
The Effect of Leaf Traits on the Excitation, Transmission, and Perception of Vibrational Mating Signals in the Tea Leafhopper Empoasca onukii Matsuda (Hemiptera: Cicadellidae)

Abstract
1. Introduction
2. Results
2.1. Female Responses to Signals from Other Leaves
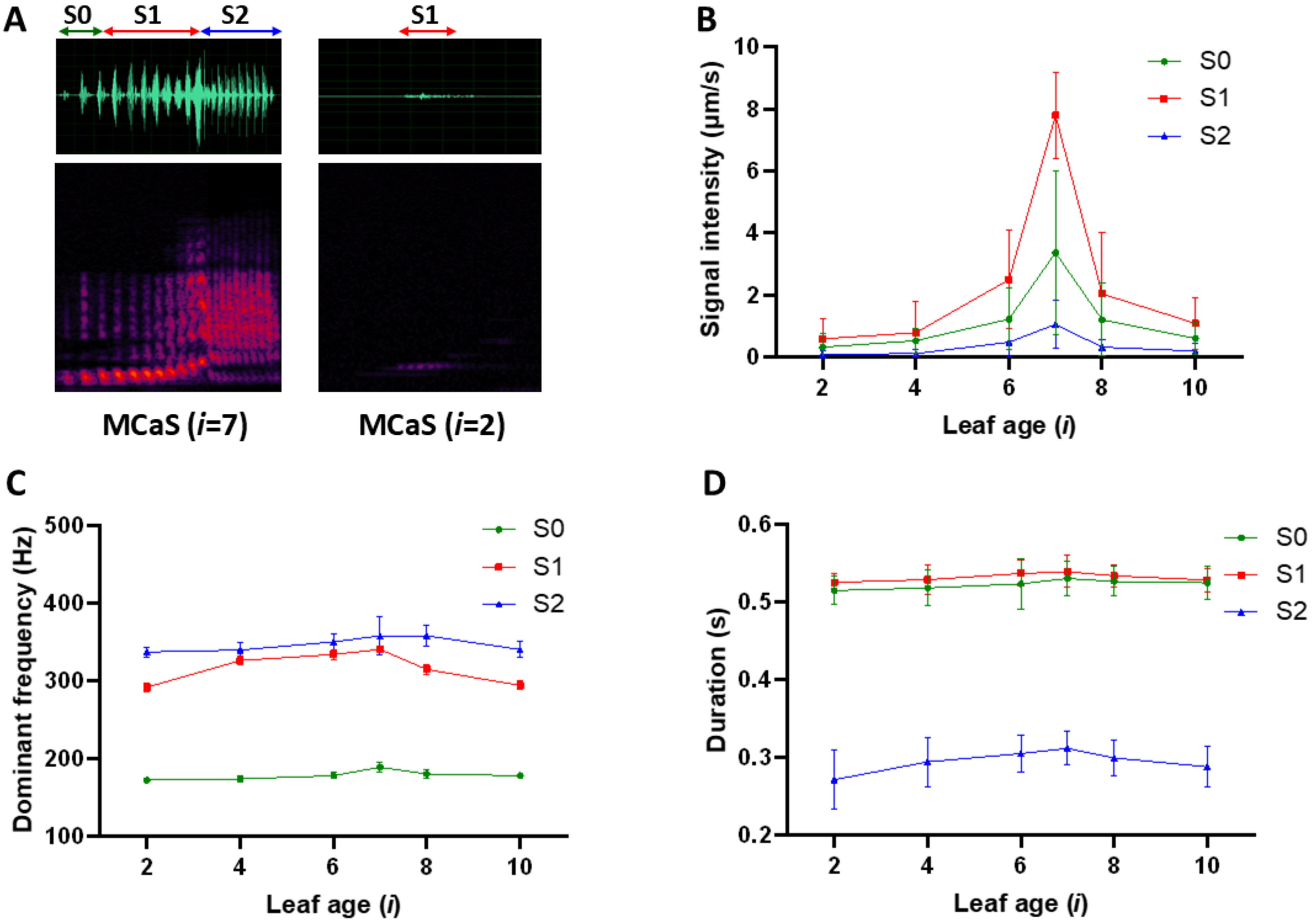
2.1.1. Variation in MCaS Parameters from Leaves of Different Ages
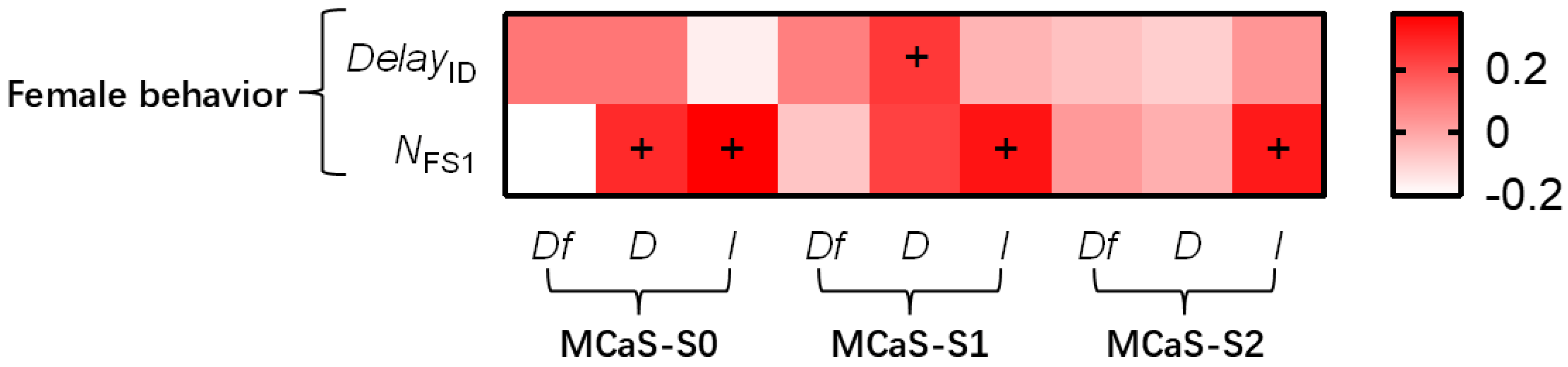
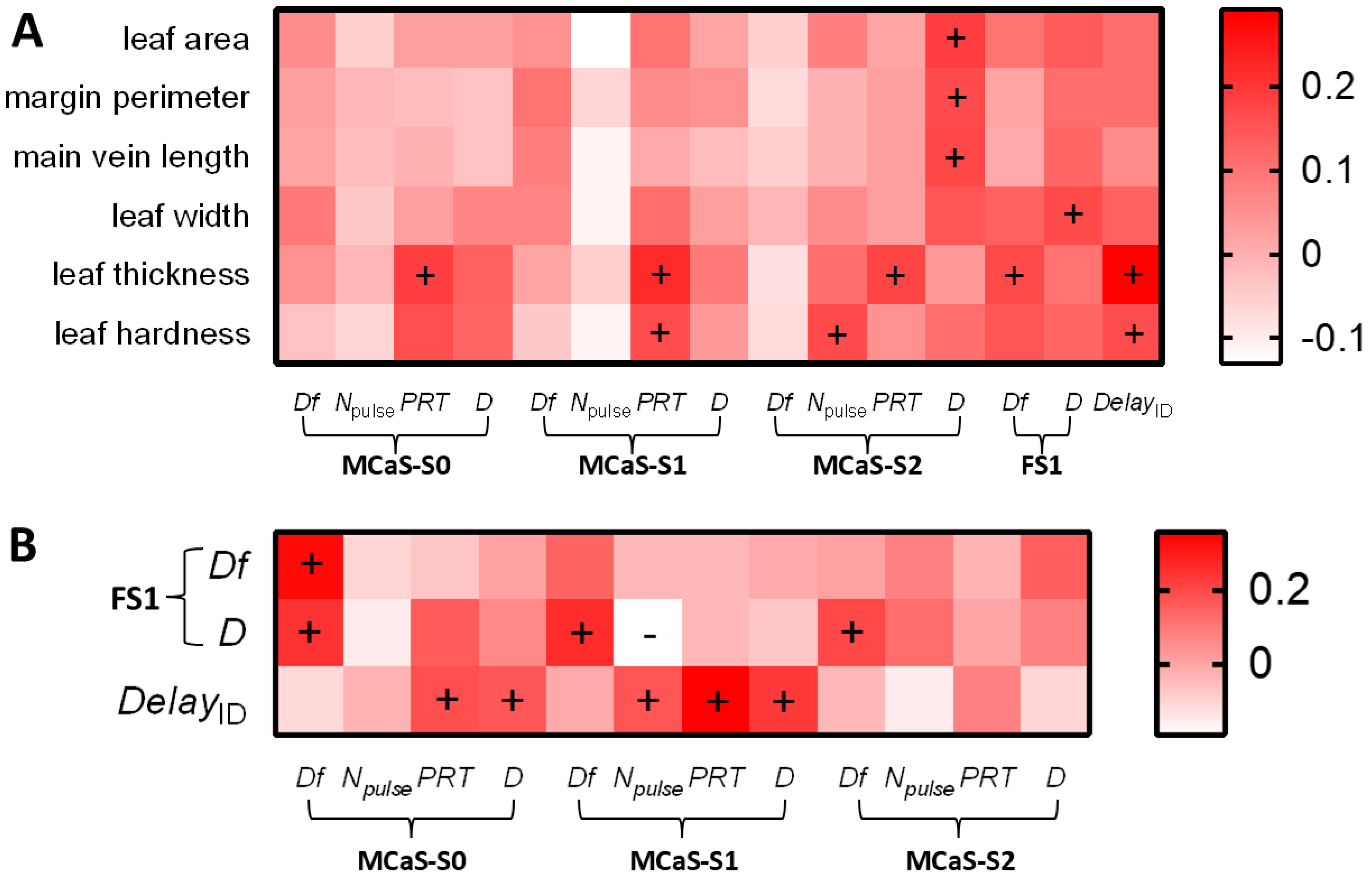
2.1.2. Effect of MCaS Parameters on Female Behavior
2.2. Female Responses to Male Signals from the Same Leaf
2.2.1. Effect of Leaf Age
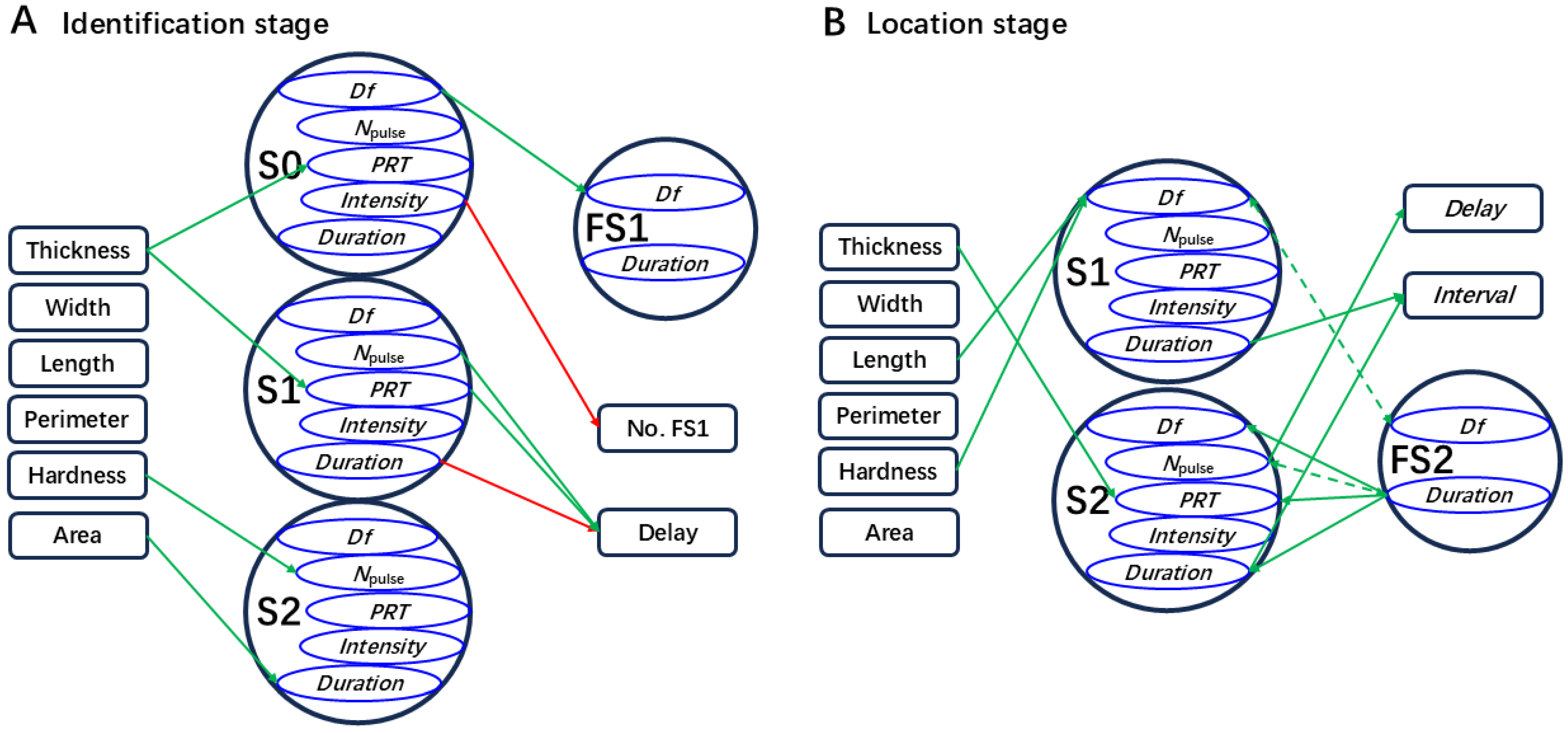
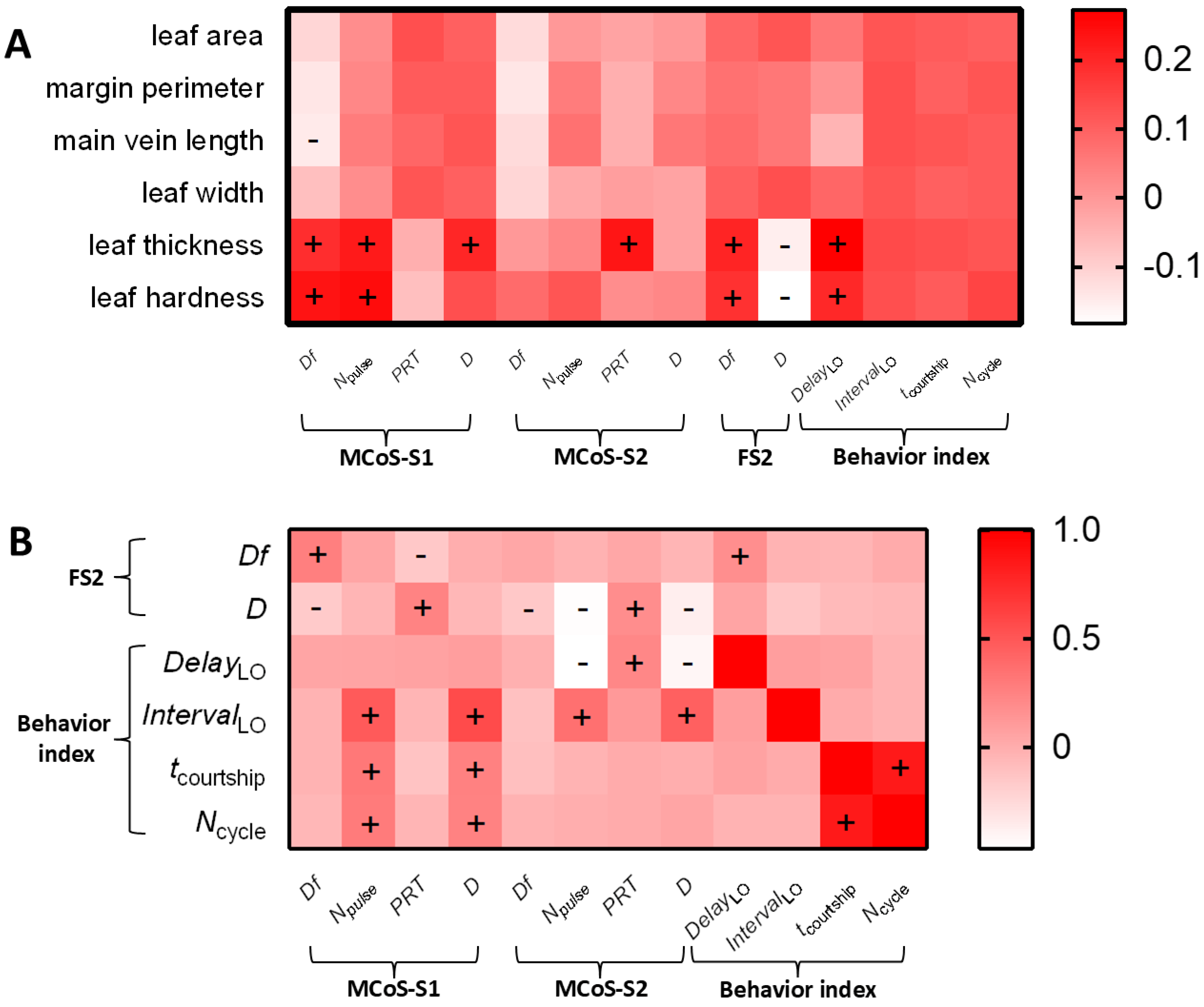
2.2.2. Effect of Leaf Traits on the Identification Stage
2.2.3. Effect of Leaf Traits on Localization Stage
3. Discussion
3.1. Effects of Leaf Traits on Mating Behavior of Leafhoppers
3.2. Potential Information Carried in Different Sections of Male Signals
4. Materials and Methods
4.1. Insect Rearing
4.2. Plants and Leaf Trait Measurements
4.3. Signal Recording and Playback
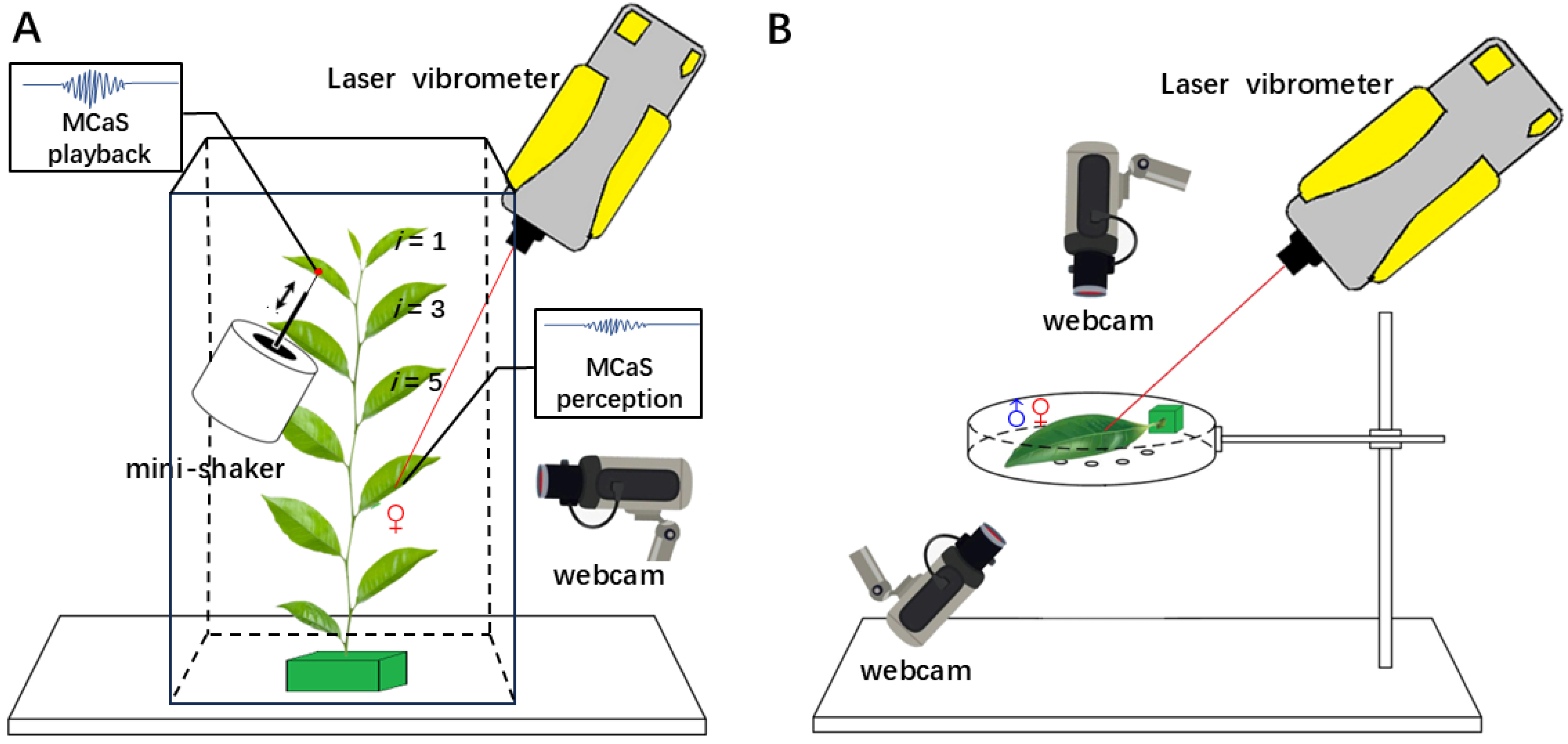
4.4. Experimental Setup
4.4.1. Female Responses to Signals from Other Leaves
4.4.2. Female Responses to Signals from the Same Leaf
4.5. Terminology and Data Analyses
4.5.1. Female Responses to Signals from Other Leaves
4.5.2. Female Responses to Signals from the Same Leaf
