
Transcriptome Analysis Reveals Distinct Differences in Organic Acid Metabolism Between the Pericarp and the Pulp of Cerasus humilis During Fruit Maturation

Abstract
1. Introduction
2. Results
2.1. Analysis of Organic Acid Components and Content
2.2. Changes in the Activity of Enzymes Related to Acid Metabolism
2.3. Screening of Genes Involved in Organic Acid Metabolism Differences Between the Pericarp and the Pulp of Cerasus humilis During Development
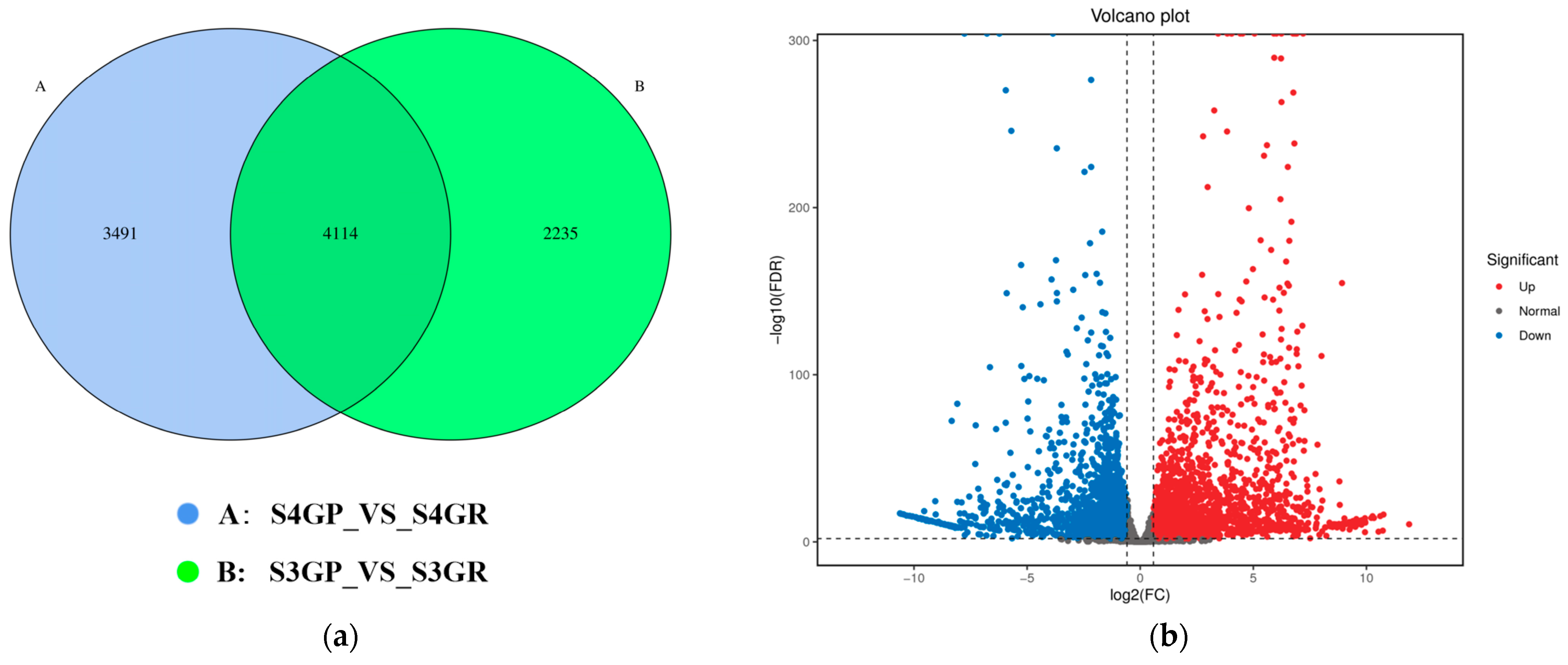
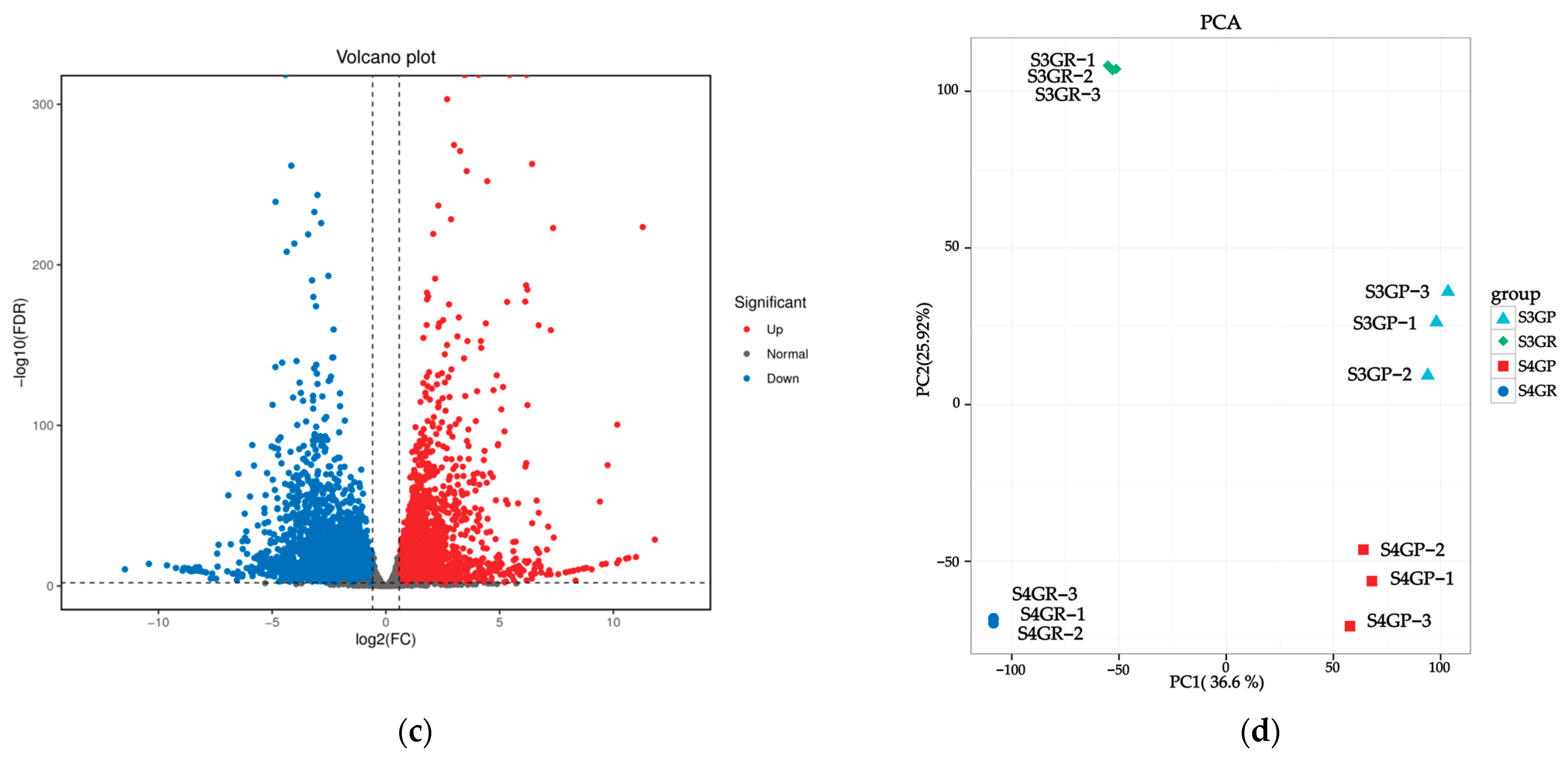
2.3.1. Differential Gene Analysis of the Transcriptome Between the Pericarp and the Pulp
2.3.2. Differential Gene Enrichment Analysis
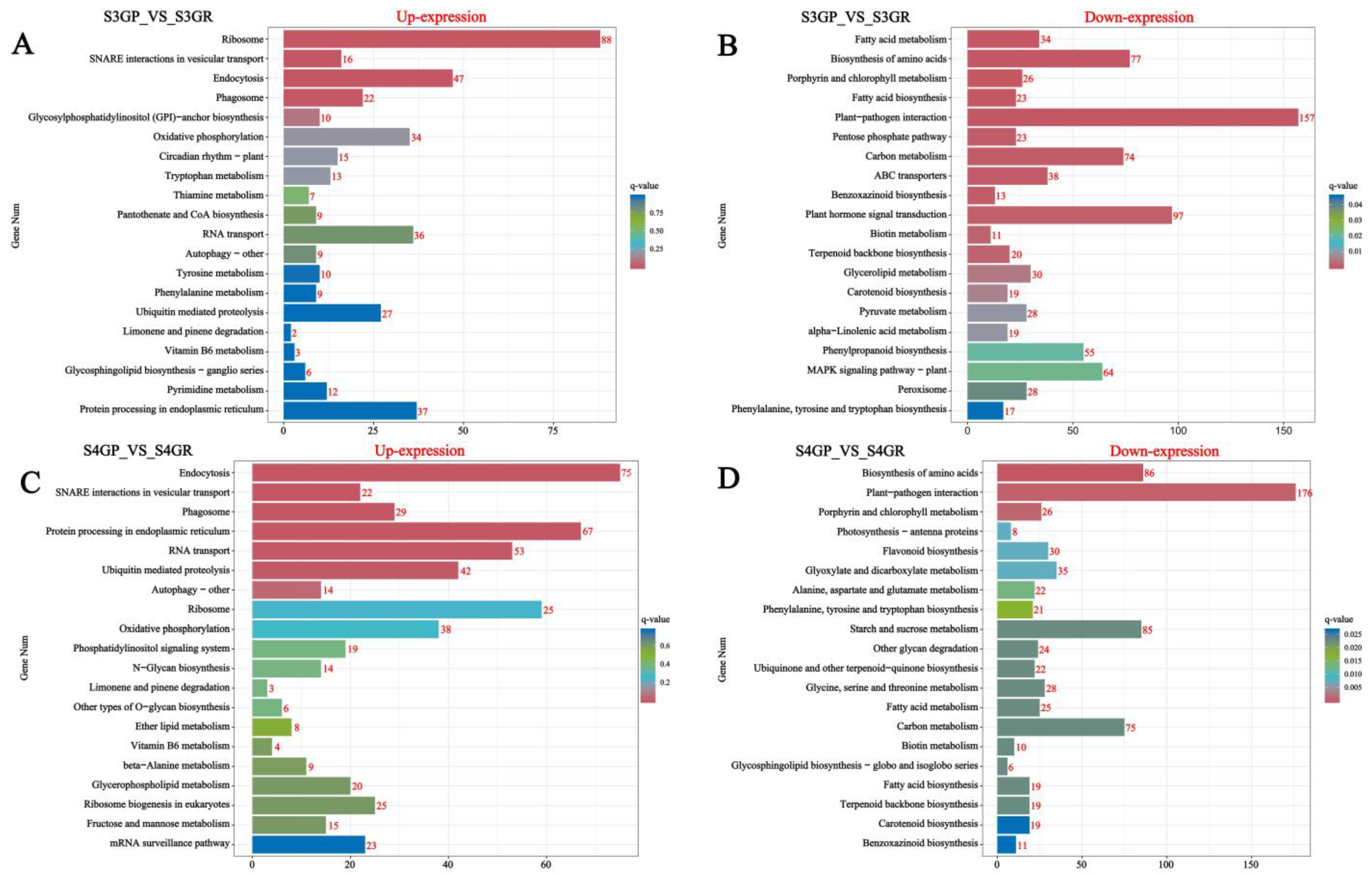
2.3.3. KEGG Annotation and Pathway Enrichment Analysis of Differentially Expressed Genes
2.3.4. Prediction of Genes Related to Organic Acid Metabolic Pathways
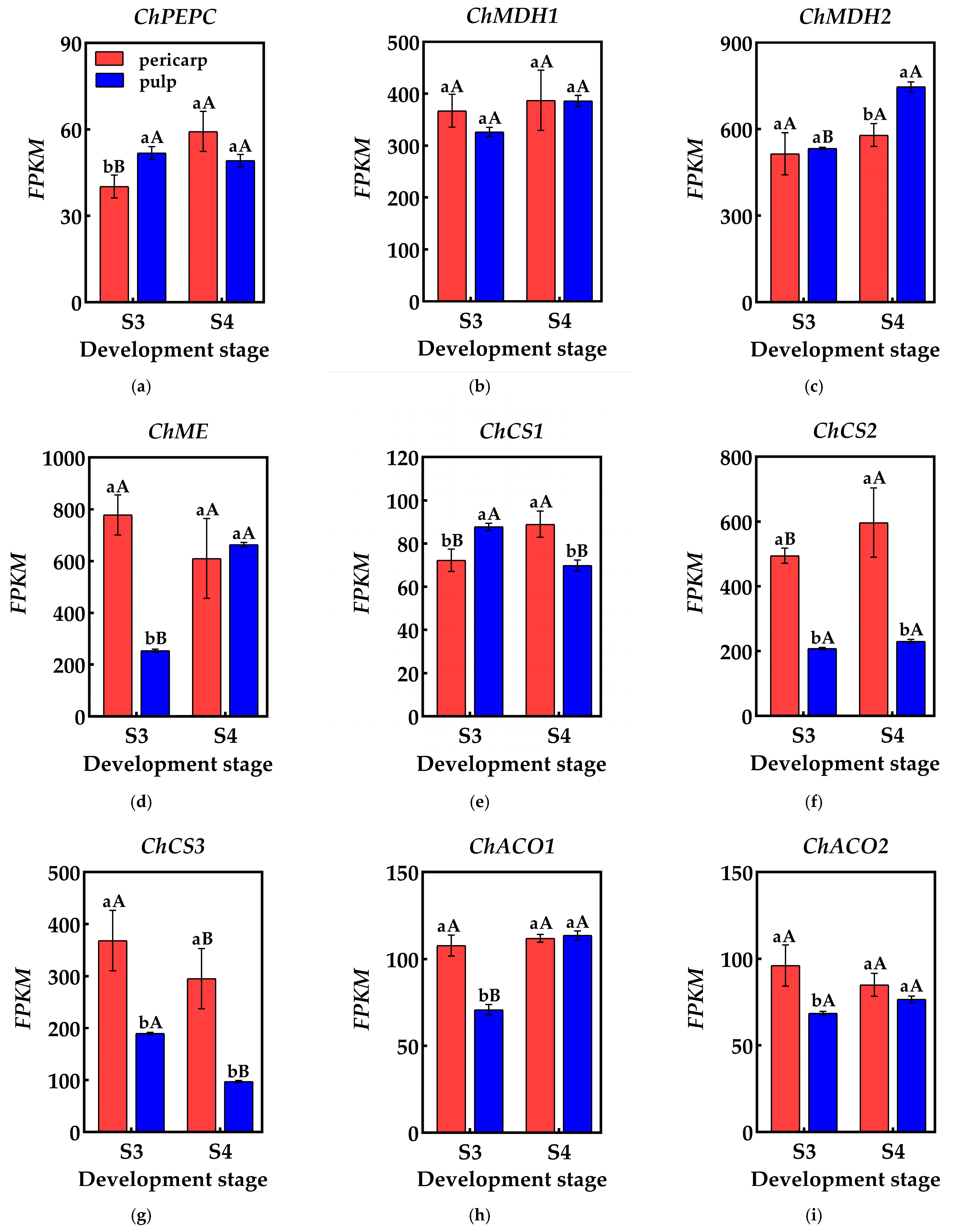
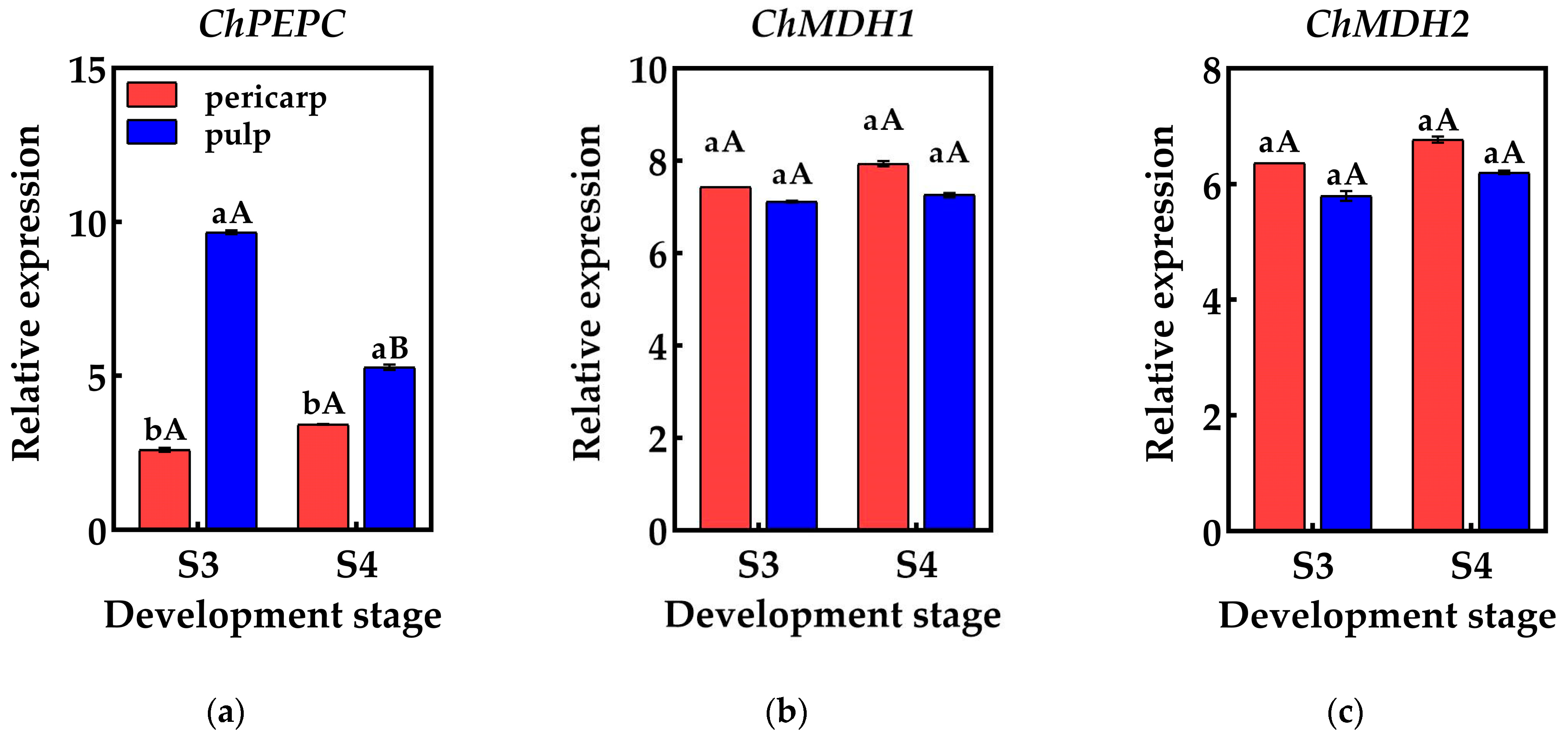
2.4. Expression Analysis of Genes Encoding Enzymes Related to Acid Metabolism
2.5. qRT-PCR Validation
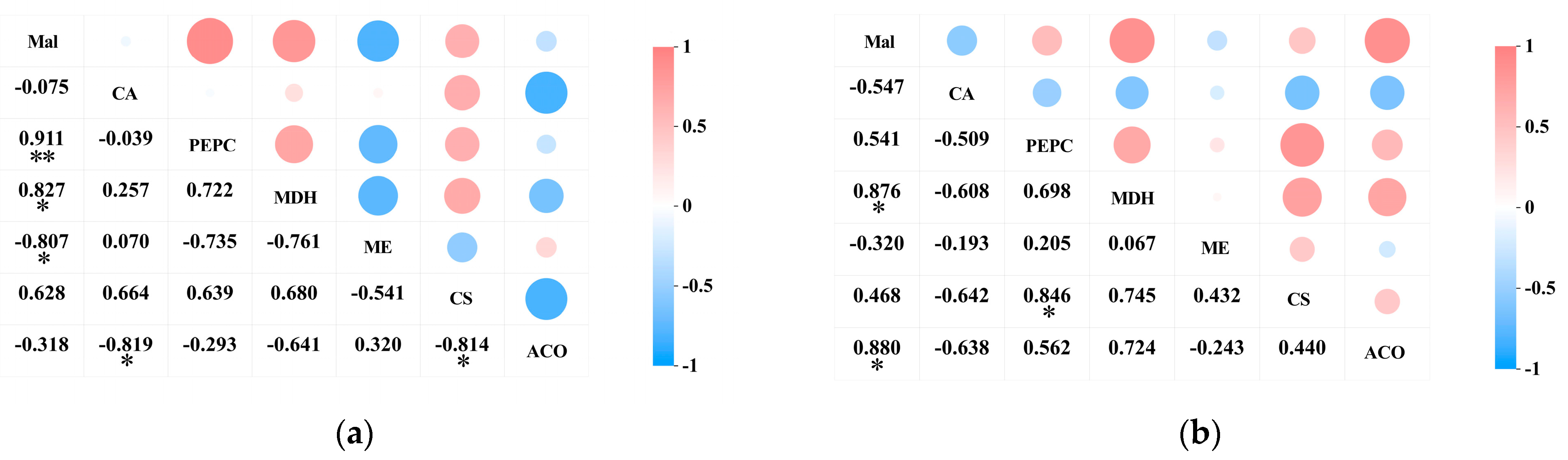
2.6. Correlation Analysis
2.6.1. The Correlation Between Acid Content in the Pericarp and the Pulp of Cerasus humilis and the Activity of Enzymes Involved in Acid Metabolism
2.6.2. Relationship Between the Activity of Acid Metabolism-Related Enzymes and the Expression of Their Encoding Genes in the Pericarp and the Pulp of Cerasus humilis
