in the micropropagated medicinal plant Hypoxis hemerocallidea. Plant Biol. 2016, 18, 805–815. [Google Scholar] [CrossRef]
- Schat, H.; Sharma, S.S.; Vooijs, R. Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol. Plant. 1997, 101, 477–482. [Google Scholar] [CrossRef]
- Vijendra, P.D.; Huchappa, K.M.; Lingappa, R.; Basappa, G.; Jayanna, S.G.; Kumar, V. Physiological and Biochemical Changes in Moth Bean (Vigna aconitifolia L.) under Cadmium Stress. J. Bot. 2016, 2016. [Google Scholar] [CrossRef]
- Gonçalves, J.F.; Antes, F.G.; Maldaner, J.; Pereira, L.B.; Tabaldi, L.A.; Rauber, R.; Rossato, L.V.; Bisognin, D.A.; Dressler, V.L.; de Moraes Flores, E.M. Cadmium and mineral nutrient accumulation in potato plantlets grown under cadmium stress in two different experimental culture conditions. Plant Physiol. Biochem. 2009, 47, 814–821. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, D.; Hou, W. Hyperaccumulation of cadmium by roots, bulbs and shoots of garlic (Allium sativum L.). Bioresour. Technol. 2001, 76, 9–13. [Google Scholar] [CrossRef]
- Liu, J.; Cai, G.; Qian, M.; Wang, D.; Xu, J.; Yang, J.; Zhu, Q. Effect of Cd on the growth, dry matter accumulation and grain yield of different rice cultivars. J. Sci. Food Agric. 2007, 87, 1088–1095. [Google Scholar] [CrossRef]
- Ozdener, Y.; Kutbay, H.G. Toxicity of copper, cadmium, nickel, lead and zinc on seed germination and seedling growth in Eruca sativa. Fresenius Environ. Bull. 2009, 18, 26–31. [Google Scholar]
- Gounden, D.; Kisten, K.; Moodley, R.; Shaik, S.; Jonnalagadda, S.B. Impact of spiked concentrations of Cd, Pb, As and Zn in growth medium on elemental uptake of Nasturtium officinale (Watercress). J. Environ. Sci. Health Part B 2016, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kutrowska, A.; Małecka, A.; Piechalak, A.; Masiakowski, W.; Hanć, A.; Barałkiewicz, D.; Andrzejewska, B.; Zbierska, J.; Tomaszewska, B. Effects of binary metal combinations on zinc, copper, cadmium and lead uptake and distribution in Brassica juncea. J. Trace Elem. Med. Biol. 2017, 44, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Haq, N.; Williams, I. Phytoaccumulation of arsenic, cadmium and lead by Brassica juncea parents and their F1 hybrids. J. Environ. Prot. 2016, 7, 613–622. [Google Scholar] [CrossRef]
- Wang, M.; Zou, J.; Duan, X.; Jiang, W.; Liu, D. Cadmium accumulation and its effects on metal uptake in maize (Zea mays L.). Bioresour. Technol. 2007, 98, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Bao, M.; Wang, L.; Khan, I.; Ullah, E.; Tung, S.A.; Samad, R.A.; Shahzad, B. Cadmium toxicity in Maize (Zea mays L.): Consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environ. Sci. Pollut. Res. 2015, 22, 17022–17030. [Google Scholar] [CrossRef]
- Khan, N.; Samiullah; Singh, S.; Nazar, R. Activities of antioxidative enzymes, sulphur assimilation, photosynthetic activity and growth of wheat (Triticum aestivum) cultivars differing in yield potential under cadmium stress. J. Agron. Crop Sci. 2007, 193, 435–444. [Google Scholar] [CrossRef]
- Cho, U.-H.; Seo, N.-H. Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci. 2005, 168, 113–120. [Google Scholar] [CrossRef]
