Agro-Morphological Characterization of Sicilian Chili Pepper Accessions for Ornamental Purposes


 ,
,  ,
, 
 ,
, 
Abstract

1. Introduction
2. Results
2.1. Germplasm Evaluation
2.2. Morphological and Agronomical Characterization of 19 Accessions
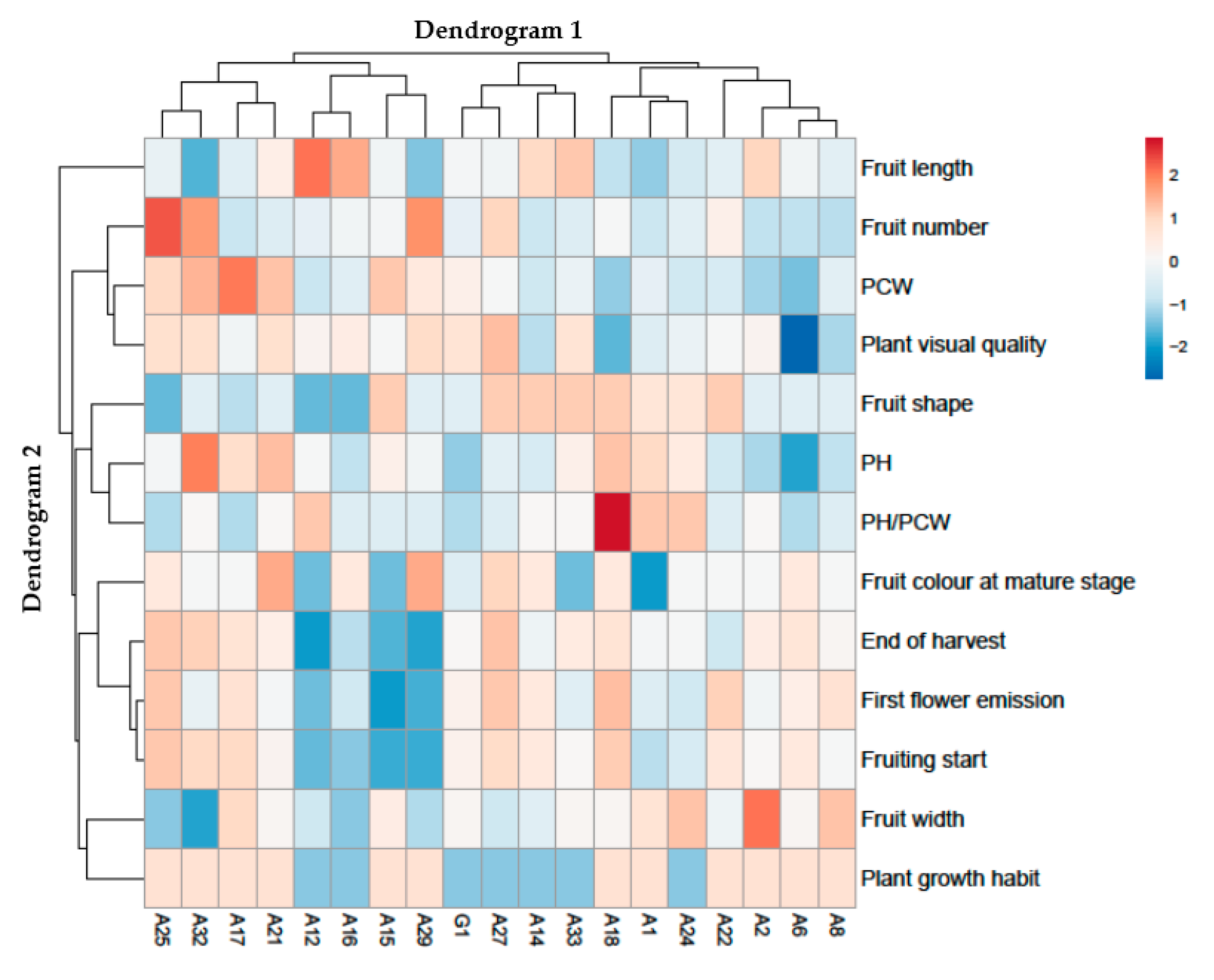
2.3. Heat Map Analysis of All Morphological and Agronomical Descriptors
3. Discussion
4. Materials and Methods
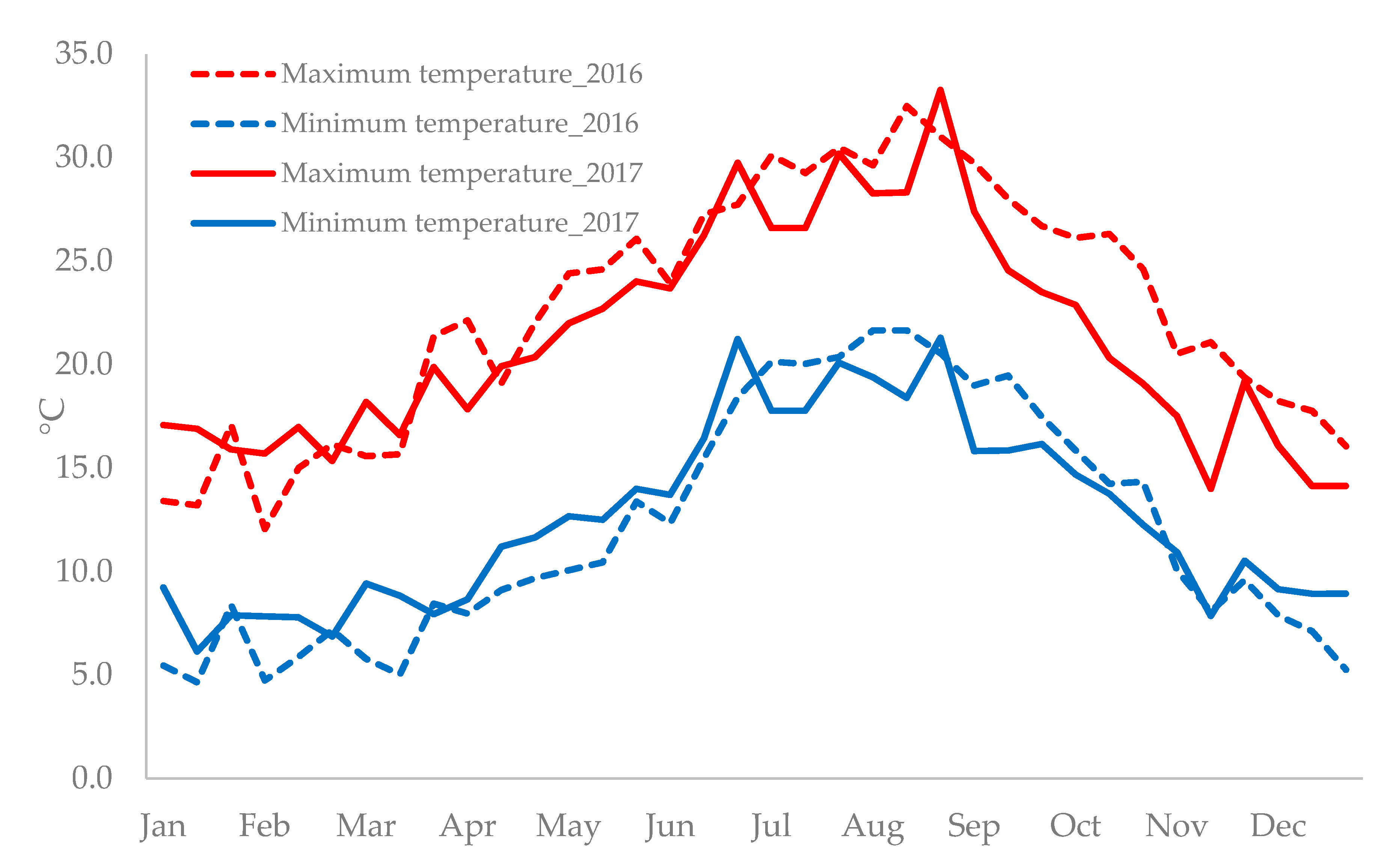
4.1. Germplasm Establishment
4.2. Germplasm Characterization
4.3. Experimental Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Conforti, F.; Statti, G.A.; Menichini, F. Chemical and biological variability of hot pepper fruits (Capsicum annuum var. acuminatum L.) in relation to maturity stage. Food Chem. 2007, 102, 1096–1104. [Google Scholar] [CrossRef]
- Bianchi, G.; Lo Scalzo, R. Characterization of hot pepper spice phytochemicals, taste compounds content and volatile profiles in relation to the drying temperature. J. Food Biochem. 2018, 42, e12675. [Google Scholar] [CrossRef]
- Dorantes, L.; Colmenero, R.; Hernandez, H.; Mota, L.; Jaramillo, M.E.; Fernandez, E.; Solano, C. Inhibition of growth of some food borne pathogenic bacteria by Capsicum annuum extracts. Int. J. Food Microbiol. 2000, 57, 125–128. [Google Scholar] [CrossRef]
- Perucka, I.; Materska, M. Phenylalanine ammonia-lyase and antioxidant activities of lipophilic fraction of fresh pepper fruits Capsicum annuum L. Innov. Food Sci. Emer. 2001, 2, 189–192. [Google Scholar] [CrossRef]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Ariño, A. Characterization of capsaicinoids and antioxidants in hot peppers as influenced by hybrid and harvesting stage. Plant. Food Hum. Nutr. 2013, 68, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Stommel, J.R.; Bosland, P.W. Ornamental pepper. In Flower Breeding and Genetics; Springer: Dordrecht, The Netherlands, 2007; pp. 561–599. [Google Scholar]
- Nascimento, M.F.; Nascimento, N.F.F.; Rêgo, E.R.; Bruckner, C.H.; Finger, F.L.; Rêgo, M.M. Genetic diversity in a structured family of six generations of ornamental chili peppers (Capsicum annuum). In Proceedings of the XXV International EUCARPIA Symposium Section Ornamentals: Crossing Borders, Melle, Belgium, 28 June–2 July June 2015; pp. 395–401. [Google Scholar] [CrossRef]