3. Discussion
3.1. Changes in Organic Acid Content in the Pericarp and the Pulp of Cerasus humilis During Development
3.2. Changes in Organic Acid Metabolism-Related Enzymes in the Pericarp and the Pulp of Cerasus humilis During Development
3.3. Acid Accumulation, Metabolic Enzymes, and Gene Expression Dynamics During Pericarp and Pulp Development in Cerasus humilis
4. Materials and Methods
4.1. Experimental Site
4.2. Sample Collection
4.3. Experimental Index Measurement
4.3.1. Determination of Organic Acids
4.3.2. Measurement of the Activity of Acid Metabolism-Related Enzymes
4.4. Correlation Analysis Methods
4.5. Screening of Genes Related to Organic Acid Metabolism Was Conducted Using Transcriptome Sequencing
4.5.1. Experimental Materials
4.5.2. RNA Extraction
4.5.3. Library Construction
4.5.4. RNA Quantification and Qualification
4.5.5. Library Preparation for Transcriptome Sequencing
4.5.6. Sequencing
Quality Control
Reads Mapping to the Reference Genome
Novel Transcripts Prediction and Gene Functional Annotation
Quantification of Gene Expression Levels
Differential Expression Analysis
GO Enrichment Analysis
KEGG Pathway Enrichment Analysis
4.5.7. RT-qPCR Validation
4.5.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shi, J.H.; Cao, J.H.; Xu, L.J. Evaluation and analysis of yield and nutritional content in Cerasus humilis. J. Hebei Sci. Technol. 2021, 2, 23–26. [Google Scholar]
