Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars


Abstract
1. Introduction
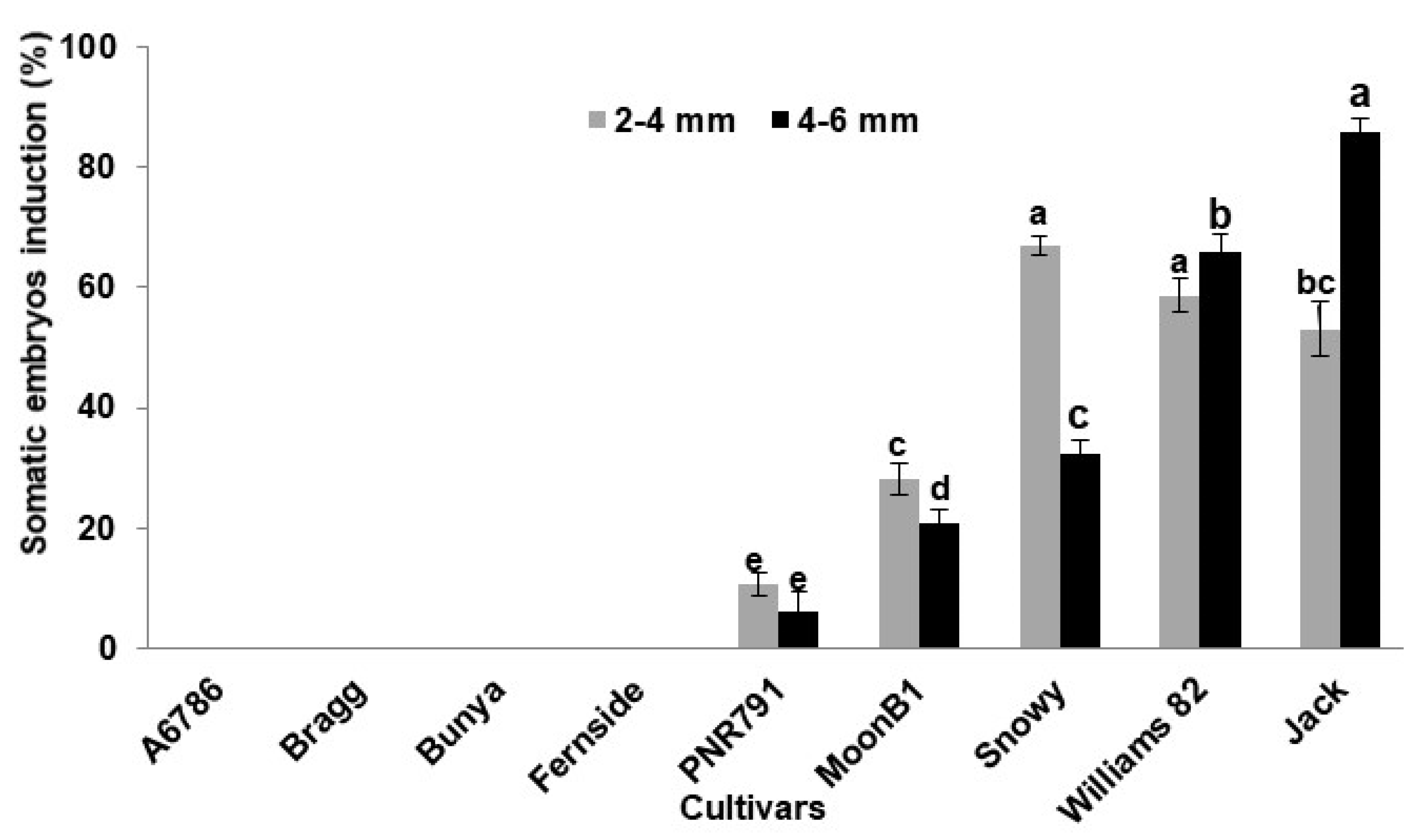
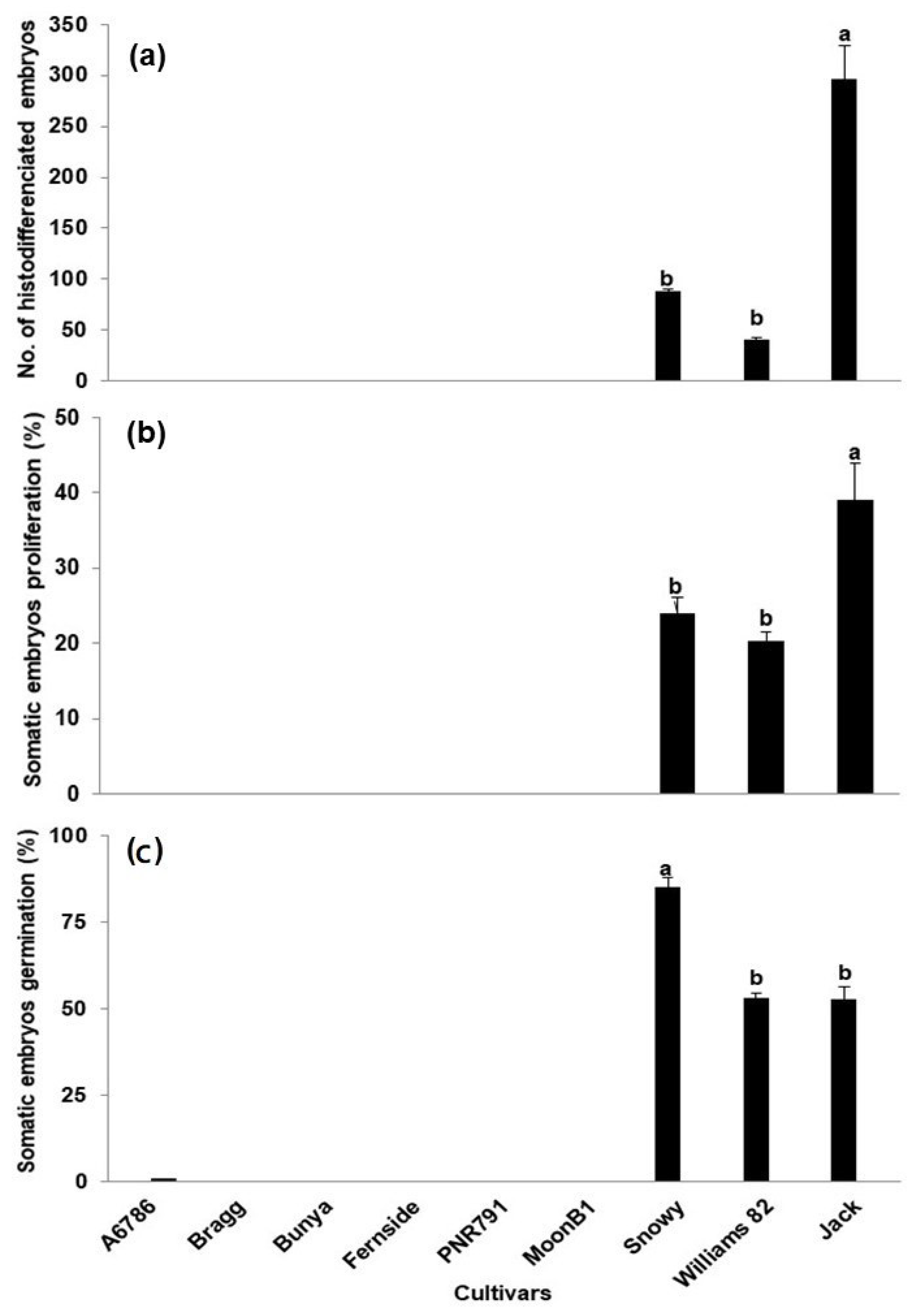
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Statistical Yearbook; Food Agriculture Organization UN: Rome, Italy, 2016. [Google Scholar]
- Birt, D.; Hendrich, S.; Anthony, M.; Alekel, D. Soybeans and the prevention of chronic human disease. In Soybeans: Improvement, Production and Uses; Boerma, H.R., Specht, J.E., Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 1047–1117. [Google Scholar]
- Wilson, R. Seed composition. In Soybean: Improvement, Production and Uses; Boerma, H.R., Specht, J.E., Eds.; ASA; CSSA; SSA: Madison, WI, USA, 2004; pp. 621–677. [Google Scholar]
- Raza, G.; Ahmad, N.; Hussain, M.; Zafar, Y.; Rahman, M. Role of Genetics and Genomics in Mitigating Abiotic Stresses in Soybeans. In Environmental Stresses in Soybean Production; Elsevier: Amsterdam, The Netherlands, 2016; pp. 205–228. [Google Scholar]
- Finer, J.J. Generation of transgenic soybean (Glycine max) via particle bombardment of embryogenic cultures. Curr. Protoc. Plant Biol. 2016, 1, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Raza, G.; Singh, M.B.; Bhalla, P.L. In vitro plant regeneration from commercial cultivars of soybean. Biomed. Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Droste, A.; Pasquali, G.; Bodanese-Zanettini, M. Transgenic fertile plants of soybean [Glycine max (L.) Merrill] obtained from bombarded embryogenic tissue. Euphytica 2002, 127, 367–376. [Google Scholar] [CrossRef]