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cividini, S.; Giuseppe, M. Biotremology in arthropods. Learn. Behav. 2020, 48, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, V.; Anfora, G.; Cocroft, R.B.; Fatouros, N.E.; Groot, A.T.; Gross, J.; Hill, P.S.M.; Hoch, H.; Ioriatti, C.; Nieri, R.; et al. Bridging biotremology and chemical ecology: A new terminology. Trends Plant Sci. 2024, 29, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Cocroft, R.B.; Rodríguez, R.L. The behavioral ecology of insect vibrational communication. Bioscience 2005, 55, 323–334. [Google Scholar] [CrossRef]
- Cocroft, R.B.; Shugart, H.J.; Konrad, K.T.; Tibbs, K. Variation in plant substrates and its consequences for insect vibrational communication. Ethology 2006, 112, 779–789. [Google Scholar] [CrossRef]
- Joyce, A.L.; White, W.H.; Medina, R.F. Host plants impact courtship vibration transmission and mating success of a parasitoid wasp, Cotesia flavipes (Hymenoptera: Braconidae). Evol. Ecol. 2014, 28, 361–372. [Google Scholar] [CrossRef]
- Velilla, E.; Polajnar, J.; Virant-Doberlet, M.; Commandeur, D.; Simon, R.; Cornelissen, J.H.; Ellers, J.; Halfwerk, W. Variation in plant leaf traits affects transmission and detectability of herbivore vibrational cues. Ecol. Evol. 2020, 10, 12277–12289. [Google Scholar] [CrossRef]