- Li, W.D.; Gu, J.R. Research progress on healthy function and pharmacological effect in Cerasus humilis of fruit medicine dualpurpose type. Mod. Chin. Med. 2017, 19, 1336–1340. [Google Scholar]
- Cao, H.Q.; Liu, S.Y.; Wang, G.Y.; Liao, G.Z.; Bai, J. Nutritional composition and development of Prunus humilis Bunge. Farm. Prod. Process. 2015, 24, 70–72. [Google Scholar]
- Chen, F.X.; Liu, X.H.; Chen, L.S. Advances in research on organic acid metabolism in fruits. J. Fruit Sci. 2005, 22, 526–531. [Google Scholar]
- Koch, R.; Alleweldt, G. Der Gaswechsel reifender Weinbeeren. VITIS-J. Grapevine Res. 1978, 17, 30. [Google Scholar]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar]
- Mu, X.P.; Wang, P.F.; Du, J.J. Comparison of fruit organic acids and metabolism-related gene expression between Cerasus humilis (Bge.) Sok and Cerasus glandulosa (Thunb.) Lois. PLoS ONE 2018, 13, e0196537. [Google Scholar]
- Sun, D. Soluble Sugar and Organic Acid Contents Evalution of Different Citrus Germplasm Resources. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2013. [Google Scholar]
- Lamikanra, O.; Inyang, I.D.; Leong, S. Distribution and effect of grape maturity on organic acid content of red muscadine grapes. J. Agric. Food Chem. 1991, 43, 3026–3028. [Google Scholar]
- Monselise, S.P.; Galily, D. Organic acids in grapefruit fruit tissues. J. Am. Soc. Hortic. Sci. 1979, 104, 895–897. [Google Scholar]
