
Nutrient Additions Regulate Height Growth Rate but Not Biomass Growth Rate of Alpine Plants Through the Contrasting Effect of Total and Available Nitrogen
 , ,
, ,
Abstract
1. Introduction
2. Results
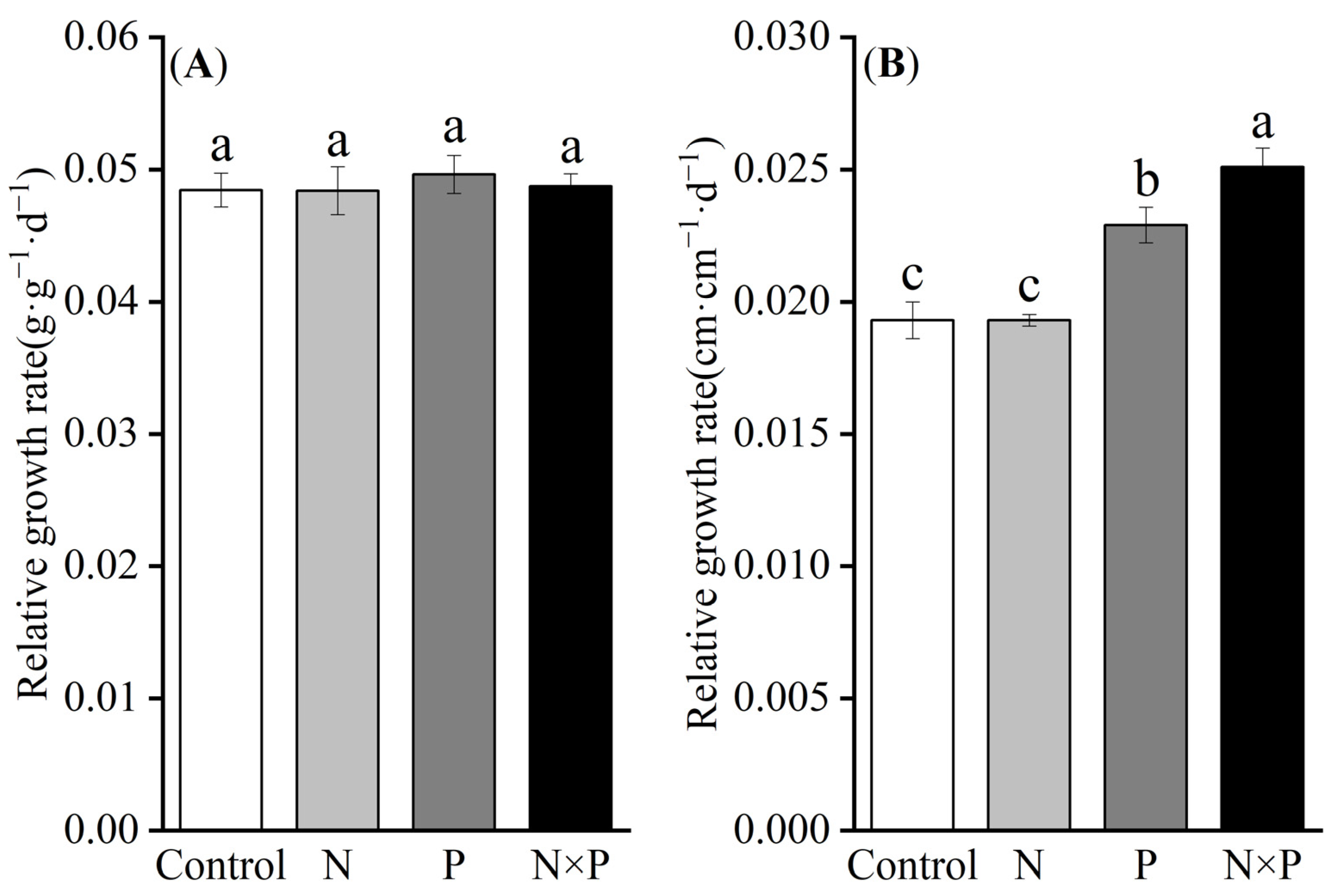
2.1. Response of the Growth Rate of Alpine Plants to Nutrient Additions
2.2. Response of the Growth Rate of Different Species to Nutrient Additions
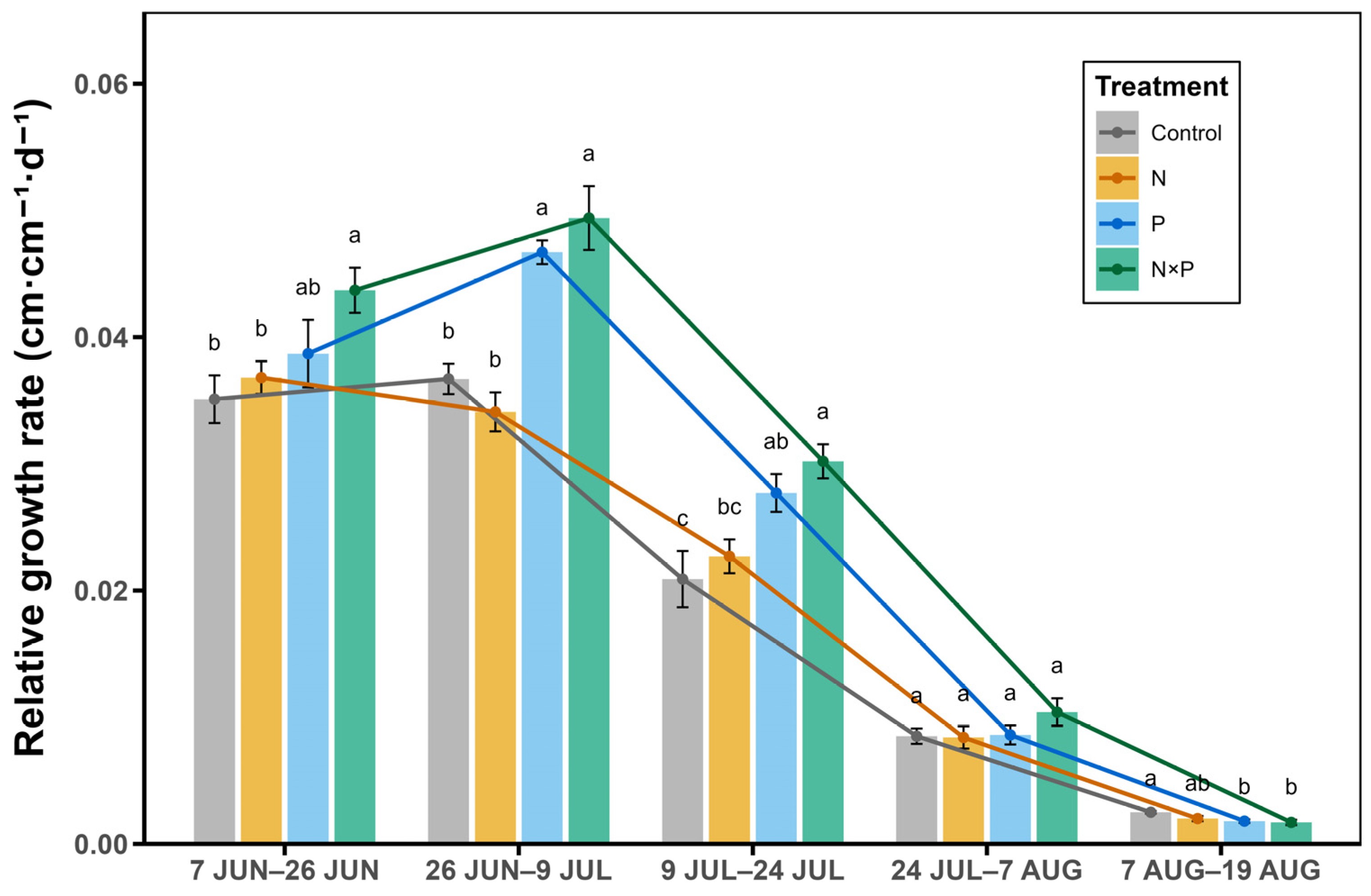
2.3. Seasonal Dynamic of the Growth Rate of Alpine Plants Under Different Nutrient Additions
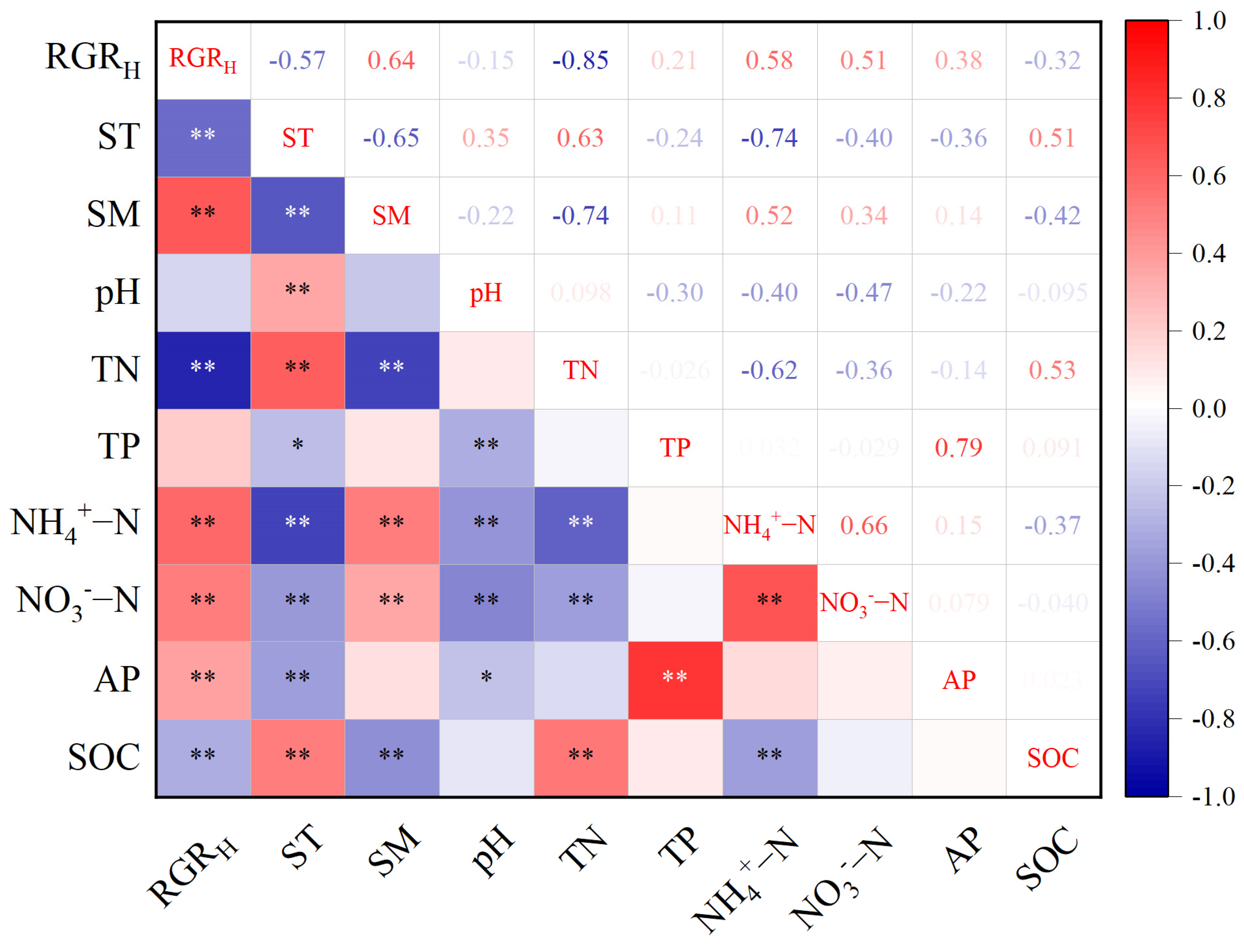
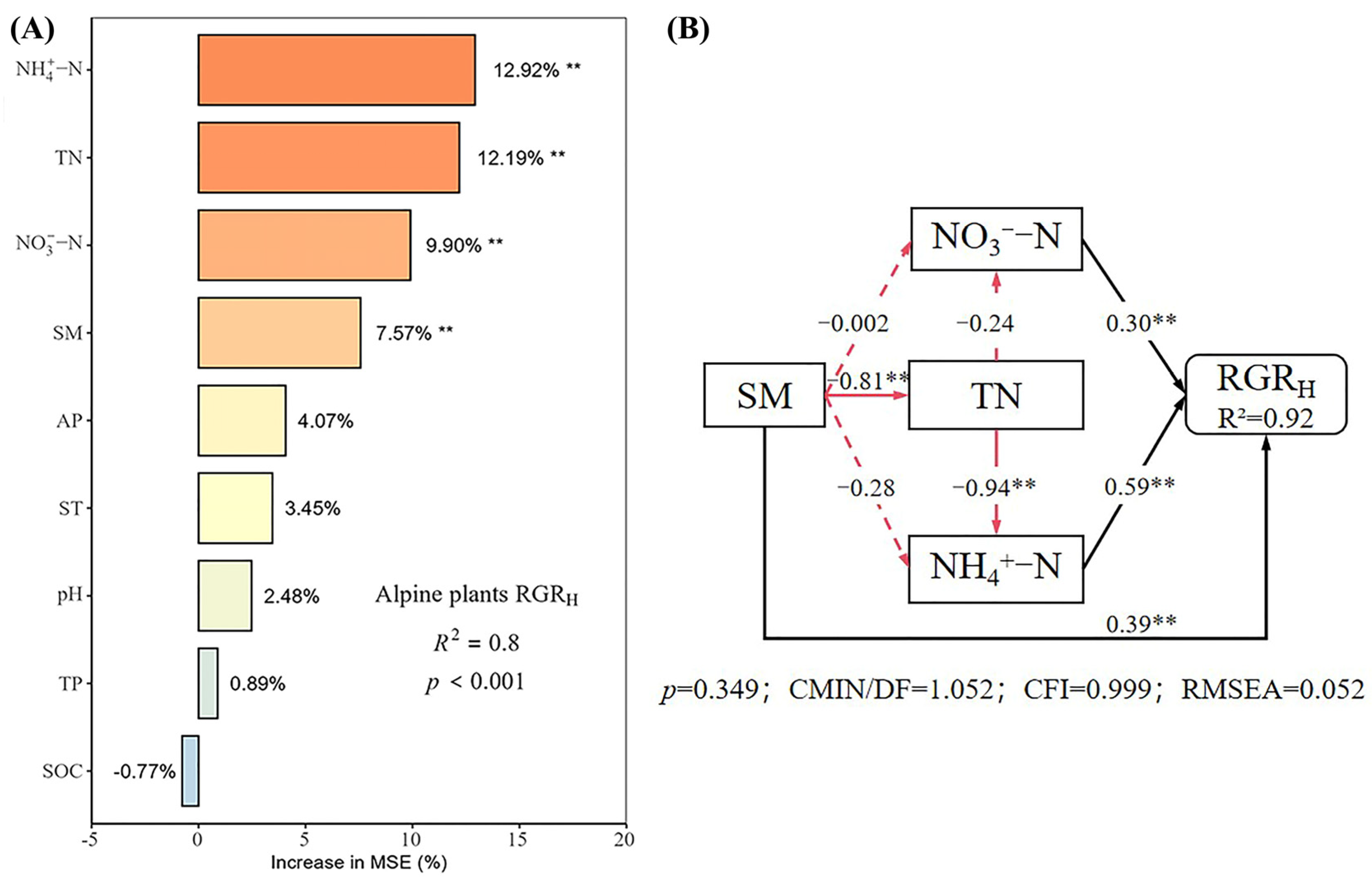
2.4. Relationship Between Growth Rate and Soil Physical and Chemical Properties
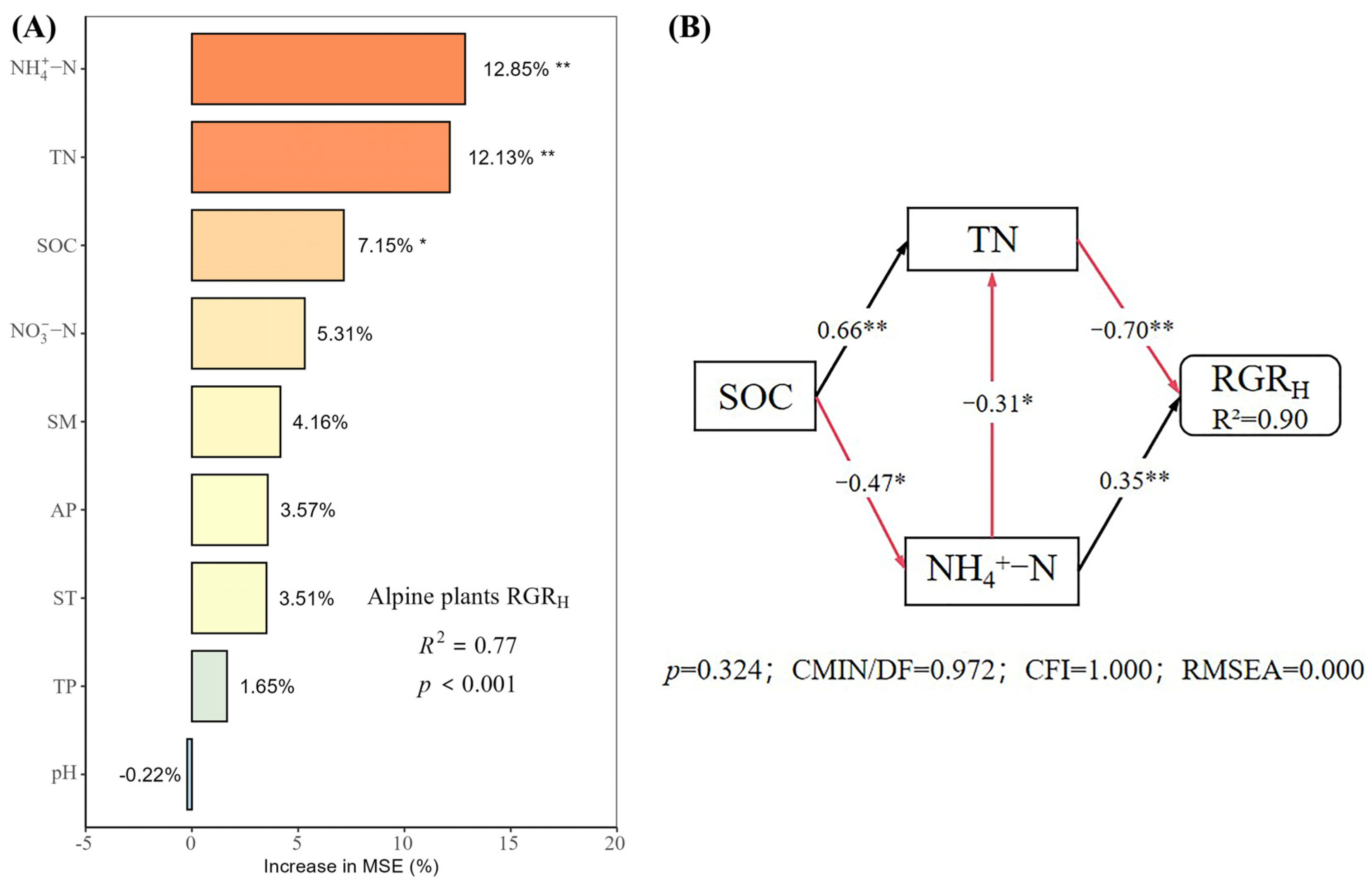
2.5. Direct and Indirect Effects of Soil Physical and Chemical Properties on Plant Growth
3. Discussion
4. Materials and Methods
4.1. Study Area and Experimental Design
4.2. Sample Collection and Measurement
4.3. Determination of Soil Physical and Chemical Properties
4.4. Calculation of the Plant Growth Rate
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hilty, J.; Muller, B.; Pantin, F.; Leuzinger, S. Plant growth: The What, the How, and the Why. New Phytol. 2021, 232, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Saussure, N.T.D. Recherches Chimiques sur la Vegetation, 5th ed.; Garrison-Morton: Nyon, Switzerland, 1804. [Google Scholar]
