Isolation of Intact Chloroplast for Sequencing Plastid Genomes of Five Festuca Species

 , and
, and
Abstract
1. Introduction
2. Results and Discussion
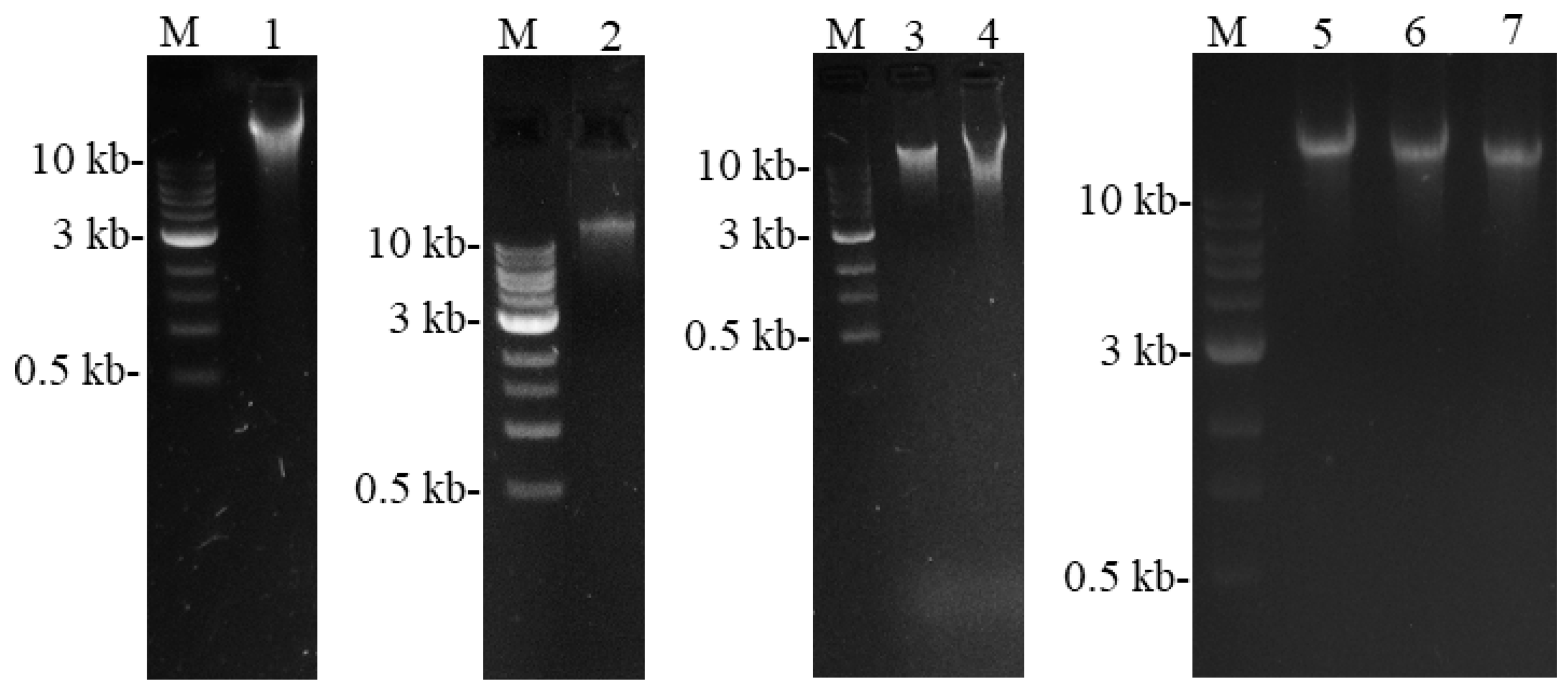
2.1. Intact Chloroplast Isolation
2.2. Extraction of cpDNA
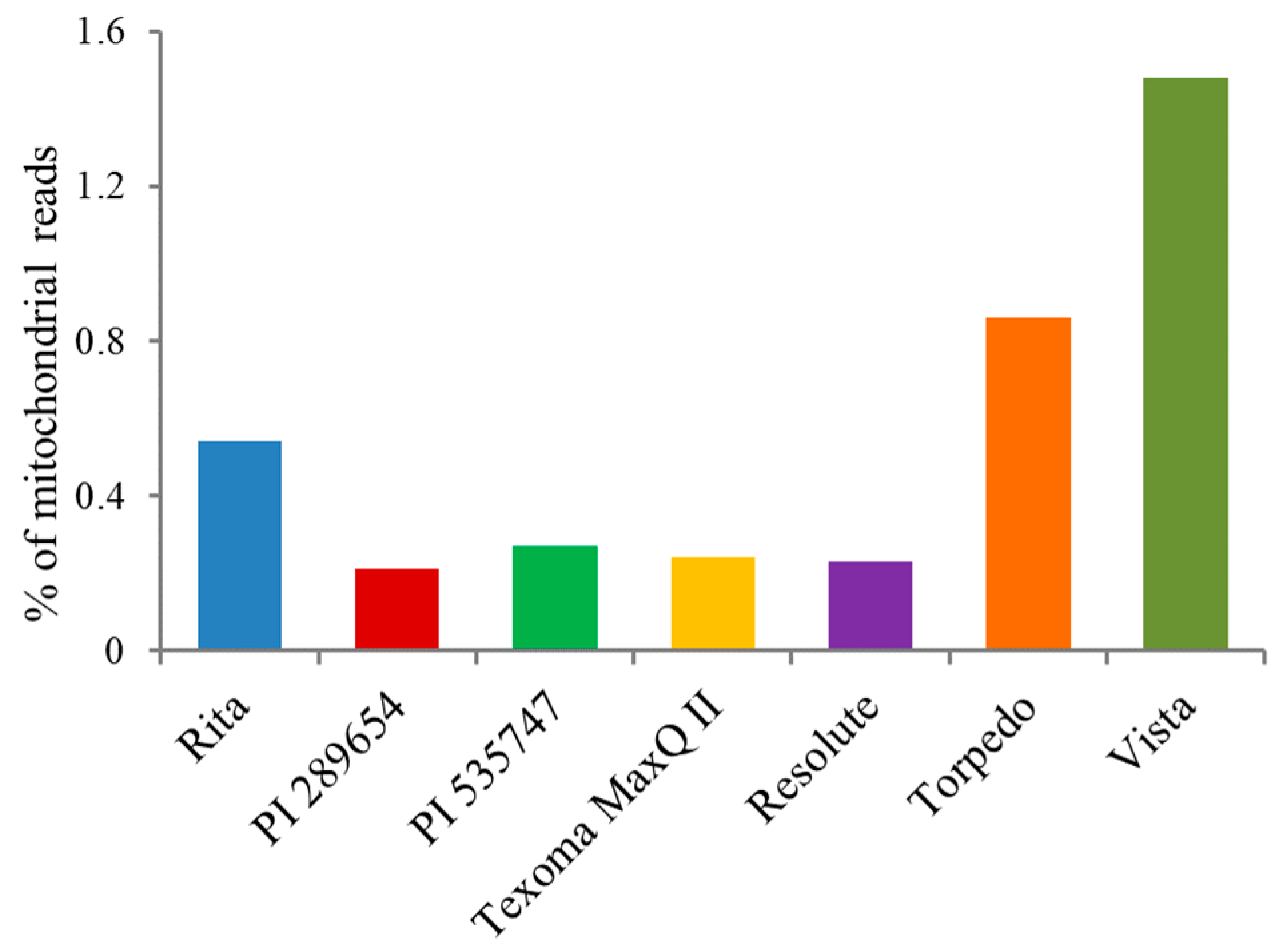
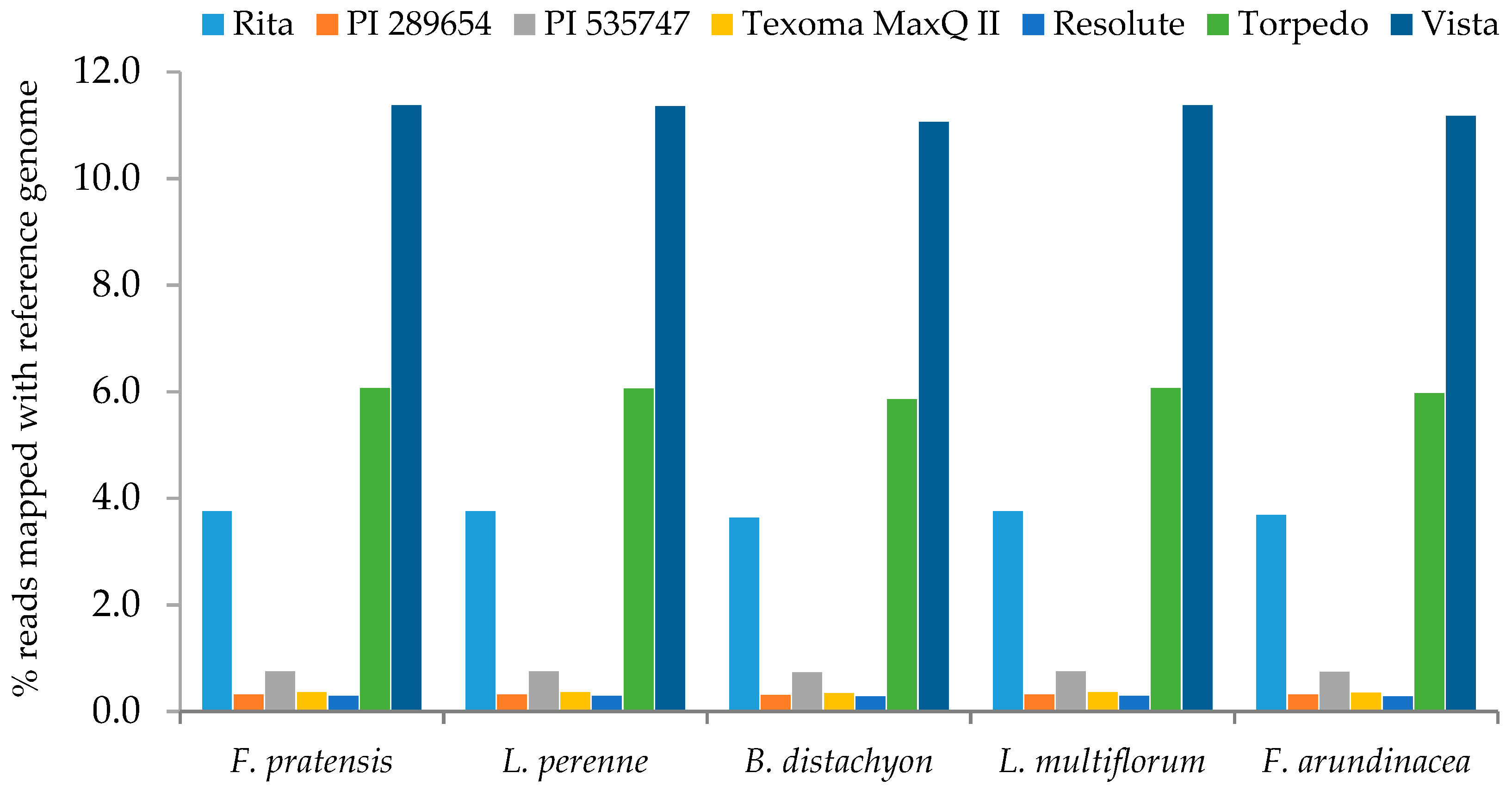
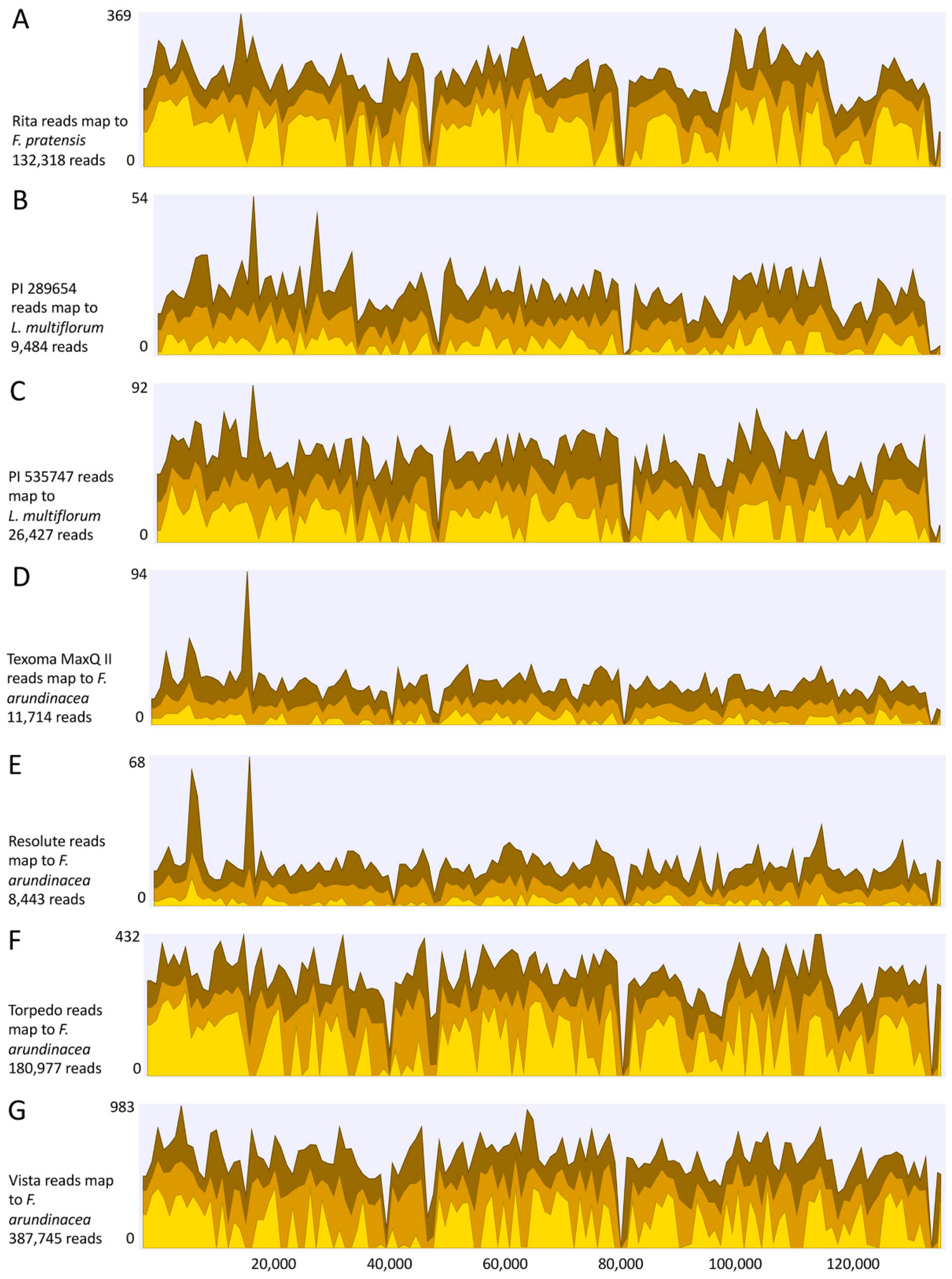
2.3. Sequencing of Plastid Genomes
3. Materials and Methods
3.1. Plant Materials
3.2. Isolation of Intact Chloroplasts
3.3. Purification of Intact Chloroplasts Using Percoll
3.4. Extraction of cpDNA
3.5. Next Generation Sequencing and Estimation of Chloroplast Reads
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.M.; Schmitz-Linneweber, C. Plastid Genomes. In Molecular Biology and Biotechnology of Plant Organelles: Chloroplasts and Mitochondria; Daniell, H., Chase, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 115–150. [Google Scholar] [CrossRef]
- Palmer, J.D.; Jorgensen, R.A.; Thompson, W.F. Chloroplast DNA variation and evolution in pisum: Patterns of change and phylogenetic analysis. Genetics 1985, 109, 195–213. [Google Scholar] [PubMed]
- Greiner, S.; Rauwolf, U.W.E.; Meurer, J.; Herrmann, R.G. The role of plastids in plant speciation. Mol. Ecol. 2011, 20, 671–691. [Google Scholar] [CrossRef] [PubMed]