- Bosland, P.W. An effective plant field cage to increase the production of genetically pure chile (Capsicum spp.) seed. HortScience 1993, 28, 1053. [Google Scholar] [CrossRef]
- Alvares, R.C. Divergência Genética Entre Acessos de Capsicum Chinense Jacq. Coletados no Sudoeste Goiano. Master’s Thesis, Universidade Federal do Goiás, Aparecida de Goiânia, Brazil, 2011; p. 59. [Google Scholar]
- Rêgo, E.R.; Rêgo, M.M.; Cruz, C.D.; Finger, F.L.; Amaral, D.S.S.L. Genetic diversity analysis of peppers: A comparison of discarding variables methods. Crop. Breed. Appl. Biotechnol. 2003, 3, 19–26. [Google Scholar] [CrossRef]
- D’Anna, F.; Sabatino, L. Morphological and agronomical characterization of eggplant genetic resources from the Sicily area. J. Food Agric. Environ. 2013, 11, 401–404. [Google Scholar]
- Sabatino, L.; Palazzolo, E.; D’Anna, F. Grafting suitability of Sicilian eggplant ecotypes onto Solanum torvum: Fruit composition, production and phenology. J. Food Agric. Environ. 2013, 11, 1195–1200. [Google Scholar]
- Sabatino, L.; Iapichino, G.; Maggio, A.; D’anna, E.; Bruno, M.; D’Anna, F. Grafting affects yield and phenolic profile of Solanum melongena L. landraces. J. Integr. Agric. 2016, 15, 1017–1024. [Google Scholar] [CrossRef]
- Raimondo, F.M.; Gianguzzi, L.; Ilardi, V. Inventario delle specie “a rischio” nella flora vascolare della Sicilia. Quad. di Bot. Ambient. Appl. 1992, 3, 65–132. [Google Scholar]
- Tuttolomondo, T.; Dugo, G.; Leto, C.; Cicero, N.; Tropea, A.; Virga, G.; Leone, R.; Licata, M.; La Bella, S. Agronomical and chemical characterisation of Thymbra capitata (L.) Cav. biotypes from Sicily, Italy. Nat. Prod. Res. 2015, 29, 1289–1299. [Google Scholar] [CrossRef]
- Tuttolomondo, T.; Dugo, G.; Ruberto, G.; Leto, C.; Napoli, E.M.; Potortì, A.G.; Fede, M.R.; Virga, G.; Leone, R.; D’Anna, E.; et al. Agronomical evaluation of Sicilian biotypes of Lavandula stoechas L. spp. stoechas and analysis of the essential oils. J. Essent. Oil Res. 2015, 27, 115–124. [Google Scholar] [CrossRef]