- Sprengel, C. Von den Substanzen der Ackerkrume und des Untergrundes (About the substances in the plow layer and the subsoil). J. Tech. Okon. Chem. 1828, 2, 397–421. [Google Scholar]
- Liebig, J.V. Die Organische Chemie in ihrer Anwendung auf Agricultur und Physiologie; Friedrich Vieweg und Sohn: Braunschweig, Germany, 1841. [Google Scholar]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Jelliffe, J.; Fuglie, K.; Morgan, S. Productivity-led Pathways to Sustainable Agricultural Growth: Six Decades of Progress. EuroChoices 2024, 23, 4–13. [Google Scholar] [CrossRef]
- Hancock, N.I. Relative Growth Rate of the Main Stem of the Cotton Plant and Its Relationship to Yield. Agron. J. 1941, 33, 590–602. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Connell, J.H.; Slatyer, R.O. Mechanisms of Succession in Natural Communities and Their Role in Community Stability and Organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Vitousek, P.M.; Chapin, M.C. Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer: New York, NY, USA, 2011. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Cham, Switzerland, 2003. [Google Scholar]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient co-limitation of primary producer communities. Ecol. Lett. 2011, 14, 852–862. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.Å.M.; Billberger, M.F.K. Nutrient limitation on terrestrial plant growth–modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.P.; Yang, Y.H.; Yu, M.; Wang, C.; Yan, J. Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type. Plant Soil 2019, 440, 523–537. [Google Scholar] [CrossRef]
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlström, A.; Van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Zhu, J.X.; Wang, Q.F.; He, N.P.; Smith, M.D.; Elser, J.J.; Du, J.Q.; Yu, G.; Yu, Q. Imbalanced atmospheric nitrogen and phosphorus depositions in China: Implications for nutrient limitation. J. Geophys. Res. Biogeosci. 2016, 121, 1605–1616. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, B.; Cao, J.; Liu, J.; Tian, S.; Du, E. Enhanced atmospheric phosphorus deposition in Asia and Europe in the past two decades. Atmos. Ocean. Sci. Lett. 2021, 14, 100051. [Google Scholar] [CrossRef]
- Li, S.; Dong, S.K.; Fu, Y.S.; Zhou, B.R.; Liu, S.L.; Shen, H.; Xu, Y.D.; Gao, X.X.; Xiao, J.N.; Wu, S.N.; et al. Air or soil temperature matters the responses of alpine plants in biomass accumulation to climate warming. Sci. Total Environ. 2022, 844, 157141. [Google Scholar] [CrossRef]
- Pugnaire, F.; Losapio, G.; Schöb, C. Species interactions involving cushion plants in high-elevation environments under a changing climate. Ecosistemas 2021, 30, 2186. [Google Scholar] [CrossRef]
