

Millennial Floristic Diversity and Land Management as Inferred from Archaeo-Palynological Research in Southern Italy

 ,
,  ,
,  ,
, 
 and
and 
Abstract
1. Introduction
Aim of the Paper
2. Results
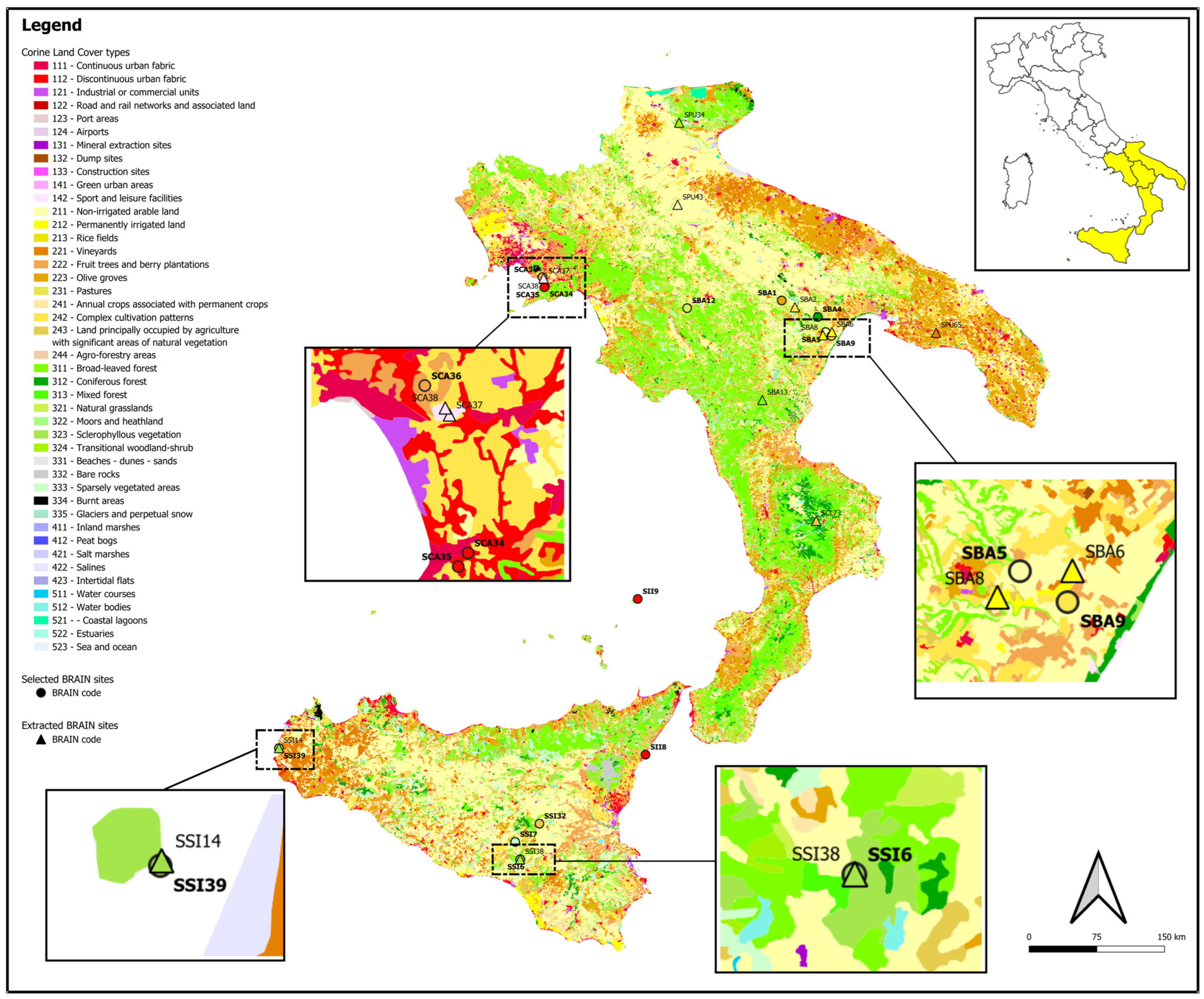
2.1. Overview of Sites in Southern Italy
2.2. Case Studies Selected from Different Regions of Southern Italy
2.2.1. Campania
2.2.2. Basilicata
2.2.3. Sicily
3. Discussion
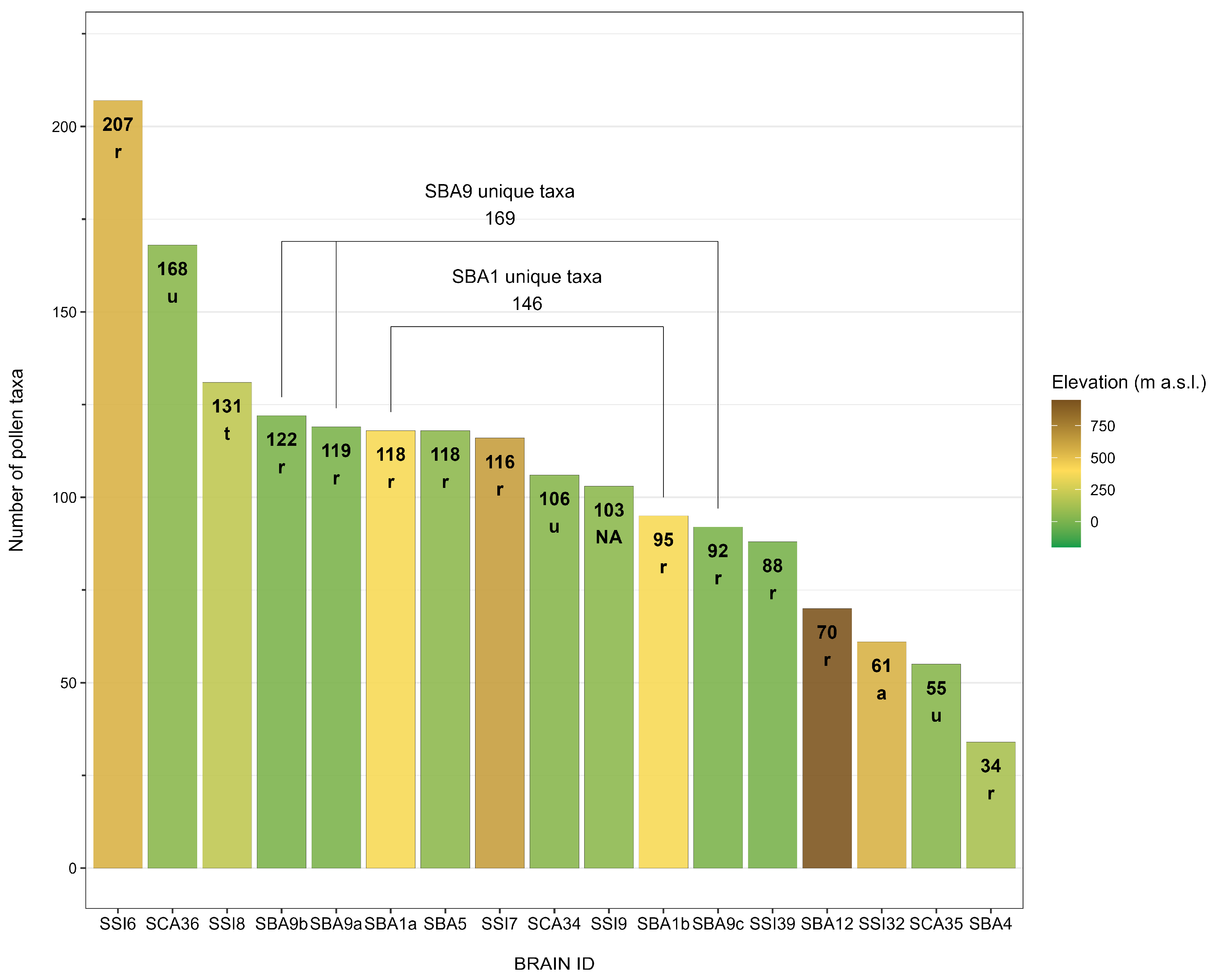
3.1. Floristic Palaeodiversity of Southern Italy
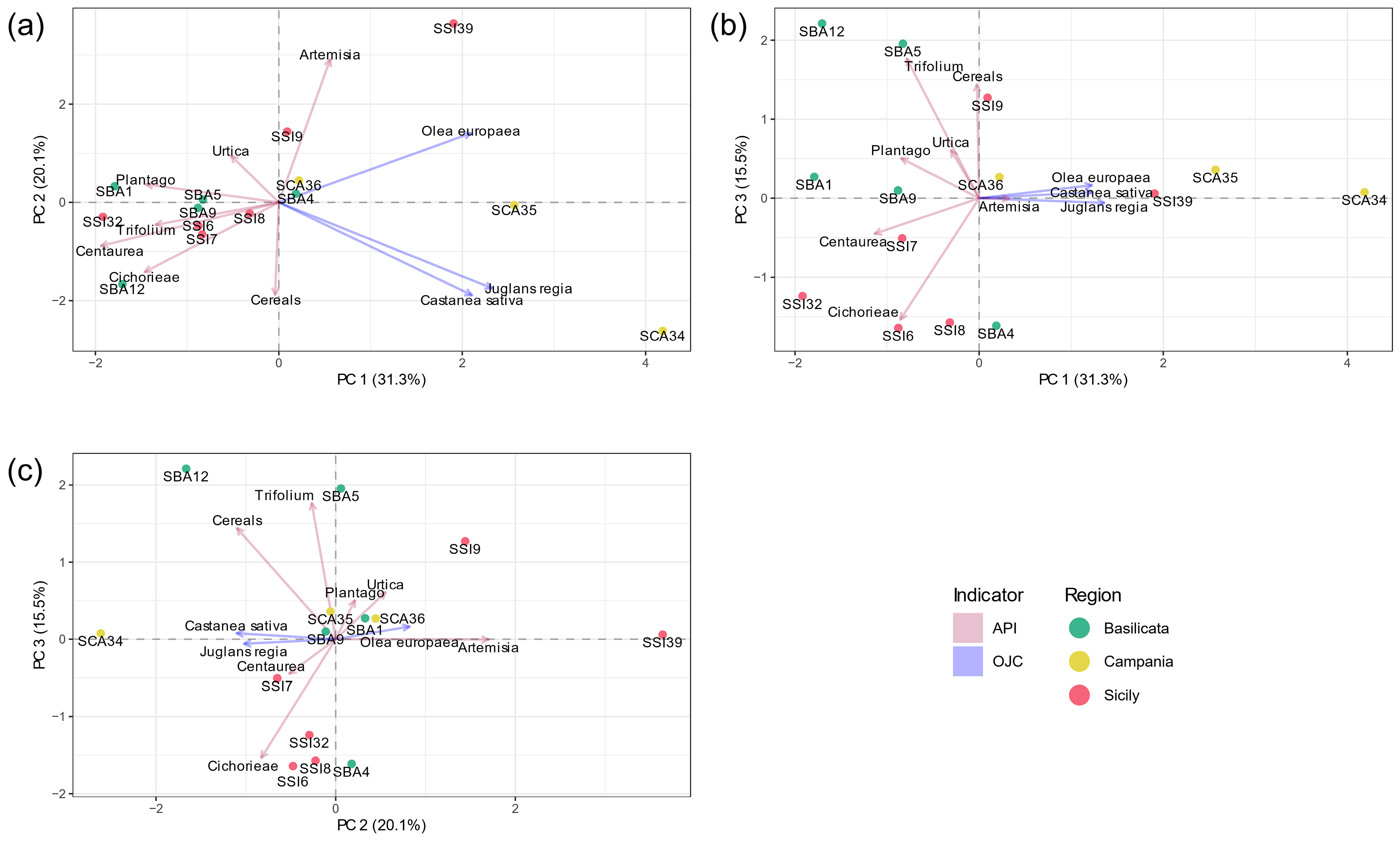
3.2. Ubiquitous Pollen Taxa in Anthropogenic Environments
3.3. Exploring the Link Between Past and Present Flora, and the Role of Morphopalynology
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peruzzi, L. Floristic inventories and collaborative approaches: A new era for checklists and floras? Plant Biosyst. 2018, 152, 177–178. [Google Scholar] [CrossRef]
- Rosati, L.; Fascetti, S.; Romano, V.A.; Potenza, G.; Lapenna, M.R.; Capano, A.; Nicoletti, P.; Farris, E.; de Lange, P.J.; Del Vico, E.; et al. New Chorological Data for the Italian Vascular Flora. Diversity 2020, 12, 22. [Google Scholar] [CrossRef]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.A.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian Vascular Flora: New Findings, Updates and Exploration of Floristic Similarities between Regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Bedini, G.; Peruzzi, L. Next Generation Floristics: A workflow to integrate novel methods in traditional floristic research. Plant Biosyst. 2022, 156, 594–597. [Google Scholar] [CrossRef]
- Carta, A.; Forbicioni, L.; Frangini, G.; Pierini, B.; Peruzzi, L. An updated inventory of the vascular flora of Elba Island (Tuscan Archipelago, Italy). Ital. Bot. 2018, 6, 1–22. [Google Scholar] [CrossRef]
- Gestri, G.; Pierini, B.; D’Antraccoli, M.; Bernardini, A.; Peruzzi, L. An updated inventory of the vascular flora of the Cerbaie hills (Tuscany, Italy). Ital. Bot. 2023, 15, 165–175. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D.; et al. A second update to the checklist of the vascular flora native to Italy. Plant Biosyst. 2024, 158, 219–296. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Roma-Marzio, F.; Carta, A.; Landi, S.; Bedini, G.; Chiarucci, A.; Peruzzi, L. Drivers of floristic richness in the Mediterranean: A case study from Tuscany. Biodivers. Conserv. 2019, 28, 1411–1429. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Jiménez-Alfaro, B.; Jandt, U.; Chytrý, M.; Field, R.; Kessler, M.; Lenoir, J.; Schrodt, F.; Wiser, S.K.; Arfin Khan, M.A.S.; et al. Global patterns of vascular plant alpha diversity. Nat. Commun. 2022, 13, 4683. [Google Scholar] [CrossRef]
- Spadaro, V.; Marino, P.; Scuderi, L.; Venturella, G.; Raimondo, F.M. Biodiversity in some populations of Crataegus (Rosaceae) from western Sicily: Description of two new species and notes on conservation and valorisation. Flora Mediterr. 2024, 34, 239–255. [Google Scholar] [CrossRef]
- Edwards, K.J.; Fyfe, R.M.; Jackson, S.T. The first 100 years of pollen analysis. Nat. Plants 2017, 3, 17001. [Google Scholar] [CrossRef]
- Luelmo-Lautenschlaeger, R.; Morales-Molino, C.; Blarquez, O.; Pérez-Díaz, S.; Sabariego-Ruiz, S.; Ochando, J.; Carrión, J.S.; Perea, R.; Fernández-González, F.; López-Sáez, J.A. Long-term vegetation history of a relict birch forest (Betula pubescens subsp. celtiberica (Rothm. & Vasc.) Rivas Mart.) in the Toledo Mountains (central Iberia). Conservation implications. Rev. Palaeobot. Palynol. 2023, 316, 104906. [Google Scholar] [CrossRef]
- Rull, V.; Burjachs, F.; Carrión, J.S.; Ejarque, A.; Fernández, S.; López-Sáez, J.A.; Luelmo-Lautenschlaeger, R.; Ochando, J.; Pérez-Díaz, S.; Revelles, J.; et al. Historical biogeography of Cannabis in the Iberian Peninsula: A probabilistic approach using palynological evidence. Perspect. Plant Ecol. Evol. Syst. 2023, 58, 125704. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M. The long history of Cannabis and its cultivation by the Romans in central Italy, shown by pollen records from Lago Albano and Lago di Nemi. Veg. Hist. Archaeobot. 2002, 11, 263–276. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; Massamba N’siala, I. A marine/terrestrial integration for mid-late Holocene vegetation history and the development of the cultural landscape in the Po Valley as a result of human impact and climate change. Veg. Hist. Archaeobot. 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Servera-Vives, G.; Miras, Y.; Riera, S.; Julià, R.; Allée, P.; Orengo, H.; Paradis-Grenouillet, S.; Palet, J.M. Tracing the land use history and vegetation dynamics in the Mont Lozère (Massif Central, France) during the last 2000 years: The interdisciplinary study case of Countrasts peat bog. Quat. Int. 2013, 353, 123–139. [Google Scholar] [CrossRef]