- Paik, S.Y.; Soo, R.K.; Chang, I.S.; Chang, P.Y.; Sung, P.H.; Suk, B.H.; Yun, W.J.; Choi, W.J. Purification and Characterization of Complement-activating Acidic polysaccharides from the Fruits of Capsicum annuum. J. Biochem. Mol. Biol. 2003, 36, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Ramanatha, R.V.; Hodgkin, T. Genetic diversity and conservation and utilization of plant genetic resources. Plant. Cell Tissue Organ. 2002, 68, 1–19. [Google Scholar] [CrossRef]
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicums, 2nd ed.; Centre for Agriculture and Biosciences International: Oxfordshire, UK, 2012; p. 248. [Google Scholar]
- IPGRI, AVRDC, CATIE. Descriptors for Capsicum (Capsicum spp.); International Plant Genetic Resources Institute: Rome, Italy, 1995; p. 51. [Google Scholar]
- Engle, L.M. Characterization of germplasm. In Proceedings of the Vegetable Germplasm Conservation and Management, Organized by the Asian Vegetable Research and Development Center-African Regional Program, Arusha, Tanzania, 26 March–1 April 2000; p. 207. [Google Scholar]
- Sharma, V.K.; Semwal, C.S.; Uniyal, S.P. Genetic variability and character association analysis in bell pepper (Capsicum annuum L.). J. Hortic. For. 2010, 2, 58–65. [Google Scholar]
- Orobiyi, A.; Loko, L.Y.; Sanoussi, F.; Agré, A.P.; Korie, N.; Gbaguidi, A.; Dansi, A. Agro-morphological characterization of chili pepper landraces (Capsicum annuum L.) cultivated in Northern Benin. Genet. Resour. Crop. Ev. 2018, 65, 555–569. [Google Scholar] [CrossRef]
- Orobiyi, A.; Loko, Y.L.; Adjatin, A.; Sanoussi, F.; Gbaguidi, A.; Dansi, A.; Sanni, A. Horticultural practices and varietal diversity of chili pepper (Capsicum annuum L.) in Central and Northern Benin. Genet. Resour. Crop. Evol. 2015, 64, 419–436. [Google Scholar] [CrossRef]
- Bozokalfa, M.K.; Kilic, M. Mathematical modeling in the estimation of pepper (Capsicum annuum L.) fruit volume. Chil. J. Agric. Res. 2010, 70, 626–632. [Google Scholar] [CrossRef]