- Alrhmoun, M.; Romano, A.; Sulaiman, N.; Pieroni, A. Old Plants for New Food Products? The Diachronic Human Ecology of Wild Herbs in the Western Alps. Plants 2025, 14, 122. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X. Response of Alpine Plants to Nitrogen Addition on the Tibetan Plateau: A Meta-analysis. J. Plant Growth Regul. 2016, 35, 974–979. [Google Scholar] [CrossRef]
- Shen, H.; Dong, S.K.; DiTommaso, A.; Xiao, J.N.; Lu, W.; Zhi, Y.L. Nitrogen Deposition Shifts Grassland Communities Through Directly Increasing Dominance of Graminoids: A 3-Year Case Study from the Qinghai-Tibetan Plateau. Front. Plant Sci. 2022, 13, 811970. [Google Scholar] [CrossRef]
- Wang, L.; Shi, J.J.; Shang, Z.H.; Shi, H.L.; Chen, W.Y.; Geng, G.C.L.; Geng, Q.B.M. Effect of phosphatic fertilizer addition on plant community characteristics in an alpine grassland around Qinghai Lake. Pratacultural Sci. 2019, 36, 1224–1230. [Google Scholar]
- Du, L.; Tang, L.S.; Zheng, X.J.; Li, Y. A global analysis of plant nutrient limitation affected by atmospheric nitrogen and phosphorous deposition. Front. Plant Sci. 2024, 15, 1473493. [Google Scholar] [CrossRef]
- Hunt, R. Plant Growth Curves: The Functional Approach to Plant Growth Analysis, 1st ed.; Edward Arnold: London, UK, 1982. [Google Scholar]