- Saradhuldhat, P.; Paull, E.R. Pineapple organic acid metabolism and accumulation during fruit development. Sci. Hortic. 2006, 112, 297–303. [Google Scholar]
- Zhou, J.Y.; Zhang, K.; Wang, Y.; Song, P.H.; Zhang, H.F.; Luo, X.; Hao, Y.X.; Deng, Q.X. Comparison of malic acid metabolism between ’Guozhenhao 1’ and its parent ’Dawuxing’ loquat fruits. J. Sichuan Agric. Univ. 2024, 42, 546–553. [Google Scholar]
- Yang, G.K.; Xun, S.Y.; Li, J.Z. Analysis of organic acid components and malic acid metabolizing enzyme activity in Hongbaoshi apple fruits. J. Fruit Sci. 2023, 40, 884–892. [Google Scholar]
- Tang, X.; Huang, M.; Deng, L. Comparative transcriptome analysis reveals potential molecular regulation of organic acid metabolism during fruit development in Late-maturing hybrid citrus varieties. Int. J. Mol. Sci. 2025, 26, 803. [Google Scholar] [CrossRef]
- Chen, M.; Xie, X.; Lin, Q. Differential expression of organic acid degradation-related genes during fruit development of navel oranges (Citrus sinensis) in two habitats. Int. J. Mol. Sci. 2013, 31, 1131–1140. [Google Scholar] [CrossRef]
- Wang, P.F.; Xue, X.F.; Mu, X.P.; Zhang, J.C.; Cao, Q.; Du, J.J. Analysis of organic acid accumulation characteristics and organic acid-metabolizing enzyme activities of chinese dwarf cherry (Cerasus humilis Bunge) fruit. Sci. Agric. Sin. 2013, 46, 4101–4109. [Google Scholar]