- Homrich, M.S.; Passaglia, L.M.P.; Pereira, J.F.; Bertagnolli, P.F.; Pasquali, G.; Zaidi, M.A.; Altosaar, I.; Bodanese-Zanettini, M.H. Resistance to Anticarsia gemmatalis Hübner (Lepidoptera, Noctuidae) in transgenic soybean (Glycine max (L.) Merrill Fabales, Fabaceae) cultivar IAS5 expressing a modified Cry1Ac endotoxin. Genet. Mol. Biol. 2008, 31, 522–531. [Google Scholar] [CrossRef]
- Santarem, E.; Pelissier, B.; Finer, J. Effect of explant orientation, pH, solidifying agent and wounding on initiation of soybean somatic embryos. In Vitro Cell. Dev. Biol. Plant 1997, 33, 13–19. [Google Scholar] [CrossRef]
- Meurer, C.A.; Dinkins, R.D.; Redmond, C.T.; McAllister, K.P.; Tucker, D.T.; Walker, D.R.; Parrot, W.A.; Trick, H.N.; Essig, J.S.; Frantz, H.M.; et al. Embryogenic Response of multiple soybean [Glycine max(L.) Merr.] cultivars across three locations. In Vitro Cell. Dev. Biol. Plant 2001, 37, 6. [Google Scholar] [CrossRef]
- Walker, D.; Parrott, W. Effect of polyethylene glycol and sugar alcohols on soybean somatic embryo germination and conversion. Plant Cell Tissue Organ Cult. 2001, 64, 55–62. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, T.; Yu, D.; Gai, J. Somatic embryogenesis and plant regeneration in Chinese soybean (Glycine max (L.) Merr.)—Impacts of mannitol, abscisic acid, and explant age. In Vitro Cell. Dev. Biol. Plant 2009, 45, 180–188. [Google Scholar] [CrossRef]