- Falster, D.S.; Westoby, M. Plant height and evolutionary games. Trends Ecol. Evol. 2003, 18, 337–343. [Google Scholar] [CrossRef]
- Ye, Q.D.; Ren, F.; Li, Y.H.; Zhang, Z.Y.; Fan, N.; Ye, J.; Li, X.L.; Li, L.P. Effects of Different Nutrient Additions on Functional Traits of Alpin Plant. Acta Agrestia Sin. 2022, 30, 2737–2744. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, M.J.; Guo, T.T.; Cui, J.; Li, L.P.; Liang, D.F.; Li, Y.H.; Zhang, Z.H.; Zhu, X.X.; Ren, F. Effects of Multi-gradient Nitrogen and Phosphorus Additions on Biomass and Nitrogen and Phosphorus Content of Alpine Meadow Plant Community. Acta Agrestia Sin. 2023, 31, 751–759. [Google Scholar] [CrossRef]
- Gao, Y.H.; Cooper, D.J.; Zeng, X.Y. Nitrogen, not phosphorus, enrichment controls biomass production in alpine wetlands on the Tibetan Plateau, China. Ecol. Eng. 2018, 116, 31–34. [Google Scholar] [CrossRef]
- Fashaho, A.; Musandu, A.O.; Lelei, J.J.; Mwonga, S.M.; Ndegwa, G.M. Effects of Nitrogen and Phosphorus Fertilizer Rates on Maize (Zea mays L.) Growth and Yields in Terraced Lands of Medium and High Altitude Regions of Rwanda. Agric. Sci. Dig. 2020, 40, 242–248. [Google Scholar] [CrossRef]
- Zhan, S.X.; Zheng, S.X.; Wang, Y.; Bai, Y.F. Response and correlation of above-and below-ground functional traits of Leymus chinensis to nitrogen and phosphorus additions. Chin. J. Plant Ecol. 2016, 40, 36–47. [Google Scholar] [CrossRef]