- Masci, L.; Liakopoulos, G.C.; Gromig, R.; Kolovos, E.; Kouli, K.; Moros, M.; Sadori, L.; Sarantis, A.; Slavin, P.; Sypiański, J. Consilience in practice: Social–ecological dynamics of the Lake Volvi region (Greece) during the last two millennia. J. Quat. Sci. 2024, 40, 459–480. [Google Scholar] [CrossRef]
- Palli, J.; Monaco, L.; Bini, M.; Cosma, E.; Giaccio, B.; Izdebski, A.; Masi, A.; Mensing, S.; Piovesan, G.; Rossi, V.; et al. The recent evolution of the salt marsh ‘Pantano Grande’ (NE Sicily, Italy): Interplay between natural and human activity over the last 3700 years. J. Quat. Sci. 2024, 39, 327–339. [Google Scholar] [CrossRef]
- Fægri, K.; Iversen, J.; Kaland, P.E.; Krzywinski, K. Textbook of Pollen Analysis, 4th ed.; The Blackburn Press: Caldwell, ID, USA, 1989. [Google Scholar]
- Mercuri, A.M.; Florenzano, A.; Clò, E.; Braga, L.; Zappa, J.; Cremaschi, M.; Zerboni, A. The precision land knowledge of the past enables tailor-made environment therapy and empathy for nature. Sci. Rep. 2025, 15, 12587. [Google Scholar] [CrossRef]
- Mercuri, A.M. Genesis and evolution of the cultural landscape in central Mediterranean: The ‘where, when and how’ through the palynological approach. Landsc. Ecol. 2014, 29, 1799–1810. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Peruzzi, L.; Conti, F.; Galasso, G.; Roma-Marzio, F.; Bartolucci, F. Floristic Richness in a Mediterranean Hotspot: A Journey across Italy. Plants 2024, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Mercuri, A.M.; Sadori, L. Mediterranean culture and climatic change: Past patterns and future trends. In The Mediterranean Sea: Its History and Present Challenges; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 507–527. [Google Scholar] [CrossRef]
- Kouli, K.; Masi, A.; Mercuri, A.M.; Florenzano, A.; Sadori, L. Regional Vegetation Histories: An Overview of the Pollen Evidence from the Central Mediterranean. Late Antiq. Archaeol. 2019, 11, 69–82. [Google Scholar] [CrossRef]
- Wagner, B.; Vogel, H.; Francke, A.; Friedrich, T.; Donders, T.; Lacey, J.H.; Leng, M.J.; Regattieri, E.; Sadori, L.; Wilke, T.; et al. Mediterranean winter rainfall in phase with African monsoons during the past 1.36 million years. Nature 2019, 573, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Cremaschi, M.; Mercuri, A.M.; Torri, P.; Florenzano, A.; Pizzi, C.; Marchesini, M.; Zerboni, A. Climate change versus land management in the Po Plain (Northern Italy) during the Bronze Age: New insights from the VP/VG sequence of the Terramara Santa Rosa di Poviglio. Quat. Sci. Rev. 2016, 136, 153–172. [Google Scholar] [CrossRef]
- Cremaschi, M.; Mercuri, A.M.; Benatti, A.; Bosi, G.; Brandolini, F.; Clò, E.; Florenzano, A.; Furia, E.; Mariani, G.S.; Mazzanti, M.; et al. The SUCCESSO-TERRA Project: A Lesson of Sustainability from the Terramare Culture, Middle Bronze Age of the Po Plain (Northern Italy). IANSA 2018, 9, 221–229. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A.; Burjachs, F.; Giardini, M.; Kouli, K.; Masi, A.; Picornell-Gelabert, L.; Revelles, J.; Sadori, L.; Servera-Vives, G.; et al. From influence to impact: The multifunctional land use in Mediterranean prehistory emerging from palynology of archaeological sites (8.0–2.8 ka BP). Holocene 2019, 29, 830–846. [Google Scholar] [CrossRef]
- Stephens, L.; Fuller, D.; Boivin, N.; Rick, T.; Gauthier, N.; Kay, A.; Marwick, B.; Armstrong, C.G.; Barton, C.M.; Denham, T.; et al. Archaeological assessment reveals Earth’s early transformation through land use. Science 2019, 365, 897–902. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A. The long-term perspective of human impact on landscape for environmental change (LoTEC) and sustainability: From botany to the interdisciplinary approach. Sustainability 2019, 11, 413. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E.; Torri, P. Anthropogenic Pollen Indicators (API) from archaeological sites as local evidence of human-induced environments in the Italian peninsula. Ann. Bot. 2013, 3, 143–153. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E. Olea, Juglans and Castanea: The OJC group as pollen evidence of the development of human-induced environments in the Italian peninsula. Quatern. Int. 2013, 303, 24–42. [Google Scholar] [CrossRef]