- Droste, A.; Silva, A.M.D.; Souza, I.F.D.; Strohm, B.W.; Neto, L.B.; Bencke, M.; Sauner, M.V.; Zanettini, M.H.B. Screening of Brazilian soybean genotypes with high potential for somatic embryogenesis and plant regeneration. Pesqui. Agropecu. Bras. 2010, 45, 6. [Google Scholar] [CrossRef][Green Version]
- Texeira, L.R.; Braccini, A.D.L.E.; Churata, B.G.M.; Vieira, E.S.N.; Martins, P.K.; Schuster, I. Evaluation of soybean cultivars on the embryogenic and organogenic potential. Acta Sci. Agron. 2011, 33, 67–74. [Google Scholar] [CrossRef]
- Thankaraj Salammal, M.; Vasudevan Ramesh, A.; Shu-Ye, J.; Andy, G.; Srinivasan, R. In vitro Regeneration and Genetic Transformation of Soybean: Current Status and Future Prospects. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships; IntechOpen: London, UK, 2013; pp. 413–446. [Google Scholar] [CrossRef]
- Samoylov, V.M.; Tucker, D.M.; Parrott, W.A. Soybean [Glycine max (L.) merrill] embryogenic cultures: The role of sucrose and total nitrogen content on proliferation. In Vitro Cell. Dev. Biol. Plant 1998, 34, 8–13. [Google Scholar] [CrossRef]
- Samoylov, V.M.; Tucker, D.M.; Thibaud-Nissen, F.; Parrott, W.A. A liquid-medium-based protocol for rapid regeneration from embryogenic soybean cultures. Plant Cell Rep. 1998, 18, 49–54. [Google Scholar] [CrossRef]
- Schmidt, M.A.; Tucker, D.M.; Cahoon, E.B.; Parrott, W.A. Towards normalization of soybean somatic embryo maturation. Plant Cell Rep. 2005, 24, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, D.H.; Donaldson, P.A. Genotype screening for proliferative embryogenesis and biolistic transformation of short-season soybean genotypes. Plant Cell Rep. 2000, 19, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Tomlin, E.; Branch, S.; Chamberlain, D.; Gabe, H.; Wright, M.; Stewart, C.N., Jr. Screening of soybean, Glycine max L.) Merrill, lines for somatic embryo induction and maturation capability from immature cotyledons. In Vitro Cell. Dev. Biol. Plant 2002, 38, 543–548. [Google Scholar] [CrossRef]
- Ko, T.-S.; Nelson, R.L.; Korban, S.S. Screening Multiple Soybean Cultivars (MG 00 to MG VIII) for Somatic Embryogenesis Following Agrobacterium-Mediated Transformation of Immature Cotyledons. Crop Sci. 2004, 44, 1825–1831. [Google Scholar] [CrossRef]
- Hiraga, S.; Minakawa, H.; Takahashi, K.; Takahashi, R.; Hajika, M.; Harada, K.; Ohtsubo, N. Evaluation of somatic embryogenesis from immature cotyledons of Japanese soybean cultivars. Plant Biotechnol. 2007, 24, 6. [Google Scholar] [CrossRef]
- Parrott, W.A.; Williams, E.G.; Hildebrand, D.F.; Collins, G.B. Effect of genotype on somatic embryogenesis from immature cotyledons of soybean. Plant Cell Tissue Organ Cult. 1989, 16, 15–21. [Google Scholar] [CrossRef]
- Delzer, B.W.; Somers, D.A.; Orf, J.H. Agrobacterium tumefaciens Susceptibility and Plant Regeneration of 10 Soybean Genotypes in Maturity Groups 00 to II. Crop Sci. 1990, 30, 320–322. [Google Scholar] [CrossRef]
- Bailey, M.A.; Boerma, H.R.; Parrott, W.A. Genotype-specific optimization of plant regeneration from somatic embryos of soybean. Plant Sci. 1993, 93, 117–120. [Google Scholar] [CrossRef]
- CSIRO. Snowy—The Best Australian Soybean CSIRO; CSIRO: Canberra, Australia, 2008.
