An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification




Abstract
1. Introduction
2. Results
2.1. Morphological Characteristics
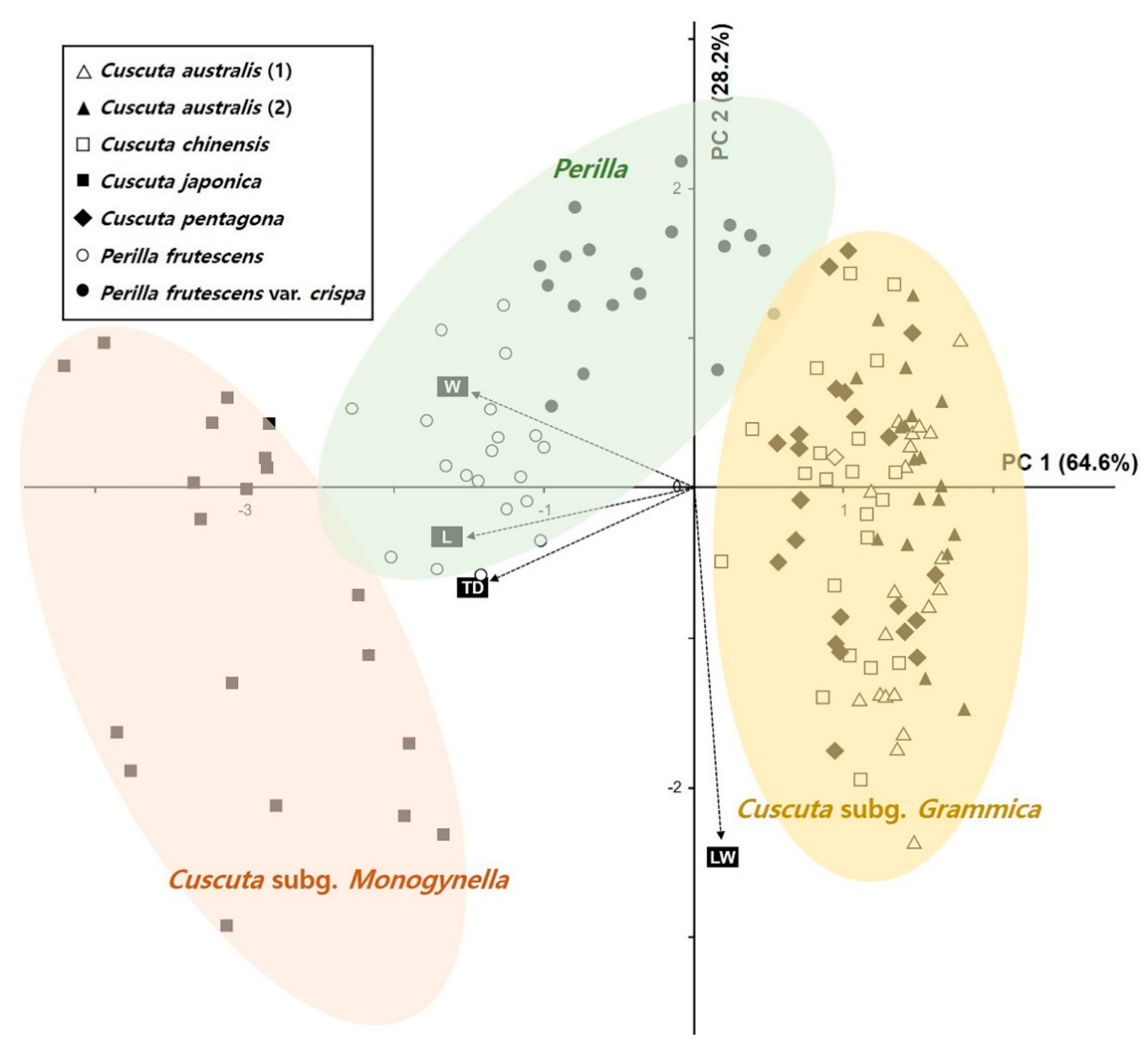
2.2. Statistical Analysis of Morphological Characteristics
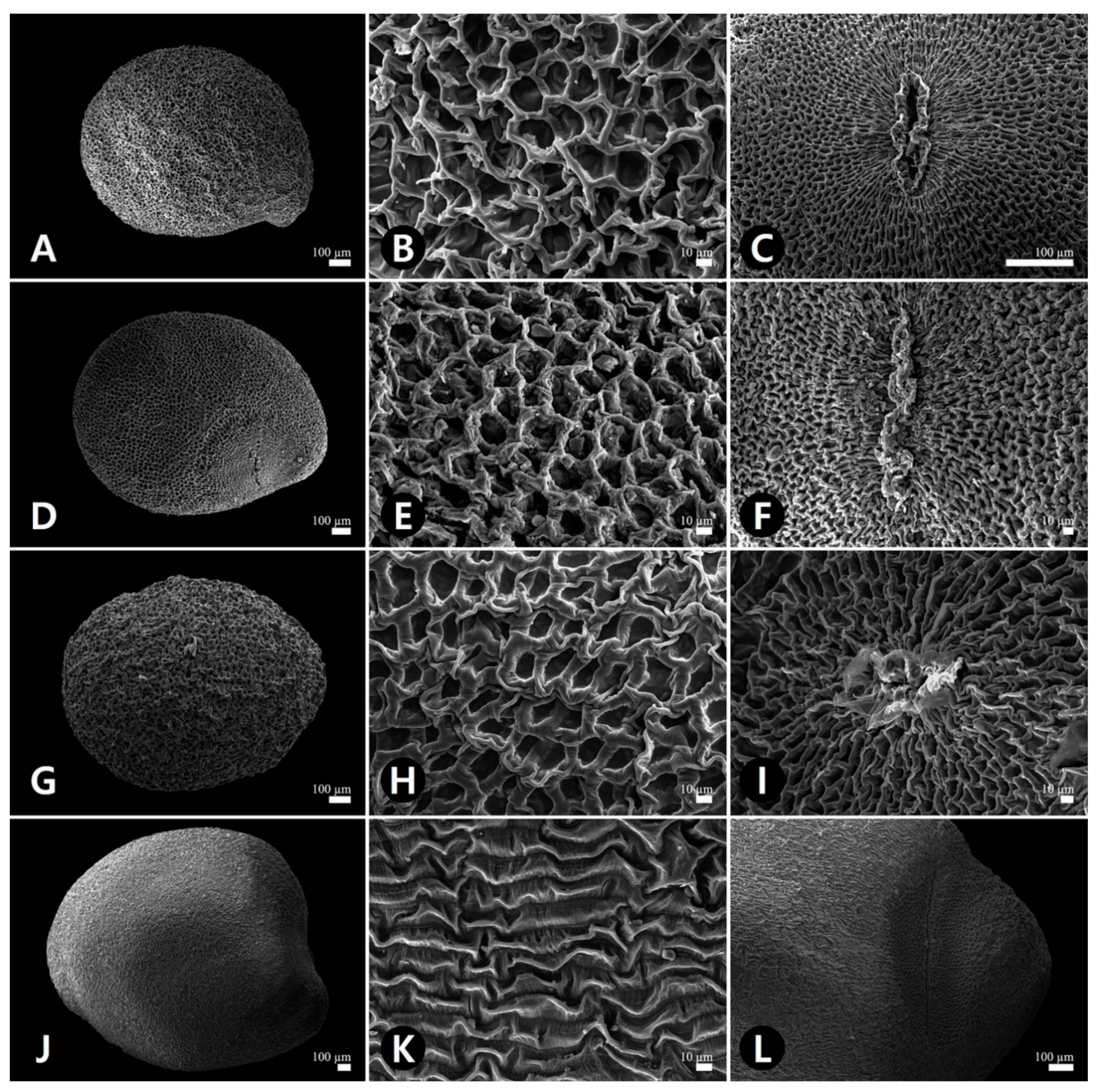
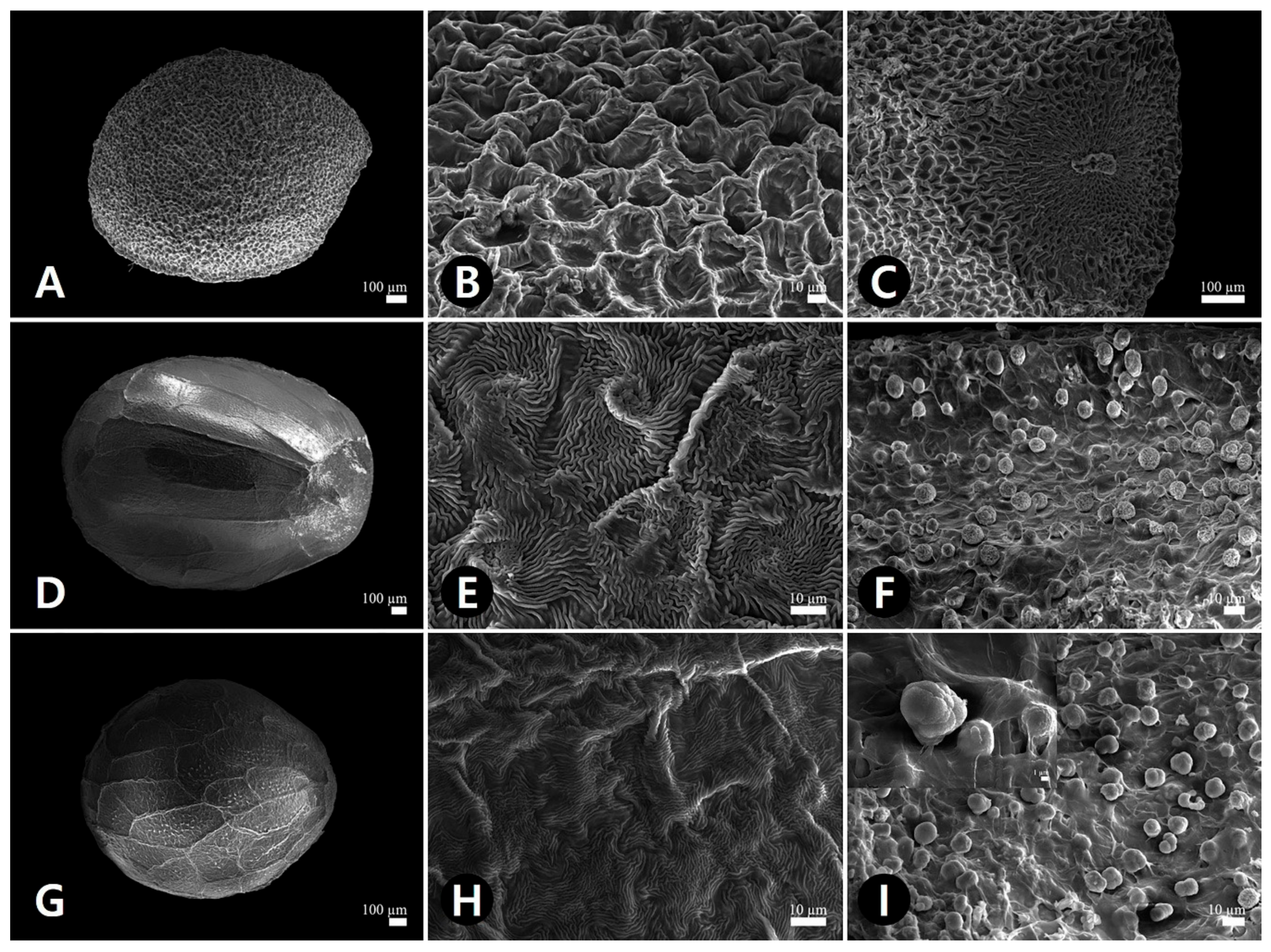
2.3. Micromorphological Characteristics
2.4. Identification Key
- 1.
- Seeds ovoid in shape; triangular cross-section; glabrous on hilum; rugulate and reticulate testa ornamentation ------------------------------------------------------------------------------------------------------------------------------------------------------------------ 2
- 1′.