- Fægri, K.; Iversen, J. Textbook of Pollen Analysis; Munksgaard: Copenhagen, Denmark, 1964. [Google Scholar]
- Favre, E.; Escarguel, G.; Suc, J.-P.; Vidal, G.; Thévenod, L. A contribution to deciphering the meaning of AP/NAP with respect to vegetation cover. Rev. Palaeobot. Palynol. 2008, 148, 13–35. [Google Scholar] [CrossRef]
- Vannière, B.; Colombaroli, D.; Chapron, E.; Leroux, A.; Tinner, W.; Magny, M. Climate versus human-driven fire regimes in Mediterranean landscapes: The Holocene record of Lago dell’Accesa (Tuscany, Italy). Quat. Sci. Rev. 2008, 27, 1181–1196. [Google Scholar] [CrossRef]
- Vannière, B.; Colombaroli, D.; Power, M.J. Power fire history of an inhabited Earth: Experiences from the PAGES global paleofire working Group. PAGES Mag. 2021, 29, 24–26. [Google Scholar] [CrossRef]
- Sweeney, L.; Harrison, S.P.; Vander Linden, M. Assessing anthropogenic influence on fire history during the Holocene in the Iberian Peninsula. Quat. Sci. Rev. 2022, 287, 107562. [Google Scholar] [CrossRef]
- Furia, E.; Clò, E.; Florenzano, A.; Mercuri, A.M. Human-induced fires and land use driven changes in tree biodiversity on the northern Tyrrhenian coast. Quat. Int. 2024, 705, 37–52. [Google Scholar] [CrossRef]
- Peyron, O.; Magny, M.; Goring, S.; Joannin, S.; de Beaulieu, J.-L.; Brugiapaglia, E.; Sadori, L.; Garfi, G.; Kouli, K.; Ioakim, C.; et al. Contrasting patterns of climatic changes during the Holocene across the Italian Peninsula reconstructed from pollen data. Clim. Past 2013, 9, 1233–1252. [Google Scholar] [CrossRef]
- Sadori, L.; Ortu, E.; Peyron, O.; Zanchetta, G.; Vannière, B.; Desmet, M.; Magny, M. The last 7 millennia of vegetation and climate changes at Lago di Pergusa (central Sicily, Italy). Clim. Past 2013, 9, 1969–1984. [Google Scholar] [CrossRef]
- Mensing, S.A.; Tunno, I.; Sagnotti, L.; Florindo, F.; Noble, P.; Archer, C.; Zimmerman, S.; Javier Pavón-Carrasco, F.; Cifani, G.; Passigli, S.; et al. 2700 years of Mediterranean environmental change in central Italy: A synthesis of sedimentary and cultural records to interpret past impacts of climate on society. Quat. Sci. Rev. 2015, 116, 72–94. [Google Scholar] [CrossRef]
- Mensing, S.; Tunno, I.; Cifani, G.; Passigli, S.; Noble, P.; Archer, C.; Piovesan, G. Human and climatically induced environmental change in the Mediterranean during the Medieval Climate Anomaly and Little Ice Age: A case from central Italy. Anthropocene 2016, 15, 49–59. [Google Scholar] [CrossRef]
- Sadori, L.; Giraudi, C.; Masi, A.; Magny, M.; Ortu, E.; Zanchetta, G.; Izdebski, A. Climate, environment and society in southern Italy during the last 2000 years. A review of the environmental, historical and archaeological evidence. Quat. Sci. Rev. 2016, 136, 173–188. [Google Scholar] [CrossRef]
- Di Rita, F.; Lirer, F.; Bonomo, S.; Cascella, A.; Ferraro, L.; Florindo, F.; Insinga, D.D.; Lurcock, P.C.; Margaritelli, G.; Petrosino, P.; et al. Late Holocene forest dynamics in the Gulf of Gaeta (central Mediterranean) in relation to NAO variability and human impact. Quat. Sci. Rev. 2018, 179, 137–152. [Google Scholar] [CrossRef]
- Di Rita, F.; Molisso, F.; Sacchi, M. Late Holocene environmental dynamics, vegetation history, human impact, and climate change in the Ancient Literna Palus (Lago Patria; Campania, Italy). Rev. Palaeobot. Palynol. 2018, 258, 48–61. [Google Scholar] [CrossRef]
- Michelangeli, F.; Di Rita, F.; Celant, A.; Tisnérat-Laborde, N.; Lirer, F.; Magri, D. Three Millennia of Vegetation, Land-Use, and Climate Change in SE Sicily. Forests 2022, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Mercuri, A.M.; Sadori, L.; Uzquiano Ollero, P. Mediterranean and north-African cultural adaptations to mid-Holocene environmental and climatic changes. Holocene 2011, 21, 189–206. [Google Scholar] [CrossRef]
- Roberts, N.; Eastwood, W.J.; Kuzucuoğlu, C.; Fiorentino, G.; Caracuta, V. Climatic, vegetation and cultural change in the eastern Mediterranean during the mid-Holocene environmental transition. Holocene 2011, 21, 147–162. [Google Scholar] [CrossRef]