- Mortimer, B. Biotremology: Do physical constraints limit the propagation of vibrational information? Anim. Behav. 2017, 130, 165–174. [Google Scholar] [CrossRef]
- Rosenthal, M.F.; Hebets, E.A.; Kessler, B.; McGinley, R.; Elias, D.O. The effects of microhabitat specialization on mating communication in a wolf spider. Behav. Ecol. 2019, 30, 1398–1405. [Google Scholar] [CrossRef]
- Elias, D.O.; Mason, A.C.; Hoy, R.R. The effect of substrate on the efficacy of seismic courtship signal transmission in the jumping spider Habronattus dossenus (Araneae: Salticidae). J. Expe Biol. 2004, 207, 4105–4110. [Google Scholar] [CrossRef]
- McNett, G.D.; Cocroft, R.B. Host shifts favor vibrational signal divergence in Enchenopa binotata treehoppers. Behav. Ecol. 2008, 19, 650–656. [Google Scholar] [CrossRef]
- Bian, L.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Chen, Z.M. Foliage intensity is an important cue of habitat location for Empoasca onukii. Insects 2020, 11, 426. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Zhou, X.S.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Xiu, C.L.; Chen, Z.M.; Bian, L. Mating and post-copulation behavior in the tea leafhopper, Empoasca onukii (Hemiptera: Cicadellidae). Front. Plant Sci. 2023, 14, 1273718. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, L.K.; Bhuyan, M.; Hazarika, B.N. Insect pests of tea and their management. Annu. Rev. Entomol. 