Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Experiments and Plant Sampling
2.1.1. Field Experiment 1: Study of Different Fertilization Rates on an S-Deficient Field
2.1.2. Field Experiment 2: Study of 45 Commercial Varieties before Fertilization and Flowering
2.1.3. Field Experiment 3: Study of 56 Commercial Varieties after Fertilization and Flowering
2.2. Multispecies Experiment under Controlled Conditions
2.3. SO42−, PO43−, Cl− and NO3−Analysis
2.4. S, P, Cl and N Analysis
3. Statistical Analysis
4. Results
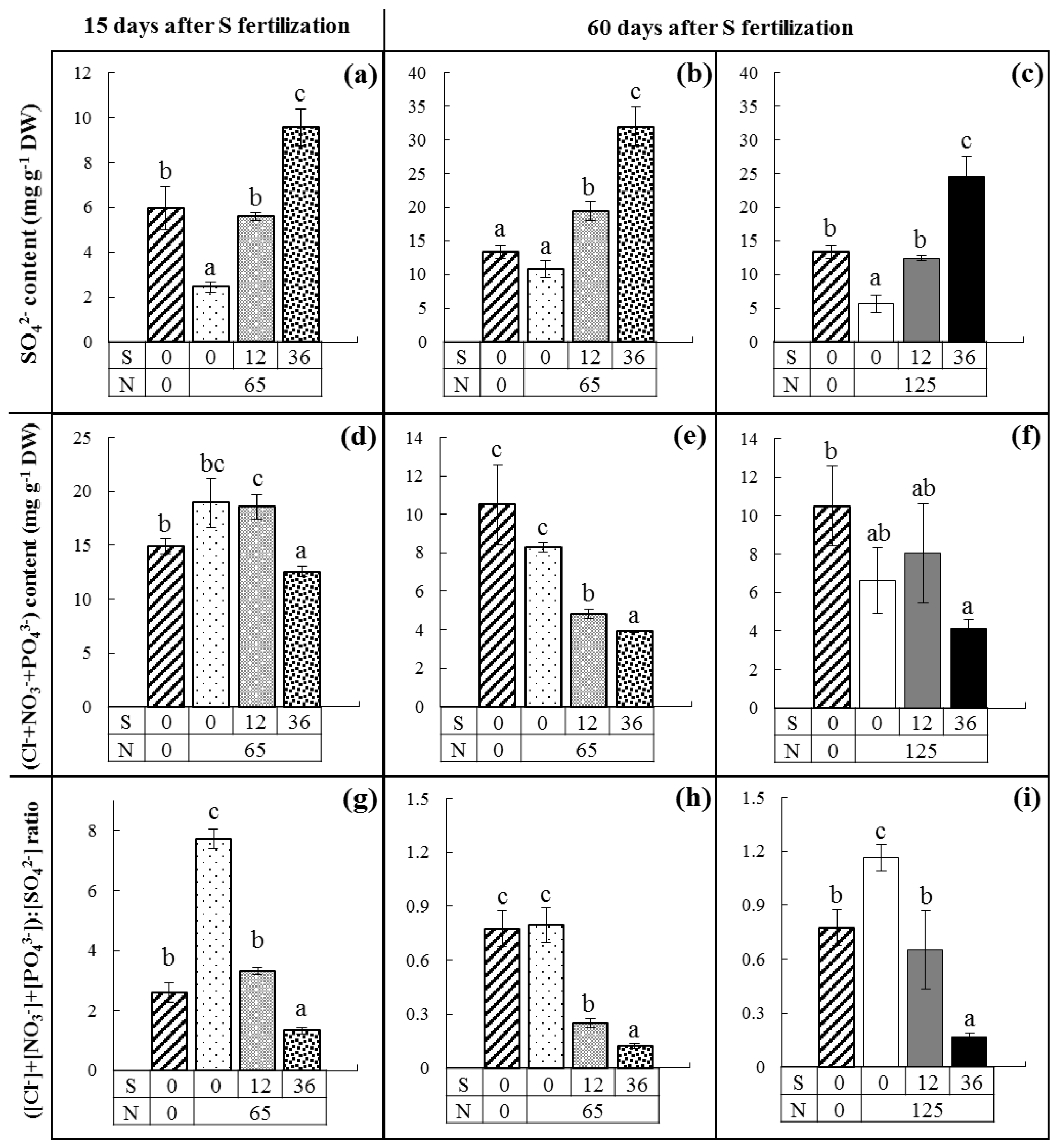
4.1. Under Field Conditions, a Decrease in the SO42− Content Was Compensated by an Increase in the (Cl−+NO3−+PO43−) Contents in Oilseed Rape Leaves Leading to an Increase in the ([Cl−]+[NO3−]+[PO43−]):[SO42−]Ratio
4.2. The ([Cl−]+[NO3−]+[PO43−]):[SO42−]Ratio Can Be Used in Other Plant Species to Detect S Deficiency under Controlled Conditions
4.3. The Simplified ([Cl]+[P]):[S] Ratio Could Be Used for an Easier and Faster Determination of S Deficiency under Field Conditions
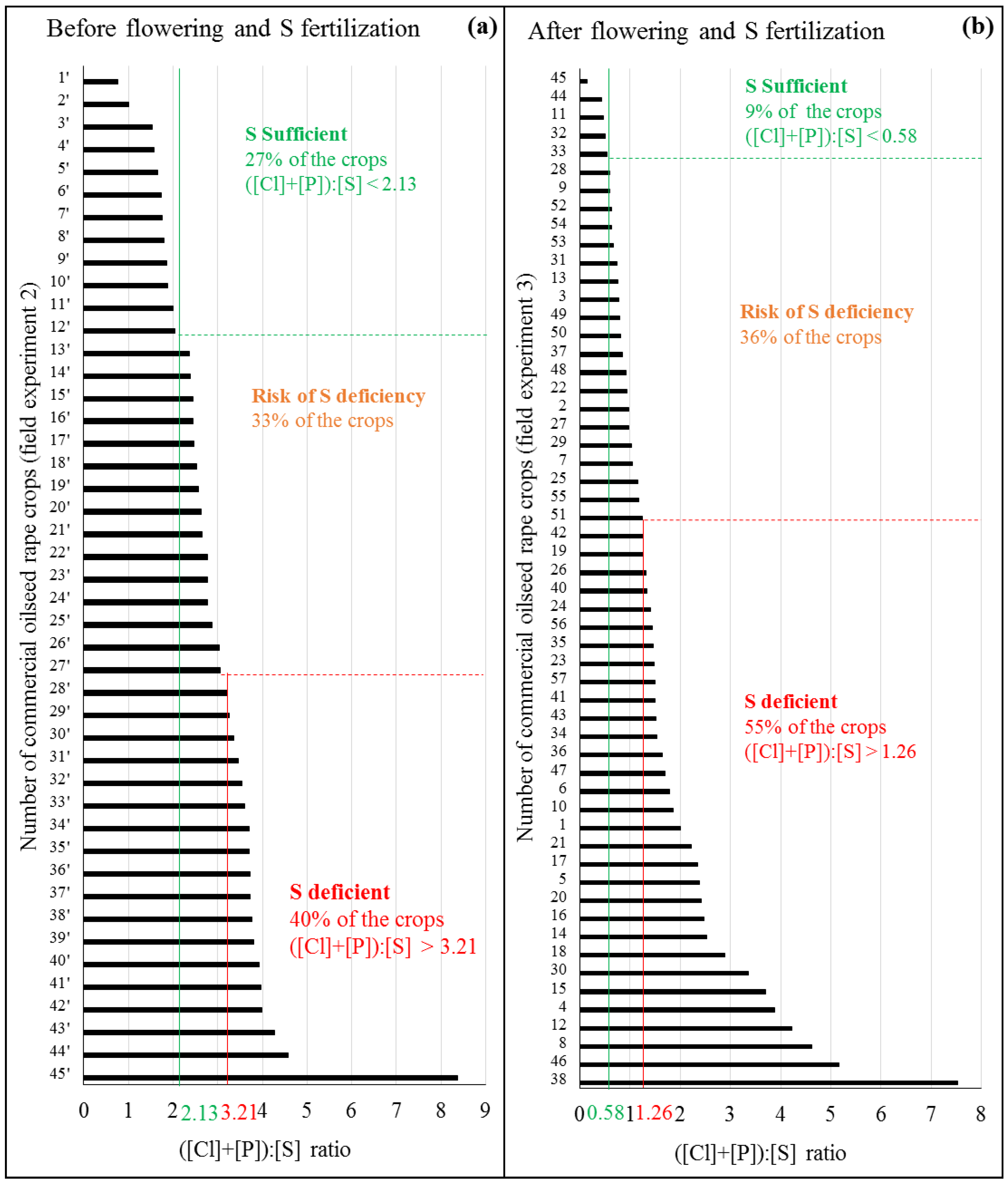
4.4. Determination of Threshold Values of the ([Cl]+[P]):[S]Ratio and Their Use on Independent Fields
5. Discussion
5.1. The ([Cl−]+[NO3−]+[PO43−]):[SO42−] Ratio as an Indicator of S Nutrition under Field Conditions
5.2. Using the ([Cl]+[P]):[S] Ratio Instead of the ([Cl−]+[NO3−]+[PO43−]):[SO42−] Ratio
5.3. Potential Thresholds of the ([Cl]+[P]):[S] Ratio
6. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| DW | dry weight |
| HPLC | high performance liquid chromatography |
| HR ICP-MS | high resolution inductively coupled plasma mass spectrometry |
| XRF | X-ray fluorescence |
References
- Janzen, H.H.; Bettany, J.R. Sulfur nutrition of rapeseed: I. Influence of fertilizer nitrogen and sulfur rates. Soil Sci. Soc. Am. J. 1984, 48, 100–107. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J. Sulphur uptake, yield responses and the interactions between nitrogen and sulphur in winter oilseed rape (Brassica napus). J. Agric. Sci. 1996, 126, 53–62. [Google Scholar] [CrossRef]
- Scherer, H.W. Sulphur in crop production. Eur. J. Agron. 2001, 14, 81–111. [Google Scholar] [CrossRef]