- Roberts, N.; Woodbridge, J.; Palmisano, A.; Bevan, A.; Fyfe, R.; Shennan, S. Mediterranean landscape change during the Holocene: Synthesis, comparison and regional trends in population, land cover and climate. Holocene 2019, 29, 923–937. [Google Scholar] [CrossRef]
- Stoddart, S.; Woodbridge, J.; Palmisano, A.; Mercuri, A.M.; Mensing, S.A.; Colombaroli, D.; Sadori, L.; Magri, D.; Di Rita, F.; Giardini, M.; et al. Tyrrhenian central Italy: Holocene population and landscape ecology. Holocene 2019, 29, 761–775. [Google Scholar] [CrossRef]
- Berglund, B.E.; Gaillard, M.J.; Björkman, L.; Persson, T. Long-term changes in floristic diversity in southern Sweden: Palynological richness, vegetation dynamics and land-use. Veget. Hist. Archaeobot. 2008, 17, 573–583. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Felde, V.A.; Seddon, A.W.R. Biodiversity trends within the Holocene. Holocene 2016, 26, 994–1001. [Google Scholar] [CrossRef]
- Clò, E.; Furia, E.; Florenzano, A.; Mercuri, A.M. Flora-vegetation history and land use in Medieval Tuscany: The palynological evidence of a local biodiversity heritage. Quat. Int. 2024, 705, 1–15. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Bhatta, K.P.; Felde, V.A.; Flantua, S.G.A.; Mottl, O.; Haberle, S.G.; Herbert, A.; Hooghiemstra, H.; Birks, H.H.; Grytnes, J.-A.; et al. Approaches to pollen taxonomic harmonisation in Quaternary palynology. Rev. Palaeobot. Palynol. 2023, 319, 104989. [Google Scholar] [CrossRef]
- Dimbleby, G.W. The Palynology of Archaeological Sites; Academic Press: London, UK, 1985. [Google Scholar]
- Jones, J.; Tinsley, H.; Brunning, R. Methodologies for Assessment of the State of Preservation of Pollen and Plant Macrofossil Remains in Waterlogged Deposits. Environ. Archaeol. 2007, 12, 71–86. [Google Scholar] [CrossRef]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell: London, UK, 1991. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de Botanique Historique et Palynologie: Marseille, France, 1992. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete; Verlag Dr. Friedrich Pfeil: München, Germany, 2015. [Google Scholar]
- Mercuri, A.M.; Clò, E.; Zappa, J.; Bosi, G.; Furia, E.; Ricucci, C.; Di Lena, M.; Camerini, F.; Florenzano, A. BRAIN—Holocene archaeo-data for assessing plant-cultural diversity in Italy and other Mediterranean region. Sci. Data 2024, 11, 520. [Google Scholar] [CrossRef] [PubMed]
- Mercuri, A.M.; Florenzano, A.; Massamba N’siala, I.; Olmi, L.; Roubis, D.; Sogliani, F. Pollen from archaeological layers and cultural landscape reconstruction: Case studies from the Bradano Valley (Basilicata, southern Italy). Plant Biosyst. 2010, 144, 888–901. [Google Scholar] [CrossRef]
- Florenzano, A. Evoluzione di un Paesaggio Mediterraneo Nella Ricostruzione Archeoambientale di siti Lucani. Ph.D. Thesis, Università di Modena e Reggio Emilia, Modena, Italy, 2013. [Google Scholar]
- Florenzano, A.; Mercuri, A.M. Dal polline nei sedimenti alla ricostruzione del paesaggio e dell’economia di Torre di Satriano. In Segni del Potere. Oggetti di Lusso dal Mediterraneo nell’Appennino Lucano di età Arcaica. Catalogue; Osanna, M., Vullo, M., Eds.; Osanna Edizioni: Venosa, Italy, 2013; pp. 163–168. [Google Scholar]
- Florenzano, A.; Mercuri, A.M.; Carter, J.C. Economy and environment of the Greek colonial system in southern Italy: Pollen and NPPs evidence of grazing from the rural site of Fattoria Fabrizio (VI-IV cent. BC.; Metaponto, Basilicata). Ann. Bot. 2013, 3, 173–181. [Google Scholar] [CrossRef]
- Florenzano, A. Chapter 7. Archaeobotany at Fattoria Fabrizio. In The Chora of Metaponto 5: A Greek Farmhouse at Ponte Fabrizio; Lanza Catti, E., Swift, K., Carter, J.C., Eds.; University of Texas Press: Austin, TX, USA, 2014; pp. 113–138. [Google Scholar]
- Florenzano, A. Chapter 8. Archaeobotanical Analysis. In The Chora of Metaponto 6: A Greek Settlement at Sant’Angelo Vecchio; Silvestrelli, F., Edlund-Berry, I.E.M., Eds.; University of Texas Press: Austin, TX, USA, 2016; pp. 159–171. [Google Scholar]
- Florenzano, A.; Mercuri, A.M. Pollen evidence and the reconstruction of plant landscape of the Pantanello area from the 7th to the 1st century BC. In The Chora of Metaponto 7: A Greek Sanctuary at Pantanello; Carter, J.C., Swift, K., Eds.; University of Texas Press: Austin, TX, USA, 2018; pp. 435–446. [Google Scholar]