- Tang, M.; Bie, Z.; Wu, M. Changes in organic acids and acid metabolism enzymes in melon fruit during development. Sci. Hortic. 2010, 123, 360–365. [Google Scholar]
- Zampini, M.; Wantling, E.; Phillips, N.; Spence, C. Multisensory flavor perception: Assessing the influence of fruit acids and color cues on the perception of fruit-flavored beverages. Food Qual. Prefer. 2008, 19, 335–343. [Google Scholar]
- Zhang, L.; Zhang, W.; Guo, J.L. Relationships between organic acid metabolism and the accumulation of sugars and calcium in fruits of Cerasus humilis during different development stages. Plants 2024, 13, 3053. [Google Scholar] [CrossRef]
- Fernie, R.A.; Carrari, F.; Sweetlove, J.L. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar]
- Wu, B.H.; Genard, M.; Lescourret, F.; Gomez, L.; Li, S.H. Influence of assimilate and water supply on seasonal variation of acids in peach (cv Suncrest). J. Sci. Food Agric. 2002, 82, 1829–1836. [Google Scholar]
- Zhang, X.X.; Wei, X.X.; Moaaz, A.M.; Muhammad, R.H.; Li, B.Q.; Li, H.; Jia, K.J.; Yang, X.L.; Ma, S.F.; Li, S.J.; et al. Changes in the content of organic acids and expression analysis of citric acid accumulation-related genes during fruit development of yellow (Passiflora edulis f. flavicarpa) and Purple (Passiflora edulis f. edulis) Passion Fruits. Int. J. Mol. Sci. 2021, 22, 5765. [Google Scholar] [PubMed]