- Seeds globose to subglobose in shape; circular to broadly triangular in cross-section; glandular hairs on hilum; rugose-colliculate testa ornamentation ----------------------------------------------------------------------------------------------------------------------------------------------------------------- 3
- 2.
- Embryo exudation in boiling water ------------------------------------------------------------------------------------------------------------------- 4
- 2′.
- No embryo exudation in boiling water ---------------------------------------------------- Cuscuta pentagona
- 3.
- Straight anticlinal cell wall boundary ------------------------------------- Perilla frutescens var. frutescens
- 3′.
- Sinuate anticlinal cell wall boundary ------------------------------------------- Perilla frutescens var. crispa
- 4.
- Seeds 2.17–3.26 × 1.57–2.78 mm; elliptic hilum; elongated epidermal cell outline; rugulate testa ornamentation; channeled anticlinal cell wall boundary; convex outer periclinal cell wall ----------------------------------------------------------------------------------------------------- Cuscuta japonica
- 4′.
- Seeds 1.17–1.82 × 1.04–1.46 mm; circular-ovate hilum; isodiametric epidermal cell outline; reticulate testa ornamentation; raised anticlinal cell wall boundary; concave outer periclinal cell wall ------------------------------------------------------------------------------ Cuscuta australis and C. chinensis
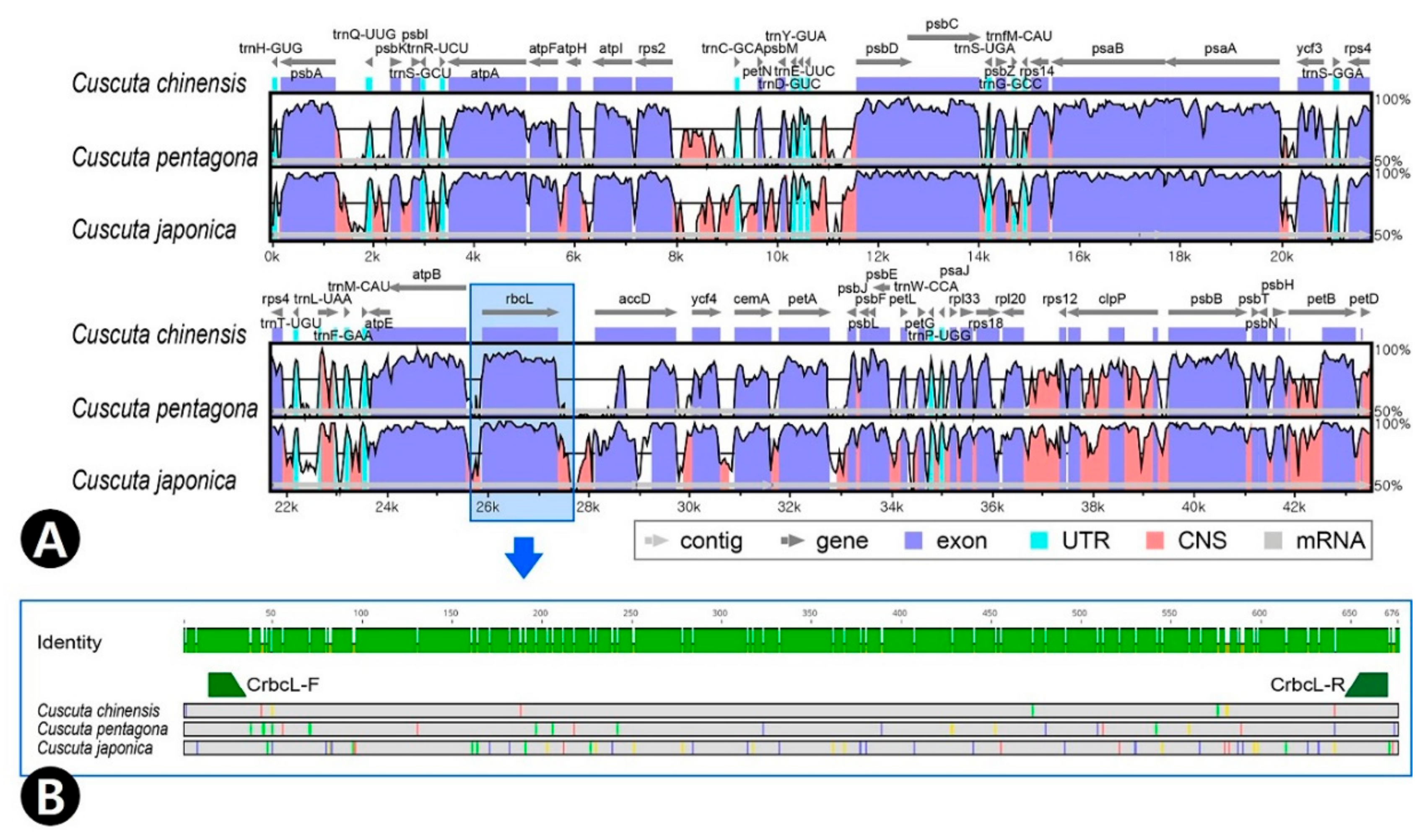
2.5. Comparision of rbcL Using Cuscuta
2.6. Comparision of ITS and rbcL Using Cuscuta and Perilla
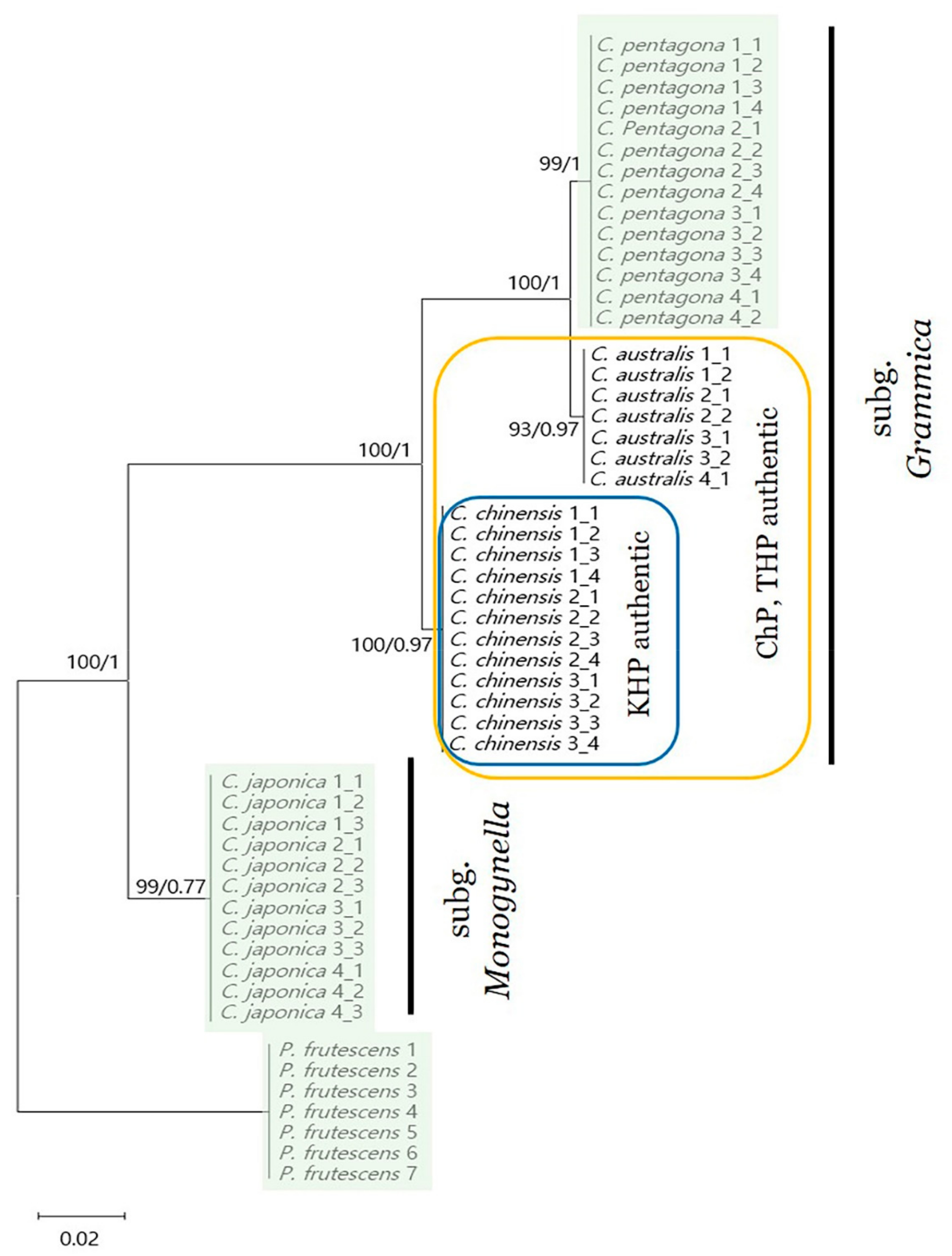
2.7. Phylogenetic Analysis
3. Discussion
3.1. Morphology-Based Identification
3.2. Micromorphology-Based Identification
3.3. DNA Barcoding-Based Identification
4. Materials and Methods
4.1. Plant Materials
4.2. General Morphology and Embryo Exudation
4.3. Statistic Analysis
4.4. Micromorphology
4.5. Sequence Analysis
4.6. Comparative Analysis and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Wyk, B.-E.; Wink, M. Medicinal Plants of the World: An Illustrated Scientific Guide to Important Medicinal Plants and Their Uses, 1st ed.; Briza Pub.: Pretoria, South Africa, 2004; 480p. [Google Scholar]
- Han, J.; Pang, X.; Liao, B.; Yao, H.; Song, J.; Chen, S. An authenticity survey of herbal medicines from markets in China using DNA barcoding. Sci. Rep. 2016, 6, 18723. [Google Scholar] [CrossRef] [PubMed]
- Joharchi, M.R.; Amiri, M.S. Taxonomic evaluation of misidentification of crude herbal drugs marketed in Iran. Avicenna J. Phytomed. 2012, 2, 105–112. [Google Scholar] [PubMed]