2009, 54, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Virant-Doberlet, M.; Stritih-Peljhan, N.; Žunič-Kosi, A.; Polajnar, J. Functional diversity of vibrational signaling systems in insects. Annu. Rev. Entomol. 2023, 68, 191–210. [Google Scholar] [CrossRef]
- Zhang, H.; Bian, L.; Cai, X.M.; Yao, Q.; Fu, N.X.; Shan, Y.; Chen, Z.M. Vibrational signals are species-specific and sex-specific for sexual communication in the tea leafhopper, Empoasca onukii. Entomol. Exp. Appl. 2023, 171, 277–286. [Google Scholar] [CrossRef]
- Mazzoni, V.; Eriksson, A.; Anfora, G.; Lucchi, A.; Virant-Doberlet, M. Active space and the role of amplitude in plant-borne vibrational communication. In Studying Vibrational Communication. Animal Signals and Communication; Cocroft, R., Gogala, M., Hill, P., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 3, pp. 125–145. [Google Scholar] [CrossRef]
- Kuhelj, A.; De Groot, M.; Pajk, F.; Simčič, T.; Virant-Doberlet, M. Energetic cost of vibrational signalling in a leafhopper. Behav. Ecol. Sociobiol. 2015, 69, 815–828. [Google Scholar]
- Virant-doberlet, M.; King, R.A.; Polajnar, J.; Symondson, W.O. Molecular diagnostics reveal spiders that exploit prey vibrational signals used in sexual communication. Mole Ecol. 2011, 20, 2204–2216. [Google Scholar]