- Hosamani, R.M. Variability, correlation and path analysis in kharif grown chilli (Capsicum annuum L.) genotypes for different characters. Capsicum Eggplant Newsl. 2003, 22, 43–46. [Google Scholar]
- Rodríguez, Y.; Depestre, T.; Gómez, O. Efficiency of selection in pepper lines (Capsicum annuum), from four sub-populations, in characters of productive interest. Cienc. Investig. Agrar. 2008, 35, 29–40. [Google Scholar]
- Manyasa, E.O.; Silim, S.N.; Christiansen, J.L. Variability patterns in Ugandan pigeonpea landraces. J. SAT Agric. Res. 2009, 7, 1–9. [Google Scholar]
- Lahbib, K.; Bnejdi, F.; El Gazzah, M. Selection of pepper parent from a collection of Capsicum annuum landraces based on genetic diversity. J. Plant. Breed. Crop. Sci. 2013, 5, 68–72. [Google Scholar] [CrossRef]
- Yatung, T.; Dubey, R.K.; Singh, V.; Upadhyay, G. Genetic diversity of chilli (Capsicum annuum L.) genotypes of India based on morpho-chemical traits. AJCS 2014, 8, 97–102. [Google Scholar]
- Bonny, S. L’agriculture écologiquement intensive: Nature et défis. Cah. Agric. 2011, 20, 451–462. [Google Scholar]
- Tripodi, P.; Ficcadenti, N.; Rotino, G.L.; Festa, G.; Bertone, A.; Pepe, A.; Caramanico, R.; Migliori, C.A.; Spadafora, D.; Schiavi, M.; et al. Genotypic and environmental effects on the agronomic, health-related compounds and antioxidant properties of chilli peppers for diverse market destinations. J. Sci. Food Agric. 2019, 99, 4550–4560. [Google Scholar] [CrossRef]
- Sonneveld, C.; Voogt, W. Plant Nutrition of Greenhouse Crops; Springer: New York, NY, USA, 2009. [Google Scholar]
- Yadav, R.K.; Sangwan, R.S.; Sabir, F.; Srivastava, A.K.; Sangwan, N.S. Effect of prolonged water stress on specialized secondary metabolites, peltate glandular trichomes, and pathway gene expression in Artemisia annua L. Plant. Physiol. Bioch. 2014, 74, 70–83. [Google Scholar] [CrossRef]