- Malhi, S.S.; Gan, Y.; Raney, J.P. Yield, seed quality, and sulfur uptake of oilseed crops in response to sulfur fertilization. Agron. J. 2007, 99, 570–577. [Google Scholar] [CrossRef]
- D’Hooghe, P.; Dubousset, L.; Gallardo, K.; Kopriva, S.; Avice, J.C.; Trouverie, J. Evidence for proteomic and metabolic adaptations associated with alterations of seed yield and quality in Sulfur-limited Brassica napus L. Mol. Cell. Proteom. 2014, 13, 1165–1183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Hawkesford, M.J.; McGrath, S.P. Sulphur assimilation and effects on yield and quality of wheat. J. Cereal Sci. 1999, 30, 1–17. [Google Scholar] [CrossRef]
- Blake-Kalff, M.M.A.; Zhao, F.J.; Hawkesford, M.J.; McGrath, S.P. Using plant analysis to predict yield losses caused by sulphur deficiency. Ann. Appl. Biol. 2001, 138, 123–127. [Google Scholar] [CrossRef]
- McNeill, A.M.; Eriksen, J.; Bergström, L.; Bergström, L.; Smith, K.A.; Marstorp, H.; Kirchmann, H.; Nilsson, I. Nitrogen and sulphur management: Challenges for organic sources in temperate agricultural systems. Soil Use Manag. 2005, 21, 82–93. [Google Scholar] [CrossRef]
- Grant, C.A.; Mahli, S.S.; Karamanos, R.E. Sulfur management for rapeseed. Field Crops Res. 2012, 128, 119–128. [Google Scholar] [CrossRef]
- Sarda, X.; Diquelou, S.; Abdallah, M.; Nési, N.; Cantat, O.; Le Gouee, P.; Avice, J.-C.; Ourry, A. Assessment of sulphur deficiency in commercial oilseed rape crops from plant analysis. J. Agric. Sci. 2014, 152, 616–633. [Google Scholar] [CrossRef]
- Ayala-Silva, T.; Beyl, C.A. Changes in spectral reflectance of wheat leaves in response to specific macronutrient deficiency. Adv. Space Res. 2005, 35, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Zuber, H.; Poignavent, G.; Le Signor, C.; Aimé, D.; Vieren, E.; Tadla, C.; Lugan, R.; Belghazi, M.; Labas, V.; Santoni, A.L.; et al. Legume adaptation to sulfur deficiency revealed by comparing nutrient allocation and seed traits in Medicago truncatula. Plant J. 2013, 76, 982–996. [Google Scholar] [CrossRef] [PubMed]
- Scherer, H.W. Sulfur in soils. J. Plant Nutr. Soil Sci. 2009, 172, 326–335. [Google Scholar] [CrossRef]