- Huang, Y.H.; Zhang, J.W.; Zhang, Y.L.; Yang, Y.; Wang, Y.J. Anatomical characteristics of leaf structure of tea plant resistant to leafhopper (Empoasca vitis Gothe). J. Tea Sci. 1998, 18, 35–38. [Google Scholar]
- De Langre, E. Plant vibrations at all scales: A review. J. Exp. Bot. 2019, 70, 3521–3531. [Google Scholar] [CrossRef]
- Polajnar, J.; Kavčič, A.; Kosi, A.; Čokl, A. Palomena prasina (Hemiptera: Pentatomidae) vibratory signals and their tuning with plant substrates. Open Life Sci. 2013, 8, 670–680. [Google Scholar]
- Čokl, A.; Zorović, M.; Millar, J.G. Vibrational communication along plants by the stink bugs Nezara viridula and Murgantia histrionica. Behav. Process. 2007, 75, 40–54. [Google Scholar]
- Kuhelj, A.; de Groot, M.; Blejec, A.; Virant-Doberlet, M. Sender-receiver dynamics in leafhopper vibrational duetting. Anim. Behav. 2016, 114, 139–146. [Google Scholar]
- Guilford, T.; Dawkins, M.S. Receiver psychology and the evolution of animal signals. Anim. Behav. 1991, 42, 1–14. [Google Scholar]
- Pollack, G. Who, what, where? Recognition and localization of acoustic signals by insects. Curr. Opin. Neurobiol. 2000, 10, 763–767. [Google Scholar] [PubMed]
- Čokl, A.; Dias, A.M.; Moraes, M.C.B.; Borges, M.; Laumann, R.A. Rivalry between stink bug females in a vibrational communication network. J. Insect Behav. 2017, 30, 741–758. [Google Scholar]