- Sears, B.B. Elimination of plastids during spermatogenesis and fertilization in the plant kingdom. Plasmid 1980, 4, 233–255. [Google Scholar] [CrossRef]
- Diekmann, K.; Hodkinson, T.R.; Fricke, E.; Barth, S. An optimized chloroplast DNA extraction protocol for grasses (Poaceae) proves suitable for whole plastid genome sequencing and SNP detection. PLoS ONE 2008, 3, e2813. [Google Scholar] [CrossRef] [PubMed]
- Price, C.A.; Cushman, J.C.; Mendiola-Morgenthaler, L.R.; Reardon, E.M. Isolation of plastids in density gradients of percoll and other silica sols. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 157–179. [Google Scholar]
- Vieira, L.d.N.; Faoro, H.; Fraga, H.P.d.F.; Rogalski, M.; de Souza, E.M.; de Oliveira Pedrosa, F.; Nodari, R.O.; Guerra, M.P. An improved protocol for intact chloroplasts and cpDNA isolation in conifers. PLoS ONE 2014, 9, e84792. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Isolation and structural analysis of chloroplast DNA. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1986; Volume 118, pp. 167–186. [Google Scholar]
- Diekmann, K.; Hodkinson, T.R.; Wolfe, K.H.; van den Bekerom, R.; Dix, P.J.; Barth, S. Complete chloroplast genome sequence of a major allogamous forage species, perennial ryegrass (Lolium perenne L.). DNA Res. 2009, 16, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, A.B.; Sharpe, R.M.; Mysayphonh, C.; Thompson, E.J.; Ward, A.D.; Lin, A. The complete chloroplast genome of tall fescue (Lolium arundinaceum; Poaceae) and comparison of whole plastomes from the family Poaceae. Am. J. Bot. 2010, 97, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Fujita, M.; Handa, H.; Murayama, S.; Uemura, M.; Kawamura, Y.; Mitsui, T.; Mikami, S.; Tozawa, Y.; Yoshinaga, T.; et al. Proteomics of the rice cell: Systematic identification of the protein populations in subcellular compartments. Mol. Genet. Genom. 2004, 271, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Scheumann, V.; Schoch, S.; Rüdiger, W. Chlorophyll b reduction during senescence of barley seedlings. Planta 1999, 209, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Führs, H.; Behrens, C.; Gallien, S.; Heintz, D.; Van Dorsselaer, A.; Braun, H.P.; Horst, W.J. Physiological and proteomic characterization of manganese sensitivity and tolerance in rice (Oryza sativa) in comparison with barley (Hordeum vulgare). Ann. Bot. 2010, 105, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Lehväslaiho, H.; Saura, A.; Lokki, J. Chloroplast DNA variation in the grass tribe Festuceae. Theor. Appl. Genet. 