- Florenzano, A. The history of pastoral activities in S Italy inferred from Palynology: A long-term perspective to support biodiversity awareness. Sustainability 2019, 11, 404. [Google Scholar] [CrossRef]
- Florenzano, A.; Zerboni, A.; Carter, J.C.; Clò, E.; Mariani, G.S.; Mercuri, A.M. Environmental and land-use changes in a Mediterranean landscape: Palynology and geoarchaeology at ancient Metapontum (Pantanello, Southern Italy). Quat. Int. 2022, 635, 105–124. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Terranova, F.; Torri, P.; Trevisan Grandi, G.; Montecchi, M.C.; Olmi, L. The Greek-Roman Theatre of Taormina: Pollen and microanthracological data for the proposal of an “Historical Green Park”. In The Archaeology of Crop Fields and Gardens; Morel, J.P., Tresserras Juan, J., Matamala, J.C., Eds.; Edipuglia: Bari, Italy, 2006; pp. 161–174. [Google Scholar]
- Terranova, F.; Accorsi, C.A.; Bandini Mazzanti, M.; Mercuri, A.M.; Torri, P.; Manicardi, E.; Montecchi, M.C.; Olmi, L.; Rinaldi, R.; Valenti, V.; et al. Indagini archeopalinologiche in Sicilia a Taormina, Piazza Armerina e Mozia. In Scienza e Patrimonio Culturale nel Mediterraneo. Diagnostica e Conservazione Esperienze e Proposte per una Carta del Rischio: Atti del 3° Convegno Internazionale di Studi per la Materia e i Segni della Storia (Palermo, 12–21 Ottobre 2007); Regione Siciliana, Assessorato dei Beni Culturali, Ambientali e Della Pubblica Istruzione, Dipartimento dei beni Culturali, Ambientali, e Dell’Educazione Permanente: Palermo, Italy, 2009; pp. 184–194. [Google Scholar]
- Montecchi, M.C. Indagini Archeopalinologiche e Microantracologiche Nell’Insediamento Medievale Nell’Area Della Villa del Casale di Piazza Armerina (Enna), con Dati Pre- e Post-Medievali. Ph.D. Thesis, Università di Ferrara, Ferrara, Italy, 2010. [Google Scholar]
- Mercuri, A.M.; Montecchi, M.C.; Florenzano, A.; Rattighieri, E.; Torri, P.; Dallai, D.; Vaccaro, E. The Late Antique plant landscape in Sicily: Pollen from the agro-pastoral Villa del Casale—Philosophiana system. Quat. Int. 2019, 499, 24–34. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Cannavò, V.; Clò, E.; Di Renzoni, A.; Florenzano, A.; Rattighieri, E.; Levi, S.T. Palynology of San Vincenzo-Stromboli: Interdisciplinary perspective for the diachronic palaeoenvironmental reconstruction of an island of Sicily. J. Archaeol. Sci. Rep. 2020, 30, 102235. [Google Scholar] [CrossRef]
- Moricca, C.; Nigro, L.; Pasta, S.; Cappella, F.; Spagnoli, F.; Sadori, L. Cultural landscape and plant use at the Phoenician Motya (Western Sicily, Italy) inferred by a disposal pit. Veg. Hist. Archaeobot. 2021, 30, 815–829. [Google Scholar] [CrossRef]
- Langgut, D.; Gleason, K.; Howe, T.N.; Korman, Y.; Berger, A. The Contribution of Palynology to the Reconstruction of Villa Gardens at Roman Stabiae. CARMEL 2024, 2, 1–26. [Google Scholar]
- Florenzano, A.; Torri, P.; Bosi, G.; Clò, E.; Caprio, P.; Mercuri, A.M. Snapshots from the past: Biodiversity of the Vesuvian area before AD 79 from new archaeopalynological studies. Quat. Int. 2025, 719, 109669. [Google Scholar] [CrossRef]
- Accorsi, C.A.; Bandini Mazzanti, M.; Mercuri, A.M. Analisi palinologiche. In L’insediamento Preistorico di Terragne (Manduria-Taranto). Nuovi dati sul Processo di Neolitizzazione nel Sud-Est Italiano; Gorgoglione, M.A., Di Lernia, S., Fiorentino, G., Eds.; Regione Puglia-C.R.S.E.C. TA/55: Manduria, Italy, 1995; pp. 185–198. [Google Scholar]
- Accorsi, C.A.; Bandini Mazzanti, M.; Marchesini, M.; Marvelli, S. Ricerche archeoambientali nella Daunia antica. Dati pollinici sull’insediamento di Arpi e sulla villa romana di Ascoli Satriano. In Agricoltura e Commerci Nell’italia Antica; Quilici, L., Quilici Gigli, S., Eds.; L’Erma di Bretschneider: Roma, Italy, 1995; pp. 103–113. [Google Scholar]
- Accorsi, C.A.; Bandini Mazzanti, M.; Fiorentino, G.; Gorgoglione, A.M.; Mercuri, A.M. Archaeological and archaeobotanical data on the Mesolithic/Ancient-Medium Neolithic site of Terragne (Taranto, Southern Italy, 96 m a.s.l., 40°24′ N 17°38′ E). In Science and Technology for the Safeguard of Cultural Heritage in the Mediterranean Basin; CNR: Catania, Italy; Palermo, Italy, 1997; pp. 1521–1527. [Google Scholar]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Trevisan Grandi, G. Il paesaggio vegetale di Jure Vetere prima e durante la vita del monastero medievale sulla base dei primi dati pollinici. In Jure Vetere. Ricerche Archeologiche Nella Prima Fondazione Monastica di Giocchino da Fiore (Indagini 2001–2005); Fonseca, C.D., Roubis, D., Sogliano, F., Eds.; Rubettino Editore: Soveria Mannelli, Italy, 2007; pp. 269–287. [Google Scholar]