- Tong, P.P.; Lu, D.Y.; Liao, G.L.; Wu, C.Y.; Wang, J.B. Identification and functional analysis of key genes regulating organic acid metabolism in jujube fruit. Agronomy 2024, 14, 2515. [Google Scholar]
- Moing, A.; Rothan, C.; Svanella, L.; Just, D.; Diakou, P.; Raymond, P.; Gaudillère, J.P.; Monet, R. Role of phospho enol pyruvate carboxylase in organic acid accumulation during peach fruit development. Physiol. Plant. 2000, 108, 1–10. [Google Scholar]
- Chen, L.; Qi, X.L.; Shi, C.S.; Dong, Y.X.; Song, L.L.; Liu, C.L.; Li, M. Advances in research of malate metabolism and regulation in fruit of hor ticultural crops. J. Fruit Sci. 2023, 40, 2598–2609. [Google Scholar]
- Sadka, A.; Dahan, E.; Cohen, L.; Marsh, B.M. Aconitase activity and expression during the development of lemon fruit. Physiol. Plant. 2000, 108, 255–262. [Google Scholar]
- Jiang, C.C.; Fang, Z.Z.; Zhou, D.R.; Pan, S.L.; Ye, X.F. Changes in secondary metabolites, organic acids and soluble sugars during the development of plum fruit cv. ’Furongli’ (Prunus salicina Lindl). J. Sci. Food Agric. 2019, 99, 1010–1019. [Google Scholar]
- Nour, V.; Trandafir, I.; Ionica, E.M. HPLC organic acid analysis in different citrus juices under reversed phase conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 44. [Google Scholar]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzbery, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEG Set | Total | COG | GO | KEGG | KOG | NR | Pfam | Swiss-Prot | eggNOG |