- DPI. Soybean-Growing Guide for Queensland—Variety Update the State of Queenzland; DPI: Melbourne, Australia, 2008. [Google Scholar]
- Gaynor, L.; Lawn, R.; James, A. Agronomic studies on irrigated soybean in southern New South Wales. II. Broadening options for sowing date. Crop Pasture Sci. 2012, 62, 1067–1077. [Google Scholar] [CrossRef]
- Watanabe, S.; Harada, K.; Abe, J. Genetic and molecular bases of photoperiod responses of flowering in soybean. Breed. Sci. 2012, 61, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.; Amberger, L.; Palmer, R.; Oglesby, L.; Ranch, J. Effect of 2,4-dichlorophenoxyacetic acid concentration on somatic embryogenesis and heritable variation in soybean [Glycine max (L) Merr.]. In Vitro Cell. Dev. Biol. 1991, 27, 84–88. [Google Scholar] [CrossRef]
- Tian, L.N.; Brown, D.C.W.; Voldeng, H.; Webb, J. In vitro response and pedigree analysis for somatic embryogenesis of long-day photoperiod adapted soybean. Plant Cell Tissue Organ Cult. 1994, 36, 269–273. [Google Scholar] [CrossRef]
- Komatsuda, T.; Ohyama, K. Genotypes of high competence for somatic embryogenesis and plant regeneration in soybean Glycine max. Theor. Appl. Genet. 1988, 75, 695–700. [Google Scholar] [CrossRef]
- Santos, K.G.B.; Mundstock, E.; Bodanese-Zanettini, M.H. Genotype-specific normalization of soybean somatic embryogenesis through the use of an ethylene inhibitor. Plant Cell Rep. 1997, 16, 859–864. [Google Scholar] [CrossRef]
- Hofmann, N.; Nelson, R.; Korban, S. nfluence of Media Components and pH on Somatic Embryo Induction in Three Genotypes of Soybean. Plant Cell Tissue Organ Cult. 2004, 77, 157–163. [Google Scholar] [CrossRef]
- Ko, T.-S.; Korban, S. Enhancing the frequency of somatic embryogenesis following Agrobacterium-mediated transformation of immature cotyledons of soybean [Glycine max (L.) Merrill.]. In Vitro Cell. Dev. Biol. Plant 2004, 40, 552–558. [Google Scholar] [CrossRef]
- Bailey, M.A.; Boerma, H.R.; Parrott, W.A. Genotype effects on proliferative embryogenesis and plant regeneration of soybean. In Vitro Cell. Dev. Biol. Plant 1993, 29, 102–108. [Google Scholar] [CrossRef]
- Lazzeri, P.; Hildebrand, D.; Collins, G. A procedure for plant regeneration from immature cotyledon tissue of soybean. Plant Mol. Biol. Rep. 1985, 3, 160–168. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Payne, R.W. GenStat. Wiley Interdisciplinary Reviews: Computational Statistics. WIREs 2009, 1, 255–258. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soybean Cultivars | Maturity Group | Total no. of Explants (Both Sizes) | Somatic Embryos Induction (%) (Mean ± S.E) | Proliferated Embryos (Mean ± S.E) | Embryos on Germination Medium | Gemination (%) (Mean ± S.E) | Plants Obtained |
|---|---|---|---|---|---|---|---|
| Snowy | III | 145 | 36 ± 2.59 c | 88 ± 2.31 b | 50 | 85 ± 2.64 a | 42 |
| Jack | II | 141 | 69 ± 2.38 a | 297 ± 32.05 a | 50 | 53 ± 3.54 b | 27 |
| Williams | III | 131 | 62 ± 2.91 b | 41 ± 2.08 c | 50 | 53 ± 1.61 b | 26 |
| MoonB1 | V | 130 | 24 ± 2.38 d | 0 | 0 | 0 | 0 |
| PNR791 | V | 120 | 8 ± 2.60 e | 0 | 0 | 0 | 0 |
| A6785 | VI | 154 | 0 | 0 | 0 | 0 | 0 |
| Bunya | VI | 168 | 0 | 0 | 0 | 0 | 0 |
| Bragg | VII | 70 | 0 | 0 | 0 | 0 | 0 |
| Fernside | VII | 134 | 0 | 0 | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, G.; Singh, M.B.; Bhalla, P.L. Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars. Plants 2020, 9, 38. https://doi.org/10.3390/plants9010038
Raza G, Singh MB, Bhalla PL. Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars. Plants. 2020; 9(1):38. https://doi.org/10.3390/plants9010038
Chicago/Turabian StyleRaza, Ghulam, Mohan B. Singh, and Prem L. Bhalla. 2020. "Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars" Plants 9, no. 1: 38. https://doi.org/10.3390/plants9010038
APA StyleRaza, G., Singh, M. B., & Bhalla, P. L. (2020). Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars. Plants, 9(1), 38. https://doi.org/10.3390/plants9010038