- Zong, N.; Shi, P.L.; Zhao, G.S.; Zheng, L.L.; Niu, B.; Zhou, T.C.; Hou, G. Variations of nitrogen and phosphorus limitation along the environmental gradient in alpine grasslands on the Northern Xizang Plateau. Chin. J. Plant Ecol. 2021, 45, 444. [Google Scholar] [CrossRef]
- Dong, K.; Li, W.J.; Tang, Y.L.; Ma, S.H.; Jiang, M.L. Co-limitation of N and P is more prevalent in the Qinghai–Tibetan Plateau grasslands. Front. Plant Sci. 2023, 14, 1140462. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.Y.; Cao, G.M.; Ma, Z.Y.; Li, Y.K.; Zhang, F.W.; Zhao, X.; Zhao, X.Q.; Jiang, L.; Sanders, N.J.; et al. Alpine grassland plants grow earlier and faster but biomass remains unchanged over 35 years of climate change. Ecol. Lett. 2020, 23, 701–710. [Google Scholar] [CrossRef]
- Dickson, T.L.; Mittelbach, G.G.; Reynolds, H.L.; Gross, K.L. Height and clonality traits determine plant community responses to fertilization. Ecology 2014, 95, 2443–2452. [Google Scholar] [CrossRef]
- Lin, W.S.; De, K.J.; Xiang, X.M.; Feng, T.X.; Li, F.; Wei, X.J. Characterization of vegetation communities and soil nutrient changes in degraded alpine meadows on the Qinghai–Tibetan Plateau. Grassl. Turf. 2024, 1–14. Available online: https://kns.cnki.net/kcms/detail/62.1156.S.20240910.1046.002.html (accessed on 30 October 2024).