|---|---|---|---|---|---|---|---|---|---|
| S3GP_vs_S3GR | 5838 | 1871 | 4877 | 3906 | 3116 | 5833 | 4359 | 4143 | 4958 |
| S4GP_vs_S4GR | 7051 | 2162 | 5925 | 4778 | 3936 | 7043 | 5283 | 4979 | 6021 |
| Numbering of New Genes | Predicting Enzyme-Encoding Genes | Genes Name |
|---|---|---|
| NewGene_1191.1 | Malate dehydrogenase gene | ChMDH1 |
| NewGene_6957.1 | Malate dehydrogenase gene | ChMDH2 |
| NewGene_20719.2 | Malic enzyme gene | ChME |
| NewGene_10906.1 | Phosphoenolpyruvate carboxykinase gene | ChPEPC |
| NewGene_3681.1 | Aconitic acid hydratase gene | ChACO1 |
| NewGene_13406.1 | Aconitic acid hydratase gene | ChACO2 |
| NewGene_14300.1 | Citrate synthase gene | ChCS1 |
| NewGene_18025.5 | Citrate synthase gene | ChCS2 |
| NewGene_14300.1 | Citrate synthase gene | ChCS3 |
| Numbering of New Genes | Forward Primer | Reverse Primer |
|---|---|---|
| NewGene_1191.1 (ChMDH1) | gcg gtg ctg caa tca tca agg | aga agg aac att gta cga gcc atc a |
| NewGene_6957.1 (ChMDH2) | gaa gtt gga tgc aac cgc ag | aac tta gaa cca cgc cca gg |
| NewGene_20719.2 (ChME) | agg cga aga cca ttg tca ag | tca gaa atg gcg gca aaa gc |
| NewGene_10906.1 (ChPEPC) | gta aac gca gtg cag gca aa | cca gga gga acg aca agc at |
| NewGene_3681.1 (ChACO1) | atc aca act gac cac atc tca cct g | atc tca tca cca cgg cga ctt c |
| NewGene_13406.1 (ChACO2) | gta aac gca gtg cag gca aa | cca gga gga acg aca agc at |