- Malhi, S.S.; Schoenau, J.J.; Grant, C.A. A review of sulphur fertilizer management for optimum yield and quality of canola in the Canadian Great Plains. Can. J. Plant Sci. 2005, 85, 297–307. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J. A risk assessment of sulphur deficiency in cereals using soil and atmospheric deposition data. Soil Use Manag. 1995, 11, 110–114. [Google Scholar] [CrossRef]
- Brunel-Muguet, S.; Mollier, A.; Kauffmann, F.; Avice, J.C.; Sénécal, E.; Goudier, D.; Bataillé, M.P.; Etienne, P. SuMoToRI, an ecophysiological model to predict growth and sulfur allocation and partitioning in oilseed rape (Brassica napus L.). Front. Plant Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Melsted, S.W.; Motto, H.L.; Peck, T.R. Critical plant nutrient composition values useful in interpreting plant analysis data. Agron. J. 1969, 61, 17–20. [Google Scholar] [CrossRef]
- Rasmussen, P.E.; Ramig, R.E.; Rohde, C.R. Tissue analyses guidelines for diagnosing sulfur deficiency in white wheat. Plant Soil 1977, 46, 153–163. [Google Scholar] [CrossRef]
- Spencer, K.; Freney, J.R. Assessing the sulfur status of field-grown wheat by plant analysis. Agron. J. 1980, 72, 469–472. [Google Scholar] [CrossRef]
- Scaife, A.; Burns, I.G. The sulphate-S/total S ratio in plants as an index of their sulphur status. Plant Soil 1986, 91, 61–71. [Google Scholar] [CrossRef]
- Pinkerton, A. Critical sulfur concentrations in oilseed rape (Brassica napus) in relation to nitrogen supply and to plant age. Aust. J. Exp. Agric. 1998, 38, 511–522. [Google Scholar] [CrossRef]
- Blake-Kalff, M.M.A.; Hawkesford, M.J.; Zhao, F.J.; McGrath, S.P. Diagnosing sulfur deficiency in field-grown oilseed rape (Brassica napus L.) and wheat (Triticum aestivum L.). Plant Soil 2000, 225, 95–107. [Google Scholar] [CrossRef]
- Maillard, A.; Sorin, E.; Etienne, P.; Diquélou, S.; Kopriva, A.; Kopriva, S.; Arkoun, M.; Gallardo, K.; Turner, M.; Cruz, F.; et al. Non-specific root transport of nutrient gives access to an early nutritional indicator: The case of sulfate and molybdate. PLoS ONE 2016, 11, e0166910. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Hawkesford, M.J.; Warrilow, A.G.S.; McGrath, S.P.; Clarkson, D.T. Responses of two wheat commercial to sulphur addition and diagnosis of sulphur deficiency. Plant Soil 1996, 181, 317–327. [Google Scholar] [CrossRef]