- Zhao, Z.; Yuen, J.P.; Wu, J.; Yu, T.; Huang, W. A systematic study on confused species of Chinese materia medica in the Hong Kong market. Ann. Acad. Med. Singap. 2006, 35, 764–769. [Google Scholar] [PubMed]
- Newmaster, S.G.; Grguric, M.; Shanmughanandhan, D.; Ramalingam, S.; Ragupathy, S. DNA barcoding detects contamination and substitution in North American herbal products. BMC Med. 2013, 11, 222. [Google Scholar] [CrossRef]
- Seethapathy, G.S.; Raclariu-Manolica, A.C.; Anmarkrud, J.A.; Wangensteen, H.; De Boer, H.J. DNA metabarcoding authentication of Ayurvedic herbal products on the European market raises concerns of quality and fidelity. Front. Plant Sci. 2019, 10, 68. [Google Scholar] [CrossRef]
- Ichim, M.C. The DNA-based authentication of commercial herbal products reveals their globally widespread adulteration. Front. Pharmacol. 2019, 10, 1227. [Google Scholar] [CrossRef]
- Ichim, M.C.; Häser, A.; Nick, P. Microscopic authentication of commercial herbal products in the globalized market: Potential and limitations. Front. Pharmacol. 2020, 11, 876. [Google Scholar] [CrossRef]
- Jamshidi-Kia, F.; Lorigooini, Z.; Amini-Khoei, H. Medicinal plants: Past history and future perspective. J. Herbmed. Pharmacol. 2018, 7, 7. [Google Scholar] [CrossRef]
- Wu, L.; Jiang, W.; Huang, T.; Li, M.; Li, W.; Zhong, H.; Yu, S. Study on the Quality of Chinese Herbal Pieces and the Development of Enterprises. Adv. Econ. Bus. Manag. Res. 2019, 91, 509–513. [Google Scholar]
- Perharic, L.; Shaw, D.; Léon, C.; De Smet, P.A.; Murray, V.S. Possible association of liver damage with the use of Chinese herbal medicine for skin disease. Vet. Hum. Toxicol. 1995, 37, 562–566. [Google Scholar]
- Nortier, J.L.; Vanherweghem, J.L. For patients taking herbal therapy—Lessons from aristolochic acid nephropathy. Nephrol. Dial. Transplant. 2007, 22, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.; Graeme, L.; Pierre, D.; Elizabeth, W.; Kelvin, C. Pharmacovigilance of herbal medicine. J. Ethnopharmacol. 2012, 140, 513–518. [Google Scholar] [CrossRef]
- World Health Organisation (WHO). WHO Guidelines on Safety Monitoring of Herbal Medicines in Pharmacovigilance Systems; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Korea Institute of Oriental Medicine. Defining Dictionary for Medicinal Herbs (Korean, ‘Hanyak Giwon Sajeon’). 2020. Available online: http://https://oasis.kiom.re.kr/herblib/hminfo/hbmcod/hbmcodList.do (accessed on 21 February 2020).