- Krasteva, L.; Todorova, T. The Bulgarian Solanaceae collections. In Compilers. Solanaceae Genetic Resources in Europe. Report of two Meetings-21 September 2001, Nijmegen, The Netherlands/22 May 2003, Skierniewice, Poland; International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 15. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| 2016 | 2017 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variance | PH (cm) | PCW (cm) | PH/PCW | Plant Growth Habit (5–7) | Plant Visual Quality (1–9) | PH (cm) | PCW (cm) | PH/PCW | Plant Growth Habit (5–7) | Plant Visual Quality (1–9) | ||||||||||
| A1 | 34.0 | bc | 34.5 | efgh | 0.99 | b | 7.0 | a | 7.2 | abc | 33.8 | bc | 34.6 | cd | 0.98 | bcd | 7 | a | 5.5 | fgh |
| A2 | 20.8 | g | 28.8 | hi | 0.72 | def | 7.0 | a | 6.9 | abcd | 22.2 | ef | 26.8 | defg | 0.83 | cdef | 7 | a | 6.6 | cdef |
| A6 | 20.2 | g | 29.5 | ghi | 0.69 | defg | 7.0 | a | 4.0 | g | 13.0 | g | 21.8 | h | 0.59 | g | 7 | a | 5.0 | gh |
| A8 | 21.9 | fg | 35.5 | efg | 0.62 | efg | 7.0 | a | 6.4 | cdef | 22.7 | ef | 32.6 | de | 0.70 | fg | 7 | a | 5.1 | gh |
| A12 | 24.8 | efg | 32.3 | fghi | 0.77 | de | 5.0 | b | 6.5 | cdef | 31.4 | cd | 27.8 | def | 1.13 | b | 5 | b | 7.2 | bcde |
| A14 | 21.6 | g | 32.9 | fghi | 0.65 | defg | 5.0 | b | 5.8 | def | 27.3 | de | 29.5 | defg | 0.93 | bcde | 5 | b | 6.0 | efg |
| A15 | 28.4 | cde | 54.0 | ab | 0.53 | gh | 7.0 | a | 5.8 | def | 31.8 | cd | 38.8 | bc | 0.82 | cdefg | 7 | a | 7.6 | abcd |
| A16 | 21.3 | g | 36.8 | def | 0.58 | fg | 5.0 | b | 5.8 | def | 23.8 | ef | 30.8 | def | 0.77 | cdefg | 5 | b | 8.2 | ab |
| A17 | 31.8 | bcd | 56.7 | a | 0.56 | fgh | 7.0 | a | 7.1 | abcd | 34.9 | abc | 49.2 | a | 0.71 | efg | 7 | a | 6.2 | efg |
| A18 | 32.1 | bcd | 27.5 | i | 1.17 | a | 7.0 | a | 6.3 | cdef | 39.3 | a | 26.3 | fgh | 1.49 | a | 7 | a | 4.6 | h |
| G1 | 21.2 | g | 54.0 | ab | 0.39 | h | 5.0 | b | 7.2 | abc | 19.2 | f | 24.6 | gh | 0.78 | cdefg | 5 | b | 7.2 | bcde |
| A21 | 33.7 | bc | 48.7 | bc | 0.69 | defg | 7.0 | a | 7.1 | abcd | 38.3 | ab | 45.3 | a | 0.85 | cdef | 7 | a | 7.5 | abcd |
| A22 | 23.9 | efg | 33.2 | fghi | 0.72 | def | 7.0 | a | 5.8 | f | 23.8 | ef | 31.3 | def | 0.76 | defg | 7 | a | 7.6 | abcd |
| A24 | 34.7 | b | 35.9 | ef | 0.97 | bc | 5.0 | b | 6.4 | cdef | 26.8 | de | 26.8 | defg | 1.00 | bc | 5 | b | 6.6 | cdef |