1987, 74, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Hu, N.; Huang, H.; Gao, J.; Zhao, Y.-J.; Gao, L.-Z. An improved chloroplast DNA extraction procedure for whole plastid genome sequencing. PLoS ONE 2012, 7, e31468. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; dePamphilis, C.W.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J.; et al. Methods for obtaining and analyzing whole chloroplast genome sequences. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2005; Volume 395, pp. 348–384. [Google Scholar]
- Takamatsu, T.; Baslam, M.; Inomata, T.; Oikawa, K.; Itoh, K.; Ohnishi, T.; Kinoshita, T.; Mitsui, T. Optimized method of extracting rice chloroplast DNA for high-quality plastome resequencing and de novo assembly. Front. Plant Sci. 2018, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Kubis, S.E.; Lilley, K.S.; Jarvis, P. Isolation and preparation of chloroplasts from Arabidopsis thaliana plants. Methods Mol. Biol. 2008, 425, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed]
- Takabe, T.; Nishimtjra, M.; Akazawa, T. Isolation of intact chloroplasts from spinach leaf by centrifugation in gradients of the modified silica “Percoll”. Agric. Biol. Chem. 1979, 43, 2137–2142. [Google Scholar] [CrossRef]
- Islam, M.S.; Studer, B.; Byrne, S.L.; Farrell, J.D.; Panitz, F.; Bendixen, C.; Møller, I.M.; Asp, T. The genome and transcriptome of perennial ryegrass mitochondria. BMC Genom. 2013, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Hand, M.L.; Spangenberg, G.C.; Forster, J.W.; Cogan, N.O.I. Plastome sequence determination and comparative analysis for members of the Lolium-Festuca grass species complex. G3 (Bethesda, Md.) 2013, 3, 607–616. [Google Scholar] [CrossRef]
- Bortiri, E.; Coleman-Derr, D.; Lazo, G.R.; Anderson, O.D.; Gu, Y.Q. The complete chloroplast genome sequence of Brachypodium distachyon: Sequence comparison and phylogenetic analysis of eight grass plastomes. BMC Res. Notes 2008, 1, 61. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety/ Accession | Common Name | Species | Type | Ploidy Status |
|---|---|---|---|---|
| Rita | Meadow fescue | F. pratensis Huds. | Bunch type | 2n = 2x = 14 |
| PI 289654 | Tetraploid fescue | F. glaucescens | Bunch type | 2n = 4x = 28 |
| PI 535747 | Atlas fescue | F. mairei St. Yves | Bunch type | 2n = 4x = 28 |
| Resolute | Tall fescue | F. arundinacea Schreb. | Mediterranean | 2n = 6x = 42 |