- Howarth, J.R.; Parmar, S.; Barraclough, P.B.; Hawkesford, M.J. A sulphur deficiency-induced gene, sdi1, involved in the utilization of stored sulphate pools under sulphur-limiting conditions has potential as a diagnostic indicator of sulphur nutritional status. Plant Biotechnol. J. 2009, 7, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Sorin, E.; Etienne, P.; Maillard, A.; Zamarreno, A.M.; Garcia-Mina, J.M.; Arkoun, M.; Jamois, F.; Cruz, F.; Yvin, J.C.; Ourry, A. Effect of sulphur deprivation on osmotic potential components and nitrogen metabolism in oilseed rape leaves: Identification of a new early indicator. J. Exp. Bot. 2015, 66, 6175–6189. [Google Scholar] [CrossRef] [PubMed]
- Blake-Kalff, M.M.A.; Harrison, K.R.; Hawkesford, M.J.; Zhao, F.J.; McGrath, S.P. Distribution of sulfur within oilseed rape leaves in response to sulfur deficiency during vegetative growth. Plant Physiol. 1998, 118, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Honsel, A.; Kojima, M.; Haas, R.; Frank, W.; Sakakabara, H.; Herschbach, C.; Heinz, R. Sulphur limitation and early sulphur deficiency responses in poplar: Significance of gene expression, metabolites, and plant hormones. J. Exp. Bot. 2012, 63, 1873–1893. [Google Scholar] [CrossRef] [PubMed]
- Rabe, E. Altered nitrogen metabolism under environmental stress conditions. In Handbook of Plant and Crop Stress, 2nd ed.; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 349–363. ISBN 978-1439813966. [Google Scholar]
- Tausz, M. The role of glutathione in plant response and adaptation to natural stress. In Significance of Glutathione to Plant Adaptation to the Environment; Grill, D., Tausz, M., Kok, L.J.D., Eds.; Springer: Houten, The Netherlands, 2001; pp. 101–122. ISBN 978-1402001789. [Google Scholar]
- Maynard, D.; Stewart, J.; Bettany, J. Use of plant analysis to predict sulfur deficiency in rapeseed (Brassica napus and B. campestris). Can. J. Soil Sci. 1983, 63, 387–396. [Google Scholar] [CrossRef]
- Blake-Kalff, M.M.A.; Zhao, F.J.; McGrath, S.P.; Withers, P.J.A. Development of the Malate:Sulphate Ratio Test for Sulphur Deficiency in Winter Wheat and Oilseed Rape; HGCA Project Report; No. 327; Home-Grown Cereals Authority: London, UK, 2004. [Google Scholar]