| A25 | 29.3 | bcde | 50.2 | b | 0.58 | fg | 7.0 | a | 6.2 | def | 26.5 | de | 38.7 | bc | 0.68 | fg | 7 | a | 8.4 | a |
| A27 | 25.0 | efg | 40.7 | de | 0.62 | efg | 5.0 | b | 7.7 | ab | 26.6 | de | 33.7 | cd | 0.79 | cdefg | 5 | b | 7.8 | abc |
| A29 | 27.7 | def | 42.8 | cd | 0.65 | defg | 7.0 | a | 6.3 | cdef | 27.5 | de | 39.1 | bc | 0.71 | efg | 7 | a | 8.4 | a |
| A32 | 41.6 | a | 52.5 | ab | 0.79 | cde | 7.0 | a | 6.8 | bcde | 38.9 | ab | 43.4 | ab | 0.90 | cdef | 7 | a | 7.8 | abc |
| A33 | 29.2 | bcde | 35.9 | ef | 0.81 | bcd | 5.0 | b | 7.8 | a | 30.3 | cd | 34.8 | cd | 0.88 | cdef | 5 | b | 6.5 | def |
| Significance | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ||||||||||
| Source of Variance | First Flower Emission (DAT) | Fruiting Start (DAT) | End of Harvest (DAT) | Fruit Number (# Plant−1) | Fruit Length (cm) | Fruit Width (cm) | Fruit Color at Mature Stage (1–10) | Fruit Shape (1–8) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | 35.7 | fg | 42.0 | gh | 79.0 | de | 132.5 | i | 1.3 | hij | 1.27 | abcde | 3.0 | g | 5.0 | b |
| A2 | 37.7 | de | 48.3 | ef | 82.0 | bcd | 109.8 | ij | 4.1 | abcd | 1.77 | a | 7.0 | d | 3.0 | c |
| A6 | 39.7 | bc | 50.3 | cde | 84.0 | b | 104.0 | ij | 2.7 | efg | 1.13 | abcdef | 8.0 | c | 3.0 | c |
| A8 | 40.7 | b | 48.7 | def | 80.0 | cde | 78.4 | j | 2.4 | fghi | 1.50 | abc | 7.0 | d | 3.0 | c |
| A12 | 31.7 | h | 39.0 | ij | 63.7 | g | 265.6 | g | 5.4 | A | 0.83 | bcdef | 4.0 | f | 1.0 | e |
| A14 | 40.0 | bc | 51.0 | bcd | 78.3 | e | 137.9 | i | 4.0 | bcde | 0.93 | bcdef | 8.0 | c | 6.0 | a |
| A15 | 29.7 | i | 37.7 | j | 65.3 | g | 345.6 | f | 2.7 | defg | 1.20 | abcde | 4.0 | f | 6.0 | a |
| A16 | 35.0 | g | 40.7 | hi | 71.3 | f | 325.4 | f | 4.7 | ab | 0.63 | ef | 8.0 | c | 1.0 | e |
| A17 | 40.7 | b | 53.3 | ab | 84.3 | b | 113.1 | i | 2.3 | fghi | 1.37 | abcd | 7.0 | d | 2.0 | d |
| A18 | 43.0 | a | 53.7 | a | 84.7 | b | 350.3 | f | 1.7 | ghij | 1.10 | abcdef | 8.0 | c | 6.0 | a |
| G1 | 39.0 | cd | 48.7 | def | 79.3 | de | 273.2 | g | 2.8 | defg | 1.07 | abcdef | 6.0 | e | 3.0 | c |