- Khaliq, A.; Aslam, F.; Matloob, A.; Hussain, S.; Geng, M.; Wahid, A.; Ur Rehman, H. Seed priming with selenium: Consequences for emergence, seedling growth, and biochemical attributes of rice. Biol. Trace Elem. Res. 2015, 166, 236–244. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J. Plant Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, A.; Zia-ul-Haq, M.; Ali, F.; Aslam, F.; Matloob, A.; Navab, A.; Hussain, S. Salinity tolerance in wheat cultivars is related to enhanced activities of enzymatic antioxidants and reduced lipid peroxidation. Clean Soil Air Water 2015, 43, 1248–1258. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Tanyolac, D.; Ayhan, B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef] [PubMed]
- De Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef] [PubMed]
- De Morais Cardoso, L.; Montini, T.A.; Pinheiro, S.S.; Pinheiro-Sant’Ana, H.M.; Martino, H.S.D.; Moreira, A.V.B. Effects of processing with dry heat and wet heat on the antioxidant profile of sorghum. Food Chem. 2014, 152, 210–217. [Google Scholar] [CrossRef]
- Epelde, L.; Mijangos, I.; Becerril, J.M.; Garbisu, C. Soil microbial community as bioindicator of the recovery of soil functioning derived from metal phytoextraction with sorghum. Soil Biol. Biochem. 2009, 41, 1788–1794. [Google Scholar] [CrossRef]
- Zhuang, P.; Wensheng, S.; Zhian, L.; Bin, L.; Jintian, L.; Jingsong, S. Removal of metals by sorghum plants from contaminated land. J. Environ. Sci. 2009, 21, 1432–1437. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativaL.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef]
- Mukherjee, S.; Choudhuri, M. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Cakmak, I.; Strbac, D.; Marschner, H. Activities of hydrogen peroxide-scavenging enzymes in germinating wheat seeds. J. Exp. Bot. 1993, 44, 127–132. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueño, M.C.; Luis, A.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Daud, M.; Variath, M.; Ali, S.; Najeeb, U.; Jamil, M.; Hayat, Y.; Dawood, M.; Khan, M.I.; Zaffar, M.; Cheema, S.A. Cadmium-induced ultramorphological and physiological changes in leaves of two transgenic cotton cultivars and their wild relative. J. Hazard. Mater. 2009, 168, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.; Safronova, V.I.; Belimov, A.A.; Dietz, K.-J. Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J. Exp. Bot. 2005, 56, 167–178. [Google Scholar] [CrossRef]
- Marchiol, L.; Leita, L.; Martin, M.; Peressotti, A.; Zerbi, G. Physiological responses of two soybean cultivars to cadmium. J. Environ. Qual. 1996, 25, 562–566. [Google Scholar] [CrossRef]
- Akhter, M.; Macfie, S. Species-specific relationship between transpiration and cadmium translocation in lettuce, barley and radish. J. Plant Stud. 2012, 1, 2. [Google Scholar]
- Wagner, G.J. Accumulation of cadmium in crop plants and its consequences to human health. Adv. Agron. 1993, 51, 173–212. [Google Scholar]
- Gomes, M.P.; Soares, A.M. Cadmium effects on mineral nutrition of the Cd-hyperaccumulator Pfaffia glomerata. Biologia 2013, 68, 223–230. [Google Scholar] [CrossRef]
- Jia, G.-X.; Zhu, Z.-Q.; Chang, F.-Q.; Li, Y.-X. Transformation of tomato with the BADH gene from Atriplex improves salt tolerance. Plant Cell Rep. 2002, 21, 141–146. [Google Scholar]