- Korea Food and Drug Administration. The Korean Herbal Pharmacopoeia, 4th ed.; Korea Food and Drug Administration: Seoul, Korea, 2012. [Google Scholar]
- Pharmacopoeia Committee of the DPRK. Pharmacopoeia of Democratic People’s Republic of Korea, 7th ed.; Medicine and Science Press: Pyeongyang, Korea, 2011. [Google Scholar]
- Committee on Chinese Medicine and Pharmacy. Taiwan Herbal Pharmacopeia, 2nd ed.; English Version; Ministry of Health and Welfare: Taipei, Taiwan, 2013. [Google Scholar]
- Chinese Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; Medical Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Moon, B.C.; Choi, G.; Yuan, Y. Origins of Herbal Medicines and Adulterants in Korea and China; Korea Institute of Oriental Medicine: Daejeon, Korea, 2017. [Google Scholar]
- Qin, D.N.; She, B.R.; She, Y.C.; Wang, J.H. Effects of flavonoids from Semen Cuscutae on the reproductive system in male rats. Asian J. Androl. 2000, 2, 99–102. [Google Scholar] [PubMed]
- Qin, D.; She, B.; She, Y. Effects of flavonoids of Semen Cuscutae on reproductive function of animals. Trad. Chin. Drug Res. Clin. Pharmacol. 2000, 11, 349–351. [Google Scholar]
- Pan, H.J.; Sun, H.X.; Pan, Y.J. Adjuvant effect of ethanol extract of Semen Cuscutae on the immune responses to ovalbumin in mice. J. Ethnopharmacol. 2005, 99, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.K.; Yu, Y.L.; Chen, K.C.; Chang, W.T.; Lee, M.S.; Yang, M.J.; Cheng, H.C.; Liu, C.H.; Chen Dz, C.; Chu, C.L. Kaempferol from Semen Cuscutae attenuates the immune function of dendritic cells. Immunobiology 2011, 216, 1103–1109. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, Y.Q.; Liang, Y.Z.; Zou, C.; Liu, H.; Qiu, F.; Liang, C.L.; Jin, X.W.; Su, Z.R.; Dai, Z. Medicinal herbs Fructus corni and Semen cuscutae suppress allograft rejection via distinct immune mechanisms. Oncotarget 2016, 7, 35680–35691. [Google Scholar] [CrossRef]
- Bao, X.; Wang, Z.; Fang, J.; Li, X. Structural features of an immunostimulating and antioxidant acidic polysaccharide from the seeds of Cuscuta chinensis. Planta Med. 2002, 68, 237–243. [Google Scholar] [CrossRef]
- Liu, J.H.; Ho, S.C.; Lai, T.H.; Liu, T.H.; Chi, P.Y.; Wu, R.Y. Protective effects of Chinese herbs on D-galactose-induced oxidative damage. Methods Find. Exp. Clin. Pharmacol. 2003, 25, 447–452. [Google Scholar] [CrossRef]
- Yen, F.L.; Wu, T.H.; Lin, L.T.; Lin, C.C. Hepatoprotective and antioxidant effects of Cuscuta chinensis against acetaminophen-induced hepatotoxicity in rats. J. Ethnopharmacol. 2007, 111, 123–128. [Google Scholar] [CrossRef]
- Nisa, M.; Akbar, S.; Tariq, M.; Hussain, Z. Effect of Cuscuta chinensis water extract on 7,12-dimethylbenz[a]anthracene-induced skin papillomas and carcinomas in mice. J. Ethnopharmacol. 1986, 18, 21–31. [Google Scholar] [CrossRef]
- Lee, J.B.; Lee, B.R. Study on the anti-cancer, anti-metastasis and effects of immune-response of aqua-acupuncture with Cuscutae Semen infusion solution. J. Acupunct. Res. 2001, 18, 94–104. [Google Scholar]
- Kim, E.Y.; Kim, E.K.; Lee, H.S.; Sohn, Y.; Soh, Y.; Jung, H.S.; Sohn, N.W. Protective effects of Cuscutae semen against dimethylnitrosamine-induced acute liver injury in Sprague-Dawley rats. Biol. Pharm. Bull. 2007, 30, 1427–1431. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yen, F.L.; Wu, T.H.; Lin, L.T.; Cham, T.M.; Lin, C.C. Nanoparticles formulation of Cuscuta chinensis prevents acetaminophen-induced hepatotoxicity in rats. Food Chem. Toxicol. 2008, 46, 1771–1777. [Google Scholar] [CrossRef]
- Peng, W.H.; Chen, Y.W.; Lee, M.S.; Chang, W.T.; Tsai, J.C.; Lin, Y.C.; Lin, M.K. Hepatoprotective Effect of Cuscuta campestris Yunck. Whole Plant on Carbon Tetrachloride Induced Chronic Liver Injury in Mice. Int. J. Mol. Sci. 2016, 17, 2056. [Google Scholar] [CrossRef]
- Moon, J.; Ha, M.J.; Shin, M.J.; Kim, O.Y.; Yoo, E.H.; Song, J.; Chung, J.H. Semen Cuscutae Administration Improves Hepatic Lipid Metabolism and Adiposity in High Fat Diet-Induced Obese Mice. Nutrients 2019, 11, 3035. [Google Scholar] [CrossRef]
- Ye, M.; Lee, S.G.; Chung, E.S.; Lim, S.J.; Kim, W.S.; Yoon, H.; Kim, S.K.; Ahn, K.S.; Jang, Y.P.; Bae, H. Neuroprotective effects of cuscutae semen in a mouse model of Parkinson’s disease. Evid. Based Complement. Altern. Med. 2014, 2014, 150153. [Google Scholar] [CrossRef]