| A21 | 37.7 | de | 48.3 | ef | 82.0 | bcd | 225.4 | h | 3.3 | cdef | 1.07 | abcdef | 10.0 | a | 3.0 | c |
| A22 | 42.3 | a | 50.7 | cde | 73.3 | f | 428.1 | e | 2.4 | fghi | 1.03 | bcdef | 7.0 | d | 6.0 | a |
| A24 | 35.0 | g | 43.7 | g | 79.7 | de | 245.9 | gh | 2.0 | fghij | 1.53 | ab | 7.0 | d | 5.0 | b |
| A25 | 42.3 | a | 54.3 | a | 89.3 | a | 963.6 | a | 2.5 | fgh | 0.60 | ef | 8.0 | c | 1.0 | e |
| A27 | 43.3 | a | 53.3 | ab | 89.3 | a | 621.6 | d | 2.7 | defg | 0.80 | cdef | 9.0 | b | 6.0 | a |
| A29 | 31.3 | h | 37.7 | j | 63.7 | g | 834.0 | b | 1.1 | ij | 0.67 | def | 10.0 | a | 3.0 | c |
| A32 | 36.7 | ef | 52.7 | abc | 88.0 | a | 793.7 | c | 0.8 | J | 0.43 | f | 7.0 | d | 3.0 | c |
| A33 | 36.0 | fg | 47.3 | f | 83.0 | bc | 224.0 | h | 4.3 | abc | 1.10 | abcdef | 4.0 | f | 6.0 | a |
| Significance | *** | *** | *** | *** | *** | *** | *** | *** | ||||||||
| Source of Variance | First Flower Emission (DAT) | Fruiting Start (DAT) | End of Harvest (DAT) | Fruit Number (# Plant−1) | Fruit Length (cm) | Fruit Width (cm) | Fruit Color at Mature Stage (1–10) | Fruit Shape (1–8) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | 33.0 | g | 38.7 | i | 76.7 | fg | 97.7 | h | 1.3 | hij | 1.4 | abcde | 3.0 | g | 5.0 | b |
| A2 | 34.3 | f | 44.3 | fg | 79.7 | cde | 81.6 | hi | 4.1 | abcd | 1.8 | a | 7.0 | d | 3.0 | c |
| A6 | 36.0 | e | 47.0 | cde | 81.7 | bc | 76.7 | hi | 2.7 | defg | 1.1 | abcdef | 8.0 | c | 3.0 | c |
| A8 | 38.0 | c | 43.3 | gh | 77.7 | defg | 64.9 | i | 2.4 | fghi | 1.5 | ab | 7.0 | d | 3.0 | c |
| A12 | 29.0 | i | 36.0 | j | 59.7 | j | 197.5 | f | 5.4 | a | 0.8 | bcdef | 4.0 | f | 1.0 | e |
| A14 | 37.0 | d | 46.3 | def | 75.0 | g | 102.4 | h | 4.0 | bcde | 0.9 | bcdef | 8.0 | c | 6.0 | a |
| A15 | 27.0 | j | 34.7 | j | 63.7 | i | 252.6 | e | 2.7 | defg | 1.2 | abcde | 4.0 | f | 6.0 | a |
| A16 | 32.0 | h | 36.7 | ij | 68.7 | h | 241.7 | e | 4.7 | ab | 0.6 | ef | 8.0 | c | 1.0 | e |
| A17 | 38.0 | c | 49.3 | ab | 81.7 | bc | 84.5 | hi | 2.4 | fghi | 1.4 | abcd | 7.0 | d | 2.0 | d |
| A18 | 40.0 | a | 50.7 | a | 82.0 | bc | 262.8 | e | 1.7 | ghij | 1.1 | abcdef | 8.0 | c | 6.0 | a |