- Roubis, D.; Sogliani, F.; Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Florenzano, A.; Massamba N’siala, I. Exploiting a monastic territory: A multidisciplinary approach using GIS and pollen analysis to study the evolution of medieval landscape of the Jure Vetere monastery (Calabria, Italy). In Plants and Culture: Seeds of the Cultural Heritage of Europe; Morel, J.P., Mercuri, A.M., Eds.; Edipuglia: Bari, Italy, 2009; pp. 107–120. [Google Scholar]
- European Environment Agency (EEA). Corine Land Cover 2018 (CLC2018) Dataset; European Environment Agency: Copenhagen, Denmark, 2019. Available online: https://land.copernicus.eu/en/products/corine-land-cover (accessed on 1 March 2025).
- Costantini, L.; Costantini Biasini, L. Archaeobotanical Investigations at Pantanello. In The Chora of Metaponto 7: A Greek Sanctuary at Pantanello; Carter, J.C., Swift, K., Eds.; University of Texas Press: Austin, TX, USA, 2018; pp. 371–427. [Google Scholar]
- Levi, S.T.; Ayala, G.; Bettelli, M.; Brunelli, D.; Cannavò, V.; Di Renzoni, A.; Ferranti, F.; Lugli, S.; Martinelli, M.C.; Mercuri, A.M.; et al. Archaeological and volcanological investigation at Stromboli, Aeolian Islands, Italy. Antiquity 2014, 88, 342. [Google Scholar]
- Manicardi, E. Primi Dati Archeopalinologici e Microantracologici Sulla Strada Fenicio-Punica Sommersa di Mozia-S. Pantaleo (Trapani). Master’s Thesis, Università degli Studi di Modena e Reggio Emilia, Modena, Italy, 2007. [Google Scholar]
- Berglund, B.E.; Ralska-Jasiewiczowa, M. Pollen analysis and pollen diagrams. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; John Wiley & Sons Inc.: Chichester, UK, 1986; pp. 455–484. [Google Scholar]
- Mariotti Lippi, M. Charred and shrunken pollen grains as a result of special depositional conditions in the Roman age Vesuvian area. Quat. Int. 2024, 711, 59–67. [Google Scholar] [CrossRef]
- Florenzano, A.; Mercuri, A.M.; Pederzoli, A.; Torri, P.; Bosi, G.; Olmi, L.; Rinaldi, R.; Bandini Mazzanti, M. The significance of intestinal parasite remains in pollen samples from Mediaeval pits in the Piazza Garibaldi of Parma, Emilia-Romagna, Northern Italy. Geoarchaeology 2012, 27, 34–47. [Google Scholar] [CrossRef]
- Bowes, K. The Roman Peasant Project 2009–2014: Excavating the Roman Rural Poor; University Museum Monograph 154, 2 vols.; University of Pennsylvania Press: Philadelphia, PA, USA, 2021; p. 824. [Google Scholar]
- Behre, K.-E. Anthropogenic Indicators in Pollen Diagrams; A.A. Balkema: Rotterdam, The Netherlands, 1986. [Google Scholar]
- Mazier, F.; Galop, D.; Gaillard, M.J.; Rendu, C.; Cugny, C.; Legaz, A.; Peyron, O.; Buttler, A. Multidisciplinary Approach to Reconstructing Local Pastoral Activities: An Example from the Pyrenean Mountains (Pays Basque). Holocene 2009, 19, 171–188. [Google Scholar] [CrossRef]
- Deza-Araujo, M.; Morales-Molino, C.; Conedera, M.; Henne, P.D.; Krebs, P.; Hinz, M.; Heits, C.; Hafner, A.; Tinner, W. A new indicator approach to reconstruct agricultural land use in Europe from sedimentary pollen assemblages. Quat. Sci. Rev. 2022, 599, 111051. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Servera-Vives, G.; Mus Amezquita, M.; Snitker, G.; Florenzano, A.; Torri, P.; Ruiz, M.; Mercuri, A.M. Human-Impact Gradients through Anthropogenic Pollen Indicators in a Mediterranean Mosaic Landscape (Balearic Islands). Sustainability 2023, 15, 8807. [Google Scholar] [CrossRef]
- Clarke, G.C.S.; Punt, W.; Hoen, P.P. Ranunculaceae. Rev. Palaeobot. Palynol. 1991, 69, 117–271. [Google Scholar] [CrossRef]
- Mazier, F. Modelisation de la Relation Entre Pluie Pollinique Actuelle, Végetation et Pratiques Pastorales en Moyenne Montagne (Pyrenees et Jura): Application Pour L’interprétation des Données Polliniques Fossils; U.F.R. Sciences et Téchniques, Université de Franche Comté,: Besançon, France, 2007. [Google Scholar]
- Portal to the Flora of Italy. Available online: http://dryades.units.it/floritaly (accessed on 23 March 2025).