- National Institute of Food and Drug Safety Evaluation. The Dispensatory on the Visual and Organoleptic Examination of Herbal Medicine; National Institute of Food and Drug Safety Evaluation: Cheongwon, Korea, 2012. [Google Scholar]
- Choi, G. List of Fake Korean Medicinal Herbs; Korea Institute of Oriental Medicine: Daejeon, Korea, 2016. [Google Scholar]
- Choi, B.H. Cuscuta L. In The Genera of Vascular Plants of Korea; Flora of Korea Editorial Committee; Academic Publishing Co.: Seoul, Korea, 2007; pp. 793–794. [Google Scholar]
- Park, S.H. New Illustrations and Photographs of Naturalized Plants of Korea; Ilchokak: Seoul, Korea, 2009. [Google Scholar]
- Dawson, J.H.; Musselman, L.J.; Wolswinkel, P.; Dörr, I. Biology and control of Cuscuta. Weed Sci. 1994, 6, 265–317. [Google Scholar]
- Costea, M.; Stefanovic, S. Cuscuta jepsonii (Convolvulaceae): An invasive weed or an extinct endemic? Am. J. Bot. 2009, 96, 1744–1750. [Google Scholar] [CrossRef]
- Costea, M.; Garcia, M.A.; Stefanovic, S. A phylogenetically based infrageneric classification of the parasitic plant genus Cuscuta (Dodders, Convolvulaceae). Syst. Bot. 2015, 40, 269–285. [Google Scholar] [CrossRef]
- Welsh, M.; Stefanovic, S.; Costea, M. Pollen evolution and its taxonomic significance in Cuscuta (dodders, Convolvulaceae). Plant Syst. Evol. 2010, 285, 83–101. [Google Scholar] [CrossRef]
- Wright, M.A.R.; Welsh, M.; Costea, M. Diversity and evolution of the gynoecium in Cuscuta (dodders, Convolvulaceae) in relation to their reproductive biology: Two styles are better than one. Plant Syst. Evol. 2011, 296, 51–76. [Google Scholar] [CrossRef]
- Riviere, S.; Clayson, C.; Dockstader, K.; Wright, M.A.R.; Costea, M. To attract or to repel? Diversity, evolution and role of the “most peculiar organ” in the Cuscuta flower (dodder, Convolvulaceae)-the infrastaminal scales. Plant Syst. Evol. 2013, 299, 529–552. [Google Scholar] [CrossRef]
- Funk, H.T.; Berg, S.; Krupinska, K.; Maier, U.G.; Krause, K. Complete DNA sequences of the plastid genomes of two parasitic flowering plant species, Cuscuta reflexa and Cuscuta gronovii. BMC Plant Biol. 2007, 7, 45. [Google Scholar] [CrossRef]
- McNeal, J.R.; Kuehl, J.V.; Boore, J.L.; de Pamphilis, C.W. Complete plastid genome sequences suggest strong selection for retention of photosynthetic genes in the parasitic plant genus Cuscuta. BMC Plant Biol. 2007, 7, 57. [Google Scholar] [CrossRef]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. The complete chloroplast genome of Cuscuta pentagona Engelm. Mitochondrial DNA B 2018, 3, 523–524. [Google Scholar] [CrossRef]
- Ji, C.Y.; Kim, T.J.; Lee, S.H.; Kim, Y.S.; Lee, G.S.; Kim, J.H.; Choi, G.; Ju, Y.S. Identification of morphological appearance of fine seed herbs using stereoscope (Report I)—Celosiae Semen, Celoisae Cristatae Semen, Cuscutae Semen, Perillae Semen. J. Korean Med. 2013, 34, 1–12. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ju, Y.S. Identification of morphological appearance of minute seed herbs using stereoscope (Report II)—Alli Tuberosi Semen, Alli Fistulosi Semen, Malvae Semen, Abutili Semen, Plantaginis Semen, Schizonepetae Semen. Korean J. Herbol. 2016, 31, 61–66. [Google Scholar] [CrossRef]
- Song, J.H.; Moon, B.C.; Choi, G.; Yang, S. Morphological identification of Lepidii Seu Descurainiae Semen and adulterant seeds using microscopic analysis. Appl. Sci. 2018, 8, 2134. [Google Scholar] [CrossRef]
- Song, J.H.; Yang, S.; Choi, G.; Moon, B.C. An external and micromorphological identification for Pharbitidis Semen and its congeneric species. Korea J. Herbol. 2018, 33, 43–51. [Google Scholar] [CrossRef]
- Song, J.H.; Cha, J.M.; Moon, B.C.; Kim, W.J.; Yang, S.; Choi, G. Mantidis Oötheca (mantis egg case) original species identification via morphological analysis and DNA barcoding. J. Ethnopharmacol. 2020, 252, 112574. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Yang, S.; Choi, G. Taxonomic implications of leaf micromorphology using microscopic analysis: A tool for identification and authentication of Korean Piperales. Plants 2020, 9, 566. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Pang, X.; Song, J.; Shi, L.; Yao, H.; Han, J.; Leon, C. A renaissance in herbal medicine identification: From morphology to DNA. Biotechnol. Adv. 2014, 32, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Tnah, L.H.; Lee, S.L.; Tan, A.L.; Lee, C.T.; Ng, K.K.S.; Ng, C.H.; Farhanah, Z.N. DNA barcode database of common herbal plants in the tropics: A resource for herbal product authentication. Food Control 2019, 95, 318–326. [Google Scholar] [CrossRef]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant Biotechnol. J. 2016, 14, 8–21. [Google Scholar] [CrossRef]