| G1 | 35.7 | e | 46.0 | def | 73.3 | defg | 204.6 | f | 2.8 | defg | 1.1 | abcdef | 6.0 | e | 3.0 | c |
| A21 | 34.7 | f | 45.3 | defg | 79.0 | cdef | 263.4 | g | 3.3 | cdef | 1.1 | abcdef | 10.0 | a | 3.0 | c |
| A22 | 39.0 | b | 47.3 | bcd | 69.7 | h | 317.4 | d | 2.4 | fghi | 1.0 | bcdef | 7.0 | d | 6.0 | a |
| A24 | 32.0 | h | 41.3 | h | 76.3 | efg | 187.8 | fg | 2.0 | fghij | 1.5 | ab | 7.0 | d | 5.0 | b |
| A25 | 40.0 | a | 50.7 | a | 85.0 | ab | 722.7 | a | 2.5 | fgh | 0.6 | ef | 8.0 | c | 1.0 | e |
| A27 | 39.0 | b | 48.7 | abc | 86.3 | a | 474.5 | c | 2.7 | defg | 0.8 | bcdef | 9.0 | b | 6.0 | a |
| A29 | 27.7 | j | 34.7 | j | 61.7 | ij | 618.8 | b | 1.1 | ij | 0.7 | def | 10.0 | a | 3.0 | c |
| A32 | 34.0 | f | 50.0 | a | 84.3 | ab | 601.9 | b | 0.8 | j | 0.4 | f | 7.0 | d | 3.0 | c |
| A33 | 33.0 | g | 45.0 | efg | 80.0 | cd | 161.4 | g | 4.3 | abc | 1.1 | abcdef | 4.0 | f | 6.0 | a |
| Significance | *** | *** | *** | *** | *** | *** | *** | *** | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virga, G.; Licata, M.; Consentino, B.B.; Tuttolomondo, T.; Sabatino, L.; Leto, C.; La Bella, S. Agro-Morphological Characterization of Sicilian Chili Pepper Accessions for Ornamental Purposes. Plants 2020, 9, 1400. https://doi.org/10.3390/plants9101400
Virga G, Licata M, Consentino BB, Tuttolomondo T, Sabatino L, Leto C, La Bella S. Agro-Morphological Characterization of Sicilian Chili Pepper Accessions for Ornamental Purposes. Plants. 2020; 9(10):1400. https://doi.org/10.3390/plants9101400
Chicago/Turabian StyleVirga, Giuseppe, Mario Licata, Beppe Benedetto Consentino, Teresa Tuttolomondo, Leo Sabatino, Claudio Leto, and Salvatore La Bella. 2020. "Agro-Morphological Characterization of Sicilian Chili Pepper Accessions for Ornamental Purposes" Plants 9, no. 10: 1400. https://doi.org/10.3390/plants9101400
APA StyleVirga, G., Licata, M., Consentino, B. B., Tuttolomondo, T., Sabatino, L., Leto, C., & La Bella, S. (2020). Agro-Morphological Characterization of Sicilian Chili Pepper Accessions for Ornamental Purposes. Plants, 9(10), 1400. https://doi.org/10.3390/plants9101400