| NewGene_14300.1 (ChCS1) | gct gct ctg aag caa cca ac | ctc gtg caa acg cga tca at |
| NewGene_18025.5 (ChCS2) | gag gtg gag atg ggt ggt tg | atg ctg ttc cca agt cgg tc |
| NewGene_14300.1 (ChCS3) | gga cta tgg tag ctg gag gag gt | gca caa tca atc aca act ctg gca t |
| Actin1 | gca gcg act gaa gac ata caa g | ggt ggc att agc aag ttc ctt |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, B.; Zhang, L.; Guo, J. Transcriptome Analysis Reveals Distinct Differences in Organic Acid Metabolism Between the Pericarp and the Pulp of Cerasus humilis During Fruit Maturation. Plants 2025, 14, 1105. https://doi.org/10.3390/plants14071105
Guo B, Zhang L, Guo J. Transcriptome Analysis Reveals Distinct Differences in Organic Acid Metabolism Between the Pericarp and the Pulp of Cerasus humilis During Fruit Maturation. Plants. 2025; 14(7):1105. https://doi.org/10.3390/plants14071105
Chicago/Turabian StyleGuo, Bingcheng, Li Zhang, and Jinli Guo. 2025. "Transcriptome Analysis Reveals Distinct Differences in Organic Acid Metabolism Between the Pericarp and the Pulp of Cerasus humilis During Fruit Maturation" Plants 14, no. 7: 1105. https://doi.org/10.3390/plants14071105
APA StyleGuo, B., Zhang, L., & Guo, J. (2025). Transcriptome Analysis Reveals Distinct Differences in Organic Acid Metabolism Between the Pericarp and the Pulp of Cerasus humilis During Fruit Maturation. Plants, 14(7), 1105. https://doi.org/10.3390/plants14071105