- Jawad Hassan, M.; Ali Raza, M.; Khan, I.; Ahmad Meraj, T.; Ahmed, M.; Abbas Shah, G.; Ansar, M.; Awan, S.A.; Khan, N.; Iqbal, N. Selenium and Salt Interactions in Black Gram (Vigna mungo L): Ion Uptake, Antioxidant Defense System, and Photochemistry Efficiency. Plants 2020, 9, 467. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, Y.; Zhang, X.-Q.; Ma, X.; Huang, L.-K.; Yan, Y.-H. Exogenous spermidine improves seed germination of white clover under water stress via involvement in starch metabolism, antioxidant defenses and relevant gene expression. Molecules 2014, 19, 18003–18024. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Prasad, P.V.; Seppanen, M. Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system. Plant Physiol. Biochem. 2010, 48, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, G.; Dominy, P.; Wu, H.; Bachir, D.M. Differences in yield components and kernel Cd accumulation in response to Cd toxicity in four barley genotypes. Chemosphere 2007, 70, 83–92. [Google Scholar] [CrossRef]
- Sandalio, L.; Dalurzo, H.; Gomez, M.; Romero-Puertas, M.; Del Rio, L. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Chien, H.-F.; Wang, J.-W.; Lin, C.C.; Kao, C.H. Cadmium toxicity of rice leaves is mediated through lipid peroxidation. Plant Growth Regul. 2001, 33, 205–213. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, H.; Qin, R.; Jiang, W.; Liu, D. Cadmium induction of lipid peroxidation and effects on root tip cells and antioxidant enzyme activities in Vicia faba L. Ecotoxicology 2009, 18, 814–823. [Google Scholar] [CrossRef]
- Liu, D.-L.; Zhang, S.-P.; Zheng, C.; Qiu, W.-W. Soil cadmium regulates antioxidases in sorghum. Agric. Sci. China 2010, 9, 1475–1480. [Google Scholar] [CrossRef]
- Liu, J.-N.; Zhou, Q.-X.; Sun, T.; Ma, L.Q.; Wang, S. Growth responses of three ornamental plants to Cd and Cd–Pb stress and their metal accumulation characteristics. J. Hazard. Mater. 2008, 151, 261–267. [Google Scholar] [CrossRef]
- Zornoza, P.; Sánchez-Pardo, B.; Carpena, R.O. Interaction and accumulation of manganese and cadmium in the manganese accumulator Lupinus albus. J. Plant Physiol. 2010, 167, 1027–1032. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jawad Hassan, M.; Ali Raza, M.; Ur Rehman, S.; Ansar, M.; Gitari, H.; Khan, I.; Wajid, M.; Ahmed, M.; Abbas Shah, G.; Peng, Y.; et al. Effect of Cadmium Toxicity on Growth, Oxidative Damage, Antioxidant Defense System and Cadmium Accumulation in Two Sorghum Cultivars. Plants 2020, 9, 1575. https://doi.org/10.3390/plants9111575
Jawad Hassan M, Ali Raza M, Ur Rehman S, Ansar M, Gitari H, Khan I, Wajid M, Ahmed M, Abbas Shah G, Peng Y, et al. Effect of Cadmium Toxicity on Growth, Oxidative Damage, Antioxidant Defense System and Cadmium Accumulation in Two Sorghum Cultivars. Plants. 2020; 9(11):1575. https://doi.org/10.3390/plants9111575
Chicago/Turabian StyleJawad Hassan, Muhammad, Muhammad Ali Raza, Sana Ur Rehman, Muhammad Ansar, Harun Gitari, Imran Khan, Muhammad Wajid, Mukhtar Ahmed, Ghulam Abbas Shah, Yan Peng, and et al. 2020. "Effect of Cadmium Toxicity on Growth, Oxidative Damage, Antioxidant Defense System and Cadmium Accumulation in Two Sorghum Cultivars" Plants 9, no. 11: 1575. https://doi.org/10.3390/plants9111575
APA StyleJawad Hassan, M., Ali Raza, M., Ur Rehman, S., Ansar, M., Gitari, H., Khan, I., Wajid, M., Ahmed, M., Abbas Shah, G., Peng, Y., & Li, Z. (2020). Effect of Cadmium Toxicity on Growth, Oxidative Damage, Antioxidant Defense System and Cadmium Accumulation in Two Sorghum Cultivars. Plants, 9(11), 1575. https://doi.org/10.3390/plants9111575