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Yang, Y.J.; Li, B.W.; Li, W.J.; Wang, G.; Knops, J.M.H. Effects of Nitrogen and Phosphorus Fertilization on Soil Carbon Fractions in Alpine Meadows on the Qinghai–Tibetan Plateau. PLoS ONE 2014, 9, e103266. [Google Scholar] [CrossRef]
- Luo, R.Y.; Fan, J.L.; Wang, W.J.; Luo, J.F.; Kuzyakov, Y.; He, J.S.; Chu, H.Y.; Ding, W.X. Nitrogen and phosphorus enrichment accelerates soil organic carbon loss in alpine grassland on the Qinghai–Tibetan Plateau. Sci. Total Environ. 2019, 650, 303–312. [Google Scholar] [CrossRef]
- Winkler, D.E.; Chapin, K.J.; Kueppers, L.M. Soil moisture mediates alpine life form and community productivity responses to warming. Ecology 2016, 97, 1553–1563. [Google Scholar] [CrossRef]
- Ma, T.; Yang, Z.; Shi, B.; Gao, W.; Li, Y.; Zhu, J.; He, J.S. Phosphorus supply suppressed microbial necromass but stimulated plant lignin phenols accumulation in soils of alpine grassland on the Tibetan Plateau. Geoderma 2023, 431, 116376. [Google Scholar] [CrossRef]
- Liu, J.; Wu, N.; Wang, H.; Sun, J.; Peng, B.; Jiang, P.; Bai, E. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 2016, 97, 1796–1806. [Google Scholar] [CrossRef]
- Gao, W.J.; Ma, T.; Shi, B.W.; Yang, Z.Y.; Li, Y.F.; Zhu, J.X.; He, J.S. Effects of nitrogen and phosphorus addition on the mineralization potential of soil organic carbon and the corresponding regulations in the Tibetan alpine grassland. Appl. Soil Ecol. 2024, 196, 105314. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Li, X.Y. Soil moisture and temperature dynamics in typical alpine ecosystems: A continuous multi-depth measurements-based analysis from the Qinghai-Tibet Plateau, China. Hydrol. Res. 2018, 49, 194–209. [Google Scholar] [CrossRef]
- Brye, K.R.; Gbur, E.E. Regional Differences in Soil Carbon and Nitrogen Storage as Affected by Land Use and Soil Moisture Regime. Soil Sci. 2010, 175, 339–348. [Google Scholar] [CrossRef]
- Han, L.; Ganjurjav, H.; Hu, G.Z.; Wu, J.S.; Yan, Y.L.; Danjiu, L.; He, S.C.; Xie, W.D.; Yan, J.; Gao, Q.Z. Nitrogen Addition Affects Ecosystem Carbon Exchange by Regulating Plant Community Assembly and Altering Soil Properties in an Alpine Meadow on the Qinghai–Tibetan Plateau. Front. Plant Sci. 2022, 13, 900722. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.H.; Cao, Z.A.; Li, X.G.; Scholten, T.; Kühn, P.; Wang, L.; Yu, R.P.; He, J.S. Soil phosphorus availability mediates the effects of nitrogen addition on community-and species-level phosphorus-acquisition strategies in alpine grasslands. Sci. Total Environ. 2024, 906, 167630. [Google Scholar] [CrossRef] [PubMed]
- Rosolem, C.A.; Tavares, C.A. Phosphorus deficiency symptoms in soybean. Rev. Bras. Cienc. Solo 2006, 30, 385–389. [Google Scholar] [CrossRef]
- Qiao, Y.; Tang, C.; Han, X.; Miao, S. Phosphorus Deficiency Delays the Onset of Nodule Function in Soybean. J. Plant Nutr. 2007, 30, 1341–1353. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X.L. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Feng, J.G.; Zhang, Q.F.; Yuan, X.; Zhu, B. Effects of nitrogen and phosphorus addition on soil organic carbon: Review and prospects. Chin. J. Plant Ecol. 2020, 46, 855–870. [Google Scholar] [CrossRef]