- Conti, F.; Abbate, G.; Alessandrini, A.; Blasi, C. An Annotated Checklist of the Italian Vascular Flora; Palombi e Partner S.r.l.: Roma, Italy, 2005. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora D’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017–2019. [Google Scholar]
- Birks, H.H.; Birks, H.J.B.; Kaland, P.E.; Moe, D. The Cultural Landscape: Past, Present and Future; Cambridge University Press: Cambridge, UK, 2004; p. 540. [Google Scholar]
- Sadori, L.; Zanchetta, G.; Giardini, M. Last Glacial to Holocene palaeoenvironmental evolution at Lago di Pergusa (Sicily, Southern Italy) as inferred by pollen, microcharcoal, and stable isotopes. Quat. Int. 2008, 181, 4–14. [Google Scholar] [CrossRef]
- Ruiz, I.; Sanz-Sánchez, M.J. Effects of Historical Land-Use Change in the Mediterranean Environment. Sci. Total Environ. 2020, 732, 139315. [Google Scholar] [CrossRef]
- Kouli, K. Plant landscape and land use at the Neolithic Lake settlement of Dispilió (Macedonia, northern Greece). Plant Biosyst. 2015, 149, 195–204. [Google Scholar] [CrossRef]
- Speciale, C.; Giannitrapani, E.; Mercuri, A.M.; Florenzano, A.; Sadori, L.; Combourieu-Nebout, N. The Establishment of the Agricultural Landscape of Central Sicily Between the Middle Neolithic and the Beginning of the Iron Age. Hum. Ecol. 2024, 52, 229–253. [Google Scholar] [CrossRef]
- Langgut, D.; Lev-Yadun, S.; Finkelstein, I. The impact of olive orchard abandonment and rehabilitation on pollen signature: An experimental approach to evaluating fossil pollen data. Ethnoarchaeology 2014, 6, 121–135. [Google Scholar] [CrossRef]
- Dimbleby, G.W. Pollen analysis of soil samples from the A.D. 79 level: Pompeii, Oplontis and Boscoreale. In The Natural History of Pompeii; Jashemski, F.W., Meyer, F.G., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 181–189. [Google Scholar]
- Mariotti Lippi, M.; Bellini, C. Unusual palynological evidence from gardens and crop fields of ancient Pompeii (Italy). In The Archaeology of Crop Fields and Gardens. Proceedings of the 1st Conference on “Crop Fields and Gardens Archaeology”, Barcelona, Spain, 1–3 June 2006; Morel, J.P., Tresserras, J., Matamala, J.C., Eds.; Edipuglia: Bari, Italy, 2006; pp. 153–159. [Google Scholar]
- Langgut, D.; Cheddadi, R.; Carrión, J.S.; Cavanagh, M.; Colombaroli, D.; Eastwood, W.J.; Greenberg, R.; Litt, T.; Mercuri, A.M.; Miebach, A.; et al. The origin and spread of olive cultivation in the Mediterranean Basin. Holocene 2019, 29, 902–922. [Google Scholar] [CrossRef]
- Santeramo, R.; Pallecchi, S.; Montanari, C.A. Wood Use and Forest Resource Management at Pompeii: Anthracological Analyses in the Area of Regio VII, Insula 14. Quat. Int. 2025, 729, 109782. [Google Scholar] [CrossRef]
- Russo Ermolli, E.; Ruello, M.R.; Cicala, L.; Di Lorenzo, H.; Molisso, F.; Pacciarelli, M. An 8300-yr record of environmental and cultural changes in the Sant’Eufemia Plain (Calabria, Italy). Quat. Int. 2018, 483, 39–56. [Google Scholar] [CrossRef]
- Kaniewski, D.; Marriner, N.; Terral, J.-F.; Besnard, G.; Tsitsou, L.; Topsakal, J.; Morhange, C.; Otto, T.; Luce, F.; Cheddadi, R. Olive production in the 21st century will be threatened by water stress and declining solar activity. Commun. Earth Environ. 2025, 6, 268. [Google Scholar] [CrossRef]
- Palli, J.; Fiolna, S.; Bini, M.; Cappella, F.; Izdebski, A.; Masi, A.; Mensing, S.; Nigro, L.; Piovesan, G.; Sadori, L.; et al. The human-driven ecological success of olive trees over the last 3700 years in the Central Mediterranean. Quat. Sci. Rev. 2025, 356, 109313. [Google Scholar] [CrossRef]
- Barone Lumaga, M.R.; Russo Ermolli, E.; Menale, B.; Vitale, S. Exine morphometric analysis as a new tool for Citrus species identification: A case study from Oplontis (Vesuvius Area, Italy). Veg. His. Archaeobot. 2020, 29, 671–680. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Torri, P.; Florenzano, A.; Clò, E.; Mariotti Lippi, M.; Sgarbi, E.; Bignami, C. Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio). Sustainability 2021, 13, 2287. [Google Scholar] [CrossRef]
- Tecchiati, U.; Salzani, P.; Gulino, F.; Proserpio, B.; Reggio, C.; Putzolu, C.; Rattighieri, E.; Clò, E.; Mercuri, A.M.; Florenzano, A. Palaeoenvironment, Settlement, and Land Use in the Late Neolithic—Bronze Age Site of Colombare di Negrar di Valpolicella (N Italy, On-Site). Quaternary 2022, 5, 50. [Google Scholar] [CrossRef]
- Servera-Vives, G.; Ricucci, C.; Snitker, G. OLEAtool: An open-source software for morphopalynological research in Olea europaea L. pollen. Open Res. Eur. 2024, 3, 29. [Google Scholar] [CrossRef]
- Montecchi, M.C.; Mercuri, A.M. When palynology meets classical archaeology: The Roman and medieval landscapes at the Villa del Casale di Piazza Armerina, UNESCO site in Sicily. Archaeol. Anthropol. Sci. 2018, 10, 743–757. [Google Scholar] [CrossRef]
- Rosati, L.; Masi, A.; Giardini, M.; Marignani, M. Under the shadow of a big plane tree: Why Platanus orientalis should be considered an archaeophyte in Italy. Plant Biosyst. 2015, 149, 185–194. [Google Scholar] [CrossRef]
- Raimondo, F.M.; Gianguzzi, L.; Ilardi, V. Inventario delle specie “a rischio” nella flora vascolare nativa della Sicilia. Quad. Bot. Ambient. E Appl. 1994, 3, 65–132. [Google Scholar]
- Barone, G.; Cirlincione, F.; Di Gristina, E.; Domina, G.; Gianguzzi, L.; Mirabile, G.; Naselli-Flores, L.; Raimondo, F.M.; Venturella, G. An analysis of botanical studies of vascular plants from Italian wetlands. Ital. Bot. 2022, 14, 45–60. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System; Open-Source Geospatial Foundation: Beaverton, OR, USA, 2009; Available online: http://qgis.org (accessed on 1 March 2025).