| Texoma MaxQ II | Tall fescue | F. arundinacea Schreb. | Continental | 2n = 6x = 42 |
| Torpedo | Tall fescue | F. arundinacea Schreb. | Rhizomatous | 2n = 6x = 42 |
| Vista | Red fescue | F. rubra ssp. rubra | Strong creeping Rhizomatous | 2n = 8x = 56 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.S.; Buttelmann, G.L.; Chekhovskiy, K.; Kwon, T.; Saha, M.C. Isolation of Intact Chloroplast for Sequencing Plastid Genomes of Five Festuca Species. Plants 2019, 8, 606. https://doi.org/10.3390/plants8120606
Islam MS, Buttelmann GL, Chekhovskiy K, Kwon T, Saha MC. Isolation of Intact Chloroplast for Sequencing Plastid Genomes of Five Festuca Species. Plants. 2019; 8(12):606. https://doi.org/10.3390/plants8120606
Chicago/Turabian StyleIslam, Md. Shofiqul, Gretta L. Buttelmann, Konstantin Chekhovskiy, Taegun Kwon, and Malay C. Saha. 2019. "Isolation of Intact Chloroplast for Sequencing Plastid Genomes of Five Festuca Species" Plants 8, no. 12: 606. https://doi.org/10.3390/plants8120606
APA StyleIslam, M. S., Buttelmann, G. L., Chekhovskiy, K., Kwon, T., & Saha, M. C. (2019). Isolation of Intact Chloroplast for Sequencing Plastid Genomes of Five Festuca Species. Plants, 8(12), 606. https://doi.org/10.3390/plants8120606