- Ma, Z.Y.; Liu, H.Y.; Mi, Z.R.; Zhang, Z.H.; Wang, Y.H.; Xu, W.; Jiang, L.; He, J.S. Climate warming reduces the temporal stability of plant community biomass production. Nat. Commun. 2017, 8, 15378. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Control | N | P | N × P |
|---|---|---|---|---|
| P. versicolor | 0.022 ± 0.001 c | 0.025 ± 0.001 bc | 0.027 ± 0.001 ab | 0.031 ± 0.001 a |
| A. obtusiloba | 0.028 ± 0.000 a | 0.021 ± 0.002 b | 0.027 ± 0.001 a | 0.027 ± 0.001 a |
| R. membranaceus | 0.017 ± 0.001 bc | 0.016 ± 0.002 c | 0.022 ± 0.002 a | 0.022 ± 0.001 ab |
| M. chinensis | 0.022 ± 0.003 a | 0.023 ± 0.002 a | 0.027 ± 0.001 a | 0.025 ± 0.001 a |
| T. lanceolata | 0.017 ± 0.001 b | 0.016 ± 0.001 b | 0.022 ± 0.000 a | 0.024 ± 0.002 a |
| E. nutans | 0.023 ± 0.002 b | 0.022 ± 0.002 b | 0.027 ± 0.001 a | 0.031 ± 0.000 a |
| P. saundersiana | 0.023 ± 0.001 a | 0.021 ± 0.001 a | 0.022 ± 0.002 a | 0.026 ± 0.002 a |
| M. ruthenica | 0.023 ± 0.001 b | 0.024 ± 0.000 b | 0.031 ± 0.001 a | 0.032 ± 0.001 a |
| S. nigrescens | 0.014 ± 0.005 b | 0.018 ± 0.001 ab | 0.025 ± 0.001 a | 0.026 ± 0.002 a |
| G. straminea | 0.019 ± 0.000 a | 0.019 ± 0.001 a | 0.016 ± 0.004 a | 0.019 ± 0.001 a |
| A. diplostephioides | 0.015 ± 0.001 b | 0.018 ± 0.001 b | 0.018 ± 0.002 b | 0.024 ± 0.001 a |
| G. boreale | 0.016 ± 0.001 b | 0.016 ± 0.001 b | 0.020 ± 0.002 b | 0.026 ± 0.002 a |
| S. pulchra | 0.013 ± 0.000 bc | 0.011 ± 0.001 c | 0.016 ± 0.001 ab | 0.018 ± 0.002 a |
| O. kansuensis | 0.021 ± 0.001 a | 0.021 ± 0.001 a | 0.023 ± 0.001 a | 0.024 ± 0.001 a |
| Factors | df | F | p |
|---|---|---|---|
| N addition | 1 | 6.586 | 0.010 * |
| P addition | 1 | 117.313 | 0.000 ** |
| N addition × P addition | 1 | 5.939 | 0.015 * |
| N addition × P addition × Species | 52 | 6.443 | 0.000 ** |
| N addition × P addition × Time | 16 | 282.104 | 0.000 ** |
| N addition × P addition × Time × Species | 208 | 3.822 | 0.000 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, R.; Wang, S.; Ma, J.; Wang, N.; Wang, X.; Ren, F.; Li, H.; Liang, D.; Hu, J.; Li, X.; et al. Nutrient Additions Regulate Height Growth Rate but Not Biomass Growth Rate of Alpine Plants Through the Contrasting Effect of Total and Available Nitrogen. Plants 2025, 14, 1143. https://doi.org/10.3390/plants14071143
Feng R, Wang S, Ma J, Wang N, Wang X, Ren F, Li H, Liang D, Hu J, Li X, et al. Nutrient Additions Regulate Height Growth Rate but Not Biomass Growth Rate of Alpine Plants Through the Contrasting Effect of Total and Available Nitrogen. Plants. 2025; 14(7):1143. https://doi.org/10.3390/plants14071143
Chicago/Turabian StyleFeng, Runfang, Shu Wang, Jikui Ma, Nannan Wang, Xiaoli Wang, Fei Ren, Honglin Li, Defei Liang, Jing Hu, Xilai Li, and et al. 2025. "Nutrient Additions Regulate Height Growth Rate but Not Biomass Growth Rate of Alpine Plants Through the Contrasting Effect of Total and Available Nitrogen" Plants 14, no. 7: 1143. https://doi.org/10.3390/plants14071143
APA StyleFeng, R., Wang, S., Ma, J., Wang, N., Wang, X., Ren, F., Li, H., Liang, D., Hu, J., Li, X., & Li, L. (2025). Nutrient Additions Regulate Height Growth Rate but Not Biomass Growth Rate of Alpine Plants Through the Contrasting Effect of Total and Available Nitrogen. Plants, 14(7), 1143. https://doi.org/10.3390/plants14071143