- Florenzano, A.; Marignani, M.; Rosati, L.; Fascetti, S.; Mercuri, A.M. Are Cichorieae an indicator of open habitats and pastoralism in current and past vegetation studies? Plant Biosyst. 2015, 149, 154–165. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 10 March 2025).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Campitelli, E. ggnewscale: Multiple Fill and Colour Scales in ‘ggplot2’, R Package Version 0.5.1; 2025. Available online: https://CRAN.R-project.org/package=ggnewscale (accessed on 10 March 2025).
- Yu, G. shadowtext: Shadow Text Grob and Layer, R Package Version 0.1.4; 2024. Available online: https://CRAN.R-project.org/package=shadowtext (accessed on 10 March 2025).
- Vignola, C.; Bonetto, J.; Furlan, G.; Mazza, M.; Nicosia, C.; Russo Ermolli, E.; Sadori, L. At the origins of Pompeii: The plant landscape of the Sarno River floodplain from the first millennium BC to the AD 79 eruption. Veget. Hist. Archaeobot. 2022, 31, 171–186. [Google Scholar] [CrossRef]
- Buldrini, F.; Dallai, D.; Torri, P. Can palynology contribute to plant diversity conservation activities? The wetland plants in southern Po plain as a case study. Ann. Bot. 2013, 3, 245–254. [Google Scholar]
- Carter, V.A.; Chiverrell, R.C.; Clear, J.L.; Kuosmanen, N.; Moravcová, A.; Svoboda, M.; Svobodová-Svitavská, H.; van Leeuwen, J.F.N.; van der Knaap, W.O.; Kuneš, P. Quantitative Palynology Informing Conservation Ecology in the Bohemian/Bavarian Forests of Central Europe. Front. Plant Sci. 2018, 8, 2268. [Google Scholar] [CrossRef]
- Bazan, G.; Castrorao Barba, A. Historical Ecology, Archaeology and Biocultural Landscapes: Cross-Disciplinary Approaches to the Long Anthropocene. Sustainability 2022, 14, 5017. [Google Scholar] [CrossRef]
- Castrorao Barba, A.; Aleo Nero, C.; Battaglia, G.; Zambito, L.; Virga, L.; Messina, A.; Cangemi, M.; Bazan, G. Continuity, Resilience, and Change in Rural Settlement Patterns from the Roman to Islamic Period in the Sicani Mountains (Central-Western Sicily). Land 2024, 13, 400. [Google Scholar] [CrossRef]
- Florenzano, A.; Clò, E.; Servera-Vives, G.; Mercuri, A.M. Palynology for Sustainability: A Classical and Versatile Tool for New Challenges: Editorial. Sustainability 2025, 17, 1938. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A.; Clò, E.; Servera-Vives, G. Palynology for Sustainability: A Classical and Versatile Tool for New Challenges—Recent Progress. Quaternary 2025, 8, 18. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| BRAIN ID | Name | No. Pollen Records | No. Pollen Samples | Count | No. Taxa | AP/NAP |
|---|---|---|---|---|---|---|
| SCA34 | Stabiae—Villa San Marco | 1 | 9 | 3114 | 88 | 29/71 |
| SCA35 | Stabiae—Villa Arianna | 1 | 4 | 1314 | 37 | 30/70 |
| SCA36 | Pompeii—Civita Giuliana | 1 | 3 | 1961 | 94 | 9/91 |
| SBA1 | Altojanni | 2 | 11–12 | 4156–5760 | 118–95 | 15/85–8/92 |
| SBA4 | Difesa S. Biagio | 1 | 23 | 6901 | 34 | 6/94 |
| SBA5 | Fattoria Fabrizio | 1 | 12 | 6532 | 118 | 82/18 |
| SBA9 | Pantanello (Pizzica Pantanello) | 3 | 16–11–13 | 6155–3465–3866 | 129–122–92 | 9/91–11/89–4/96 |
| SBA12 | Torre di Satriano | 1 | 4 | 1191 | 70 | 7/93 |
| SSI6 | Piazza Armerina—Villa Romana del Casale | 1 | 45 | 16,322 | 207 | 11/89 |
| SSI7 | Philosophiana (Sofiana) | 1 | 12 | 5397 | 116 | 9/91 |
| SSI8 | Taormina—Teatro grecoromano | 1 | 9 | 3640 | 131 | 14/86 |
| SSI9 | Stromboli—San Vincenzo | 1 | 23 | 5328 | 103 | 13/87 |
| SSI32 | Morgantina—agorà | 1 | 10 | 2883 | 61 | 10/90 |
| SSI39 | Mozia—Stagnone di Marsala | 1 | 10 | 819 | 88 | 49/51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clò, E.; Mercuri, A.M.; Zappa, J.; Ricucci, C.; Braga, L.; Florenzano, A. Millennial Floristic Diversity and Land Management as Inferred from Archaeo-Palynological Research in Southern Italy. Plants 2025, 14, 1367. https://doi.org/10.3390/plants14091367
Clò E, Mercuri AM, Zappa J, Ricucci C, Braga L, Florenzano A. Millennial Floristic Diversity and Land Management as Inferred from Archaeo-Palynological Research in Southern Italy. Plants. 2025; 14(9):1367. https://doi.org/10.3390/plants14091367
Chicago/Turabian StyleClò, Eleonora, Anna Maria Mercuri, Jessica Zappa, Cristina Ricucci, Lorenzo Braga, and Assunta Florenzano. 2025. "Millennial Floristic Diversity and Land Management as Inferred from Archaeo-Palynological Research in Southern Italy" Plants 14, no. 9: 1367. https://doi.org/10.3390/plants14091367
APA StyleClò, E., Mercuri, A. M., Zappa, J., Ricucci, C., Braga, L., & Florenzano, A. (2025). Millennial Floristic Diversity and Land Management as Inferred from Archaeo-Palynological Research in Southern Italy. Plants, 14(9), 1367. https://doi.org/10.3390/plants14091367