- Čokl, A.; Blassioli-Moraes, M.C.; Laumann, R.A.; Žunič, A.; Borges, M. Stinkbugs: Multisensory Communication with Chemical and Vibratory Signals Transmitted Through Different Media. In Biotremology: Studying Vibrational Behavior. Animal Signals and Communication; Hill, P., Lakes-Harlan, R., Mazzoni, V., Narins, P., Virant-Doberlet, M., Wessel, A., Eds.; Springer: Cham, Switzerland, 2019; Volume 6, pp. 91–122. [Google Scholar]
- Cocroft, R.B.; Hamel, J.; Su, Q.; Gibson, J. Vibrational Playback Experiments: Challenges and Solutions. In Studying Vibrational Communication. Animal Signals and Communication; Cocroft, R., Gogala, M., Hill, P., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 3, pp. 249–274. [Google Scholar]
- Nieri, R.; Anfora, G.; Mazzoni, V.; Stacconi, R. Semiochemicals, semiophysicals and their integration for the development of innovative multi-modal systems for agricultural pests’ monitoring and control. J. Mediterr. Ecol. 2021, 19, 42. [Google Scholar]
- Chen, Z.M.; Luo, Z.X. Management of insect pests on tea plantations: Safety, sustainability, and efficiency. Annu. Rev. Entomol. 2025, 70, 359–377. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Traits of Host Plants |
|---|
| Leaf area (cm2), leaf thickness (mm), leaf width (mm), main vein length (mm), leaf margin perimeter (mm), and leaf hardness (g). Leaf age (i), leaves from the top to the bottom of a tea branch were numbered. |