- Carver, M.F.F. Monitoring Winter Barley, Wheat, Oilseed Rape and Spring Barley for Sulphur in England and Wales to Predict Fertiliser Need; HGCA Project Report; Rep, No. 374; Home-Grown Cereals Authority: London, UK, 2005. [Google Scholar]
- Reussi, N.; Echeverría, H.; Rozas, H.S. Diagnosing sulfur deficiency in spring red wheat; plant analysis. J. Plant Nutr. 2011, 34, 573–589. [Google Scholar] [CrossRef]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.-M.; Yvin, J.C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Evans, E.J.; Bilsborrow, P.E.; Syers, J.K. Influence of sulphur and nitrogen on seed yield and quality of low glucosinolate oilseed rape (Brassica napus L.). J. Sci. Food Agric. 1993, 63, 29–37. [Google Scholar] [CrossRef]
- Dubousset, L.; Etienne, P.; Avice, J.C. Is the remobilization of S and N reserves for seed filling of winter oilseed rape modulated by sulphate restrictions occurring at different growth stages? J. Exp. Bot. 2010, 61, 4313–4324. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.; Etienne, P.; Ourry, A.; Meuriot, F. Do initial S reserves and mineral S availability alter leaf S-N mobilization and leaf senescence in oilseed rape? Plant Sci. 2011, 180, 511–520. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Day of Treatment | SO42− Content (mg g−1 DW) | (Cl−+NO3−+PO42−) Content (mg g−1 DW) | ([Cl−]+[NO3−]+[PO42−])/[SO42−] Ratio | |||
|---|---|---|---|---|---|---|---|
| +S | −S | +S | −S | +S | −S | ||
| B. napus | 0 | 24.38 ± 1.83 | 59.79 ± 7.37 | 2.32 ± 0.28 | |||
| 3 | 26.20 ± 1.23 | 15.90 ± 1.44*** | 63.65 ± 0.96 | 70.86 ± 5.68 | 2.43 ± 0.11 | 4.46 ± 0.39** | |
| 13 | 29.85 ± 0.50 | 8.05 ± 0.48*** | 62.83 ± 5.86 | 86.42 ± 4.66** | 2.10 ± 0.20 | 10.74 ± 0.17*** | |
| B. oleracea | 135 | 7.05 ± 1.22 | 0.35 ± 0.04*** | 44.9 ± 6.85 | 61.87 ± 3.27* | 6.36 ± 1.06 | 176.77 ± 40.89** |
| T. aestivum | 0 | 3.03 ± 0.09 | 63.58 ± 0.80 | 20.98 ± 0.76 | |||
| 8 | 1.94 ± 0.05 | 1.15 ± 0.03 | 59.34 ± 2.10 | 68.04 ± 0.84** | 30.58 ± 0.53 | 59.16 ± 2.36*** | |
| 16 | 2.17 ± 0.06 | 0.50 ± 0.05*** | 64.79 ± 1.29 | 70.19 ± 1.80* | 29.85 ± 0.53 | 140.38 ± 15.81*** | |
| Z. mays | 0 | 2.02 ± 0.49 | 74.74 ± 1.55 | 37.00 ± 7.16 | |||
| 5 | 3.44 ± 0.07 | 1.39 ± 0.08*** | 78.31 ± 2.82 | 91.71 ± 1.28** | 22.76 ± 1.09 | 56.33 ± 3.45*** | |