- Kim, C.C.; Chung, Y.J.; Oh, S.M. Taxonomic evaluation of selected Cuscuta species (Cuscutaceae) based on seed morphology. Korean J. Weed Sci. 2000, 20, 255–263. [Google Scholar]
- Lyshede, O.B. Studies on mature seeds of Cuscuta pedicellata and C. campestris by electron microscopy. Ann. Bot. 1992, 69, 365–371. [Google Scholar] [CrossRef]
- Park, I.; Song, J.H.; Yang, S.; Kim, W.J.; Choi, G.; Moon, B.C. Cuscuta species identification based on the morphology of reproductive organs and complete chloroplast genome sequences. Int. J. Mol. Sci. 2019, 20, 2726. [Google Scholar] [CrossRef]
- Abdel Khalik, K.N. Seed morphology of Cuscuta L. (Convolvulaceae) in Egypt and its systematic significance. Feddes Repert. 2006, 117, 217–224. [Google Scholar] [CrossRef]
- Abdel Khalik, K. Systematic implications of seed coat diversity in some representatives of the genus Ipomoea (Convolvulaceae). Turk. J. Bot. 2013, 37, 811–824. [Google Scholar] [CrossRef]
- He, X.H.; Yang, W.Z.; Ye, M.; Wang, Q.; Guo, D. Differentiation of Cuscuta chinensis and Cuscuta australis by HPLC-DAD-MS Analysis and HPLC-UV Quantitation. Planta Med. 2011, 77, 1950–1957. [Google Scholar] [CrossRef] [PubMed]
- Kshirsagar, P.; Umdale, S.; Chavan, J.; Gaikwad, N. Molecular authentication of medicinal plant, Swertia chirayita and its adulterant species. Proc. Natl. Acad. Sci. India Sect. B 2017, 87, 101–107. [Google Scholar] [CrossRef]
- Li, M.; Cao, H.; But, P.P.H.; Shaw, P.C. Identification of herbal medicinal materials using DNA barcodes. J. Syst. Evol. 2011, 49, 271–283. [Google Scholar] [CrossRef]
- Parveen, I.; Gafner, S.; Techen, N.; Murch, S.J.; Khan, I.A. DNA barcoding for the identification of botanicals in herbal medicine and dietary supplements: Strengths and limitations. Planta Med. 2016, 82, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Raclariu, A.C.; Heinrich, M.; Ichim, M.C.; de Boer, H. Benefits and limitations of DNA barcoding and metabarcoding in herbal product authentication. Phytochem. Anal. 2018, 29, 123–128. [Google Scholar] [CrossRef]
- Yuncker, T.G.; Cuscuta, L. Cuscuta L. In North American Flora II; Part 4; The New York Botanical Garden Press: New York, NY, USA, 1965; pp. 1–51. [Google Scholar]
- Fang, R.; Lytton, J.M.; Uzi, P. Cuscuta Linnaeus. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1995; Volume 16, pp. 322–325. [Google Scholar]
- Griesbach, R.J.; Austin, S. Comparison of the Munsell and Royal Horticultural Society’s color charts in describing flower color. Taxon 2005, 54, 771–773. [Google Scholar] [CrossRef]
- Grandin, U. PC-ORD version 5: A user-friendly toolbox for ecologists. J. Veg. Sci. 2006, 17, 843–844. [Google Scholar] [CrossRef]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Barthlott, W. Microstructural features of seed surface. In Current Concepts in Plant Taxonomy; Heywood, V.H., Moore, D.C., Eds.; Academic Press: London, UK, 1984; pp. 95–105. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR Protocols; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Shape a | Cross-Sectional Shape b | Length (mm) | Width (mm) | L/W | Color c | Embryo d |
|---|---|---|---|---|---|---|---|
| Cuscuta australis (1) | ovo | tri | 1.17-(1.48)-1.72 | 1.06-(1.12)-1.24 | 1.08-(1.32)-1.56 | Gr-Br (199A-B) | + |
| Cuscuta australis (2) | ovo | tri | 1.25-(1.39)-1.53 | 0.98-(1.15)-1.34 | 1.06-(1.22)-1.43 | Gr-Br (N199A-B), Bl (202A) | + |
| Cuscutachinensis | ovo | tri | 1.29-(1.56)-1.82 | 1.04-(1.25)-1.46 | 1.03-(1.24)-1.50 | Br (200A-D), Gr-Br (199A-B) | + |
| Cuscutajaponica | ovo | tri | 2.17-(2.71)-3.26 | 1.57-(2.16)-2.78 | 1.05-(1.27)-1.57 | Gr-Br (N199C-D), Bl (203) | + |
| Cuscutapentagona | ovo | tri | 1.36-(1.65)-1.87 | 1.09-(1.32)-1.47 | 1.03-(1.24)-1.48 | Br (200A-D) | − |
| Perilla frutescens var. frutescens | glo to subglo | cir to broadly tri | 2.19-(2.40)-2.83 | 1.79-(2.00)-2.28 | 1.06-(1.20)-1.39 | Gr-Br (199A-B, N199A-C) | − |
| Perilla frutescens var. crispa | glo to subglo | cir to broadly tri | 1.58-(1.97)-2.45 | 1.54-(1.87)-2.11 | 0.96-(1.05)-1.20 | Gr-Br (199A-B, N199A-C) | − |
| Taxa | Ornamentation a (Type) | Cell Outline b | Epidermal Cell Diameter (μm) | Anticlinal Cell Wall c | Periclinal Cell Wall d | Hilum Shape e | Hairy f |