| Signal parameters |
| Male signal: male calling signal (MCaS, with three sections: S0, S1, and S2) and male courtship signal (MCoS, with two sections: S1, and S2). Female signal: female signal responses to the MCaS (FS1) or MCoS (FS2). Parameter: dominant frequency (Df), PRT, Npulse, duration (D), and intensity (I). PRT: pulse repetition time in each section of the MCaS or MCoS. Npulse: the number of pulses in each section of the MCaS or MCoS. |
| Behavioral indexes |
| Female behavior: DelayID, DelayLO, and NFS1. Male behavior: IntervalLO, tcourtship, and Ncycle. DelayID: the time from the end of the MCaS to the beginning of the subsequent FS1. DelayLO: the time from the end of the MCoS to the beginning of the subsequent FS2. NFS1: the total number of FS1 in a signal sample in Section 4.4.1. IntervalLO: the interval from the end of the MCoS to the beginning of the subsequent MCoS. tcourtship: the whole time spent in courtship from the first MCaS to copulation in a signal sample in Section 4.4.2. Ncycle: the number of localization cycles in a signal sample in Section 4.4.2. Some males needed to re-enter the identification stage several times during the localization process. We defined each time that the male entered the localization stage in the localization process as a localization cycle. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, Y.; Yao, Q.; Jia, Q.; Lu, J.; Cai, X.; Chen, Z.; Bian, L. The Effect of Leaf Traits on the Excitation, Transmission, and Perception of Vibrational Mating Signals in the Tea Leafhopper Empoasca onukii Matsuda (Hemiptera: Cicadellidae). Plants 2025, 14, 1147. https://doi.org/10.3390/plants14071147
Shan Y, Yao Q, Jia Q, Lu J, Cai X, Chen Z, Bian L. The Effect of Leaf Traits on the Excitation, Transmission, and Perception of Vibrational Mating Signals in the Tea Leafhopper Empoasca onukii Matsuda (Hemiptera: Cicadellidae). Plants. 2025; 14(7):1147. https://doi.org/10.3390/plants14071147
Chicago/Turabian StyleShan, Yao, Qiuyi Yao, Qisheng Jia, Jiping Lu, Xiaoming Cai, Zongmao Chen, and Lei Bian. 2025. "The Effect of Leaf Traits on the Excitation, Transmission, and Perception of Vibrational Mating Signals in the Tea Leafhopper Empoasca onukii Matsuda (Hemiptera: Cicadellidae)" Plants 14, no. 7: 1147. https://doi.org/10.3390/plants14071147
APA StyleShan, Y., Yao, Q., Jia, Q., Lu, J., Cai, X., Chen, Z., & Bian, L. (2025). The Effect of Leaf Traits on the Excitation, Transmission, and Perception of Vibrational Mating Signals in the Tea Leafhopper Empoasca onukii Matsuda (Hemiptera: Cicadellidae). Plants, 14(7), 1147. https://doi.org/10.3390/plants14071147