| 18 | 1.38 ± 0.05 | 0.27 ± 0.02*** | 70.38 ± 3.46 | 103.28 ± 0.79*** | 51.00 ± 2.19 | 382.51 ± 23.44*** | |
| S. lycopersicum | 75 | 63.75 ± 4.79 | 50.39 ± 2.42* | 68.84 ± 4.61 | 87.77 ± 2.44* | 1.07 ± 0.15 | 1.74 ± 0.10** |
| M. truncatula | 0 | 3.31 ± 0.16 | 25.32 ± 0.60 | 7.55 ± 0.21 | |||
| 8 | 3.60 ± 0.12 | 0.15 ± 0.01*** | 25.40 ± 1.54 | 34.27 ± 3.44* | 7.05 ± 0.48 | 228.47 ± 25.21*** | |
| 21 | 4.75 ± 0.18 | 0.26 ± 0.01*** | 25.92 ± 1.94 | 34.90 ± 2.46* | 5.49 ± 0.50 | 134.23 ± 12.48*** | |
| Before Flowering and Fertilization | S-(N) Fertilization (kg S-(N) ha−1 | After Flowering and Fertilization | |||||
|---|---|---|---|---|---|---|---|
| Plots number | Soil type | Soil S content (mg kg−1) | ([Cl]+[P])/[S] ratio | Status of plots | ([Cl]+[P])/[S] ratio | Status of plots | |
| 36′ | Compact silt | 136 | 3.72 ± 0.43 b | S deficient | 10-(166) | 1.54 ± 0.21 b** | S deficient |
| 20′ | Superficial clay-limestone | 239 | 2.64 ± 0.13 c | Risk of S deficiency | 100-(146) | 0.59 ± 0.10 a*** | S sufficient |
| 11′ | Superficial clay-limestone | 350 | 1.99 ± 0.14 a | S sufficient | 0-(110) | 1.42 ± 0.05 b** | S deficient |
| 8′ | deep clay-limestone | 668 | 1.80 ± 0.06 a | S sufficient | 15-(151) | 1.41 ± 0.24 b | S deficient |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etienne, P.; Sorin, E.; Maillard, A.; Gallardo, K.; Arkoun, M.; Guerrand, J.; Cruz, F.; Yvin, J.-C.; Ourry, A. Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves. Plants 2018, 7, 37. https://doi.org/10.3390/plants7020037
Etienne P, Sorin E, Maillard A, Gallardo K, Arkoun M, Guerrand J, Cruz F, Yvin J-C, Ourry A. Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves. Plants. 2018; 7(2):37. https://doi.org/10.3390/plants7020037
Chicago/Turabian StyleEtienne, Philippe, Elise Sorin, Anne Maillard, Karine Gallardo, Mustapha Arkoun, Jérôme Guerrand, Florence Cruz, Jean-Claude Yvin, and Alain Ourry. 2018. "Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves" Plants 7, no. 2: 37. https://doi.org/10.3390/plants7020037
APA StyleEtienne, P., Sorin, E., Maillard, A., Gallardo, K., Arkoun, M., Guerrand, J., Cruz, F., Yvin, J.-C., & Ourry, A. (2018). Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves. Plants, 7(2), 37. https://doi.org/10.3390/plants7020037