|---|---|---|---|---|---|---|---|
| Cuscuta australis (1) | ret (I) | iso | 11.0-(21.5)-27.4 | str, rsd | ccv | cir-ova | - |
| Cuscuta australis (2) | ret (I) | iso | 15.7-(20.2)-26.3 | str, rsd | ccv | cir-ova | - |
| Cuscutachinensis | ret (I) | iso | 19.5-(25.4)-35.0 | str, rsd | ccv | cir-ova | - |
| Cuscutajaponica | rug (II) | elg | 49.3-(70.8)-99.2 | str, chn | cvx | ell | - |
| Cuscutapentagona | ret (I) | iso | 15.2-(19.5)-24.3 | str, rsd | ccv | cir-ova | - |
| Perilla frutescens var. frutescens | rgs-col (III-1) | elg | 32.6-(40.5)-53.0 | str, chn | cvx | cir | gl |
| Perilla frutescens var. crispa | rgs-col (III-2) | elg | 20.8-(24.8)-39.8 | sin, chn | cvx | cir | gl |
| Primer Name | Primer Sequence (5′ –> 3′) | Position |
|---|---|---|
| CrbcL-F | GGTACATGGACAACTGTGTGG | rbcL |
| CrbcL-R | TGAGCCAAAGAAGTATTTGCAGTG |
| Region | Species | Alignment Length | Parsimony Informative Site | Variable site | Nucleotides Diversity (Pi) | No. of Haplotypes | ||
|---|---|---|---|---|---|---|---|---|
| Number | % | Number | % | |||||
| ITS | Four Cuscuta | 798 | 202 | 25.3% | 205 | 25.7% | 0.08883 | 13 |
| C. pentagona | 784 | 25 | 3.2% | 27 | 3.4% | 0.01661 | 9 | |
| C. australis | 784 | 0 | 0.0% | 0 | 0.0% | 0 | 1 | |
| C. chinensis | 784 | 0 | 0.0% | 0 | 0.0% | 0 | 1 | |
| C. japonica | 793 | 0 | 0.0% | 1 | 0.1% | 0.00042 | 2 | |
| P. frutescens | 615 | 5 | 0.8% | 0 | 0.0% | 0.00461 | 4 | |
| Total | 809 | 287 | 35.5% | 289 | 35.7% | 0.17662 | 17 | |
| rbcL | Four Cuscuta | 656 | 70 | 10.7% | 70 | 10.7% | 0.04645 | 4 |
| C. pentagona | 656 | 0 | 0.00% | 0 | 0.0% | 0 | 1 | |
| C. australis | 656 | 0 | 0.00% | 0 | 0.0% | 0 | 1 | |
| C. chinensis | 656 | 0 | 0.00% | 0 | 0.0% | 0 | 1 | |
| C. japonica | 656 | 0 | 0.00% | 0 | 0.0% | 0 | 1 | |
| P. frutescens | 656 | 0 | 0.00% | 0 | 0.0% | 0 | 1 | |
| Total | 656 | 102 | 15.5% | 102 | 15.50% | 0.06212 | 5 | |
| Scientific Name | Collection Site (Commercial Suppliers) | Collection Date | Voucher No. |
|---|---|---|---|
| Cuscuta australis (1) †,‡ | Hapcheon-gun, Gyeongsangnam-do, Korea | 5 August 2017 | NIBRVP0000652021 M |
| Hapcheon-gun, Gyeongsangnam-do, Korea | 20 August 2019 | KIOM201901022449 M | |
| Hapcheon-gun, Gyeongsangnam-do, Korea | 20 August 2019 | KIOM201901022450 B,M | |
| Cuscuta australis (2) †,‡ | China ∥ (Kwang Myung Dang Co., Ulsan, Korea) | 12 May 2017 | 2-19-0369 M |
| Cuscutachinensis *,† | Hallim-eup, Jeju-si, Jeju, Korea | 29 August 2016 | KIOM201601017927 B,M |
| Hallim-eup, Jeju-si, Jeju, Korea | 29 August 2016 | KIOM201601017928 B,M | |
| Yeongcheon-si, Gyeongsangbuk-do, Korea | 2 August 2016 | NIBRGR0000431983 B,M | |
| Cuscutajaponica‡ | Danyang-gun, Chungcheongbuk-do, Korea | 28 July 2016 | KIOM201701018784 B,M |
| Bonghwa-gun, Gyeongsangbuk-do, Korea | 25 August 2016 | MBC_KIOM-2016-279 B,M | |
| Gujwa-eup, Jeju-si, Jeju, Korea | 29 August 2016 | NIBRVP0000646517 M | |
| Jeongseon-gun, Gangwon-do, Korea | 13 October 2016 | NIBRVP0000591879 M | |
| Cuscutapentagona | Aewol-eup, Jeju-si, Jeju, Korea | 29 August 2016 | KIOM201601017931 B,M |
| Yuseong-gu, Daejeon, Korea | 27 July 2016 | KIOM201701018523 B,M | |
| Jinan-gun, Jeollabuk-do, Korea | 3 October 2016 | MBC_KIOM-2016-365 B | |
| Danyang-gun, Chungcheongbuk-do, Korea | 28 July 2016 | KIOM201701018786 M | |
| Perilla frutescens var. frutescens § | Inje-gun, Gangwon-do, Korea | 8 October 2010 | NIBRVP0000272211 M |
| Perilla frutescens var. crispa § | Buan-gun, Jellabuk-do, Korea | 13 October 2006 | VP-CNNU-356073-5524 M |
| Daejeong-eup, Seogwipo-si, Jeju, Korea | 29 August 2016 | KOSPVP0000238074 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, I.; Yang, S.; Choi, G.; Moon, B.C.; Song, J.-H. An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification. Plants 2020, 9, 1410. https://doi.org/10.3390/plants9111410
Park I, Yang S, Choi G, Moon BC, Song J-H. An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification. Plants. 2020; 9(11):1410. https://doi.org/10.3390/plants9111410
Chicago/Turabian StylePark, Inkyu, Sungyu Yang, Goya Choi, Byeong Cheol Moon, and Jun-Ho Song. 2020. "An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification" Plants 9, no. 11: 1410. https://doi.org/10.3390/plants9111410
APA StylePark, I., Yang, S., Choi, G., Moon, B. C., & Song, J.-H. (2020). An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification. Plants, 9(11), 1410. https://doi.org/10.3390/plants9111410