
Yield Determination in Major Small Grain Crops in Response to Nitrogen Fertilization

 , ,
, , 
Abstract
1. Introduction
2. Results
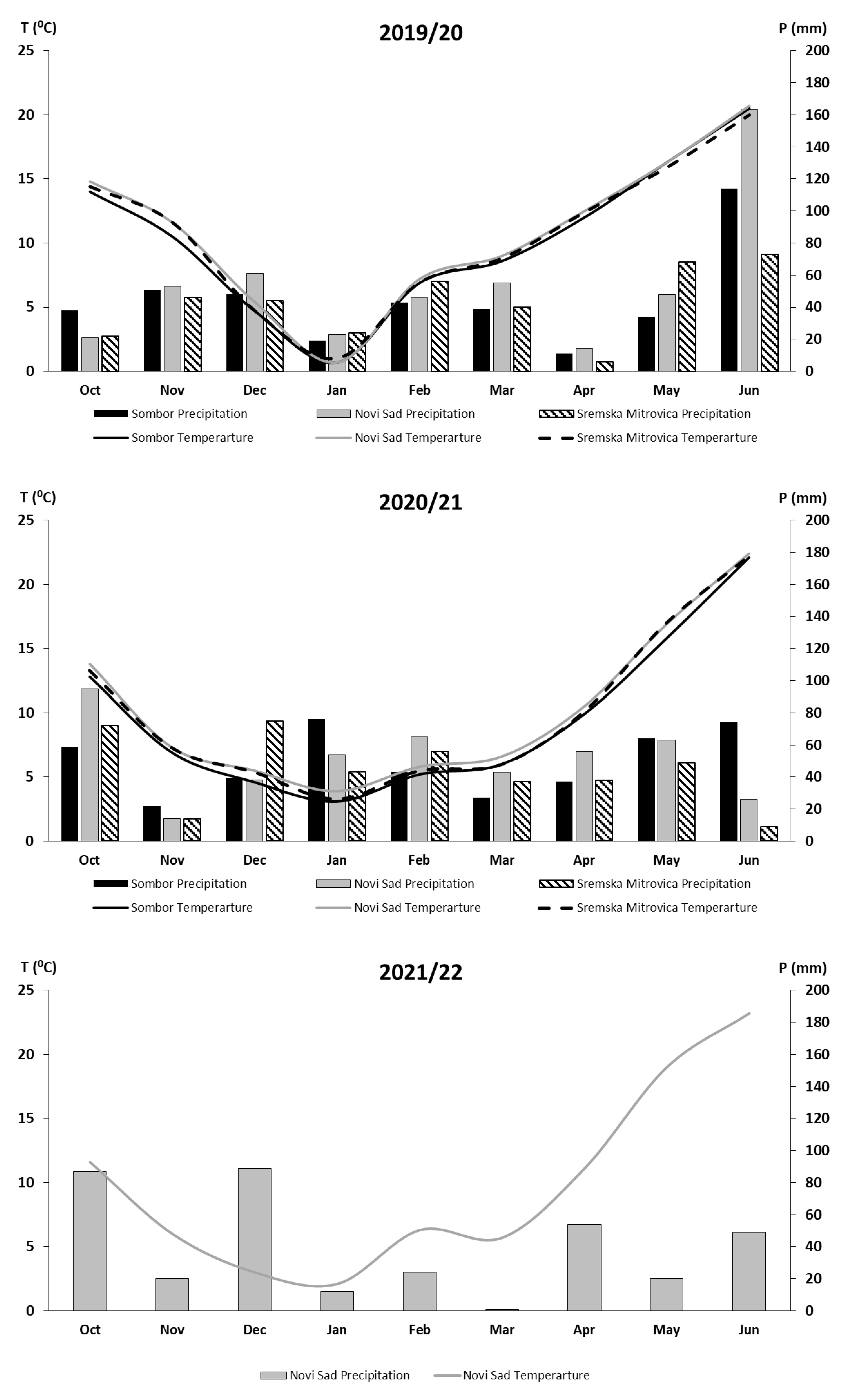
2.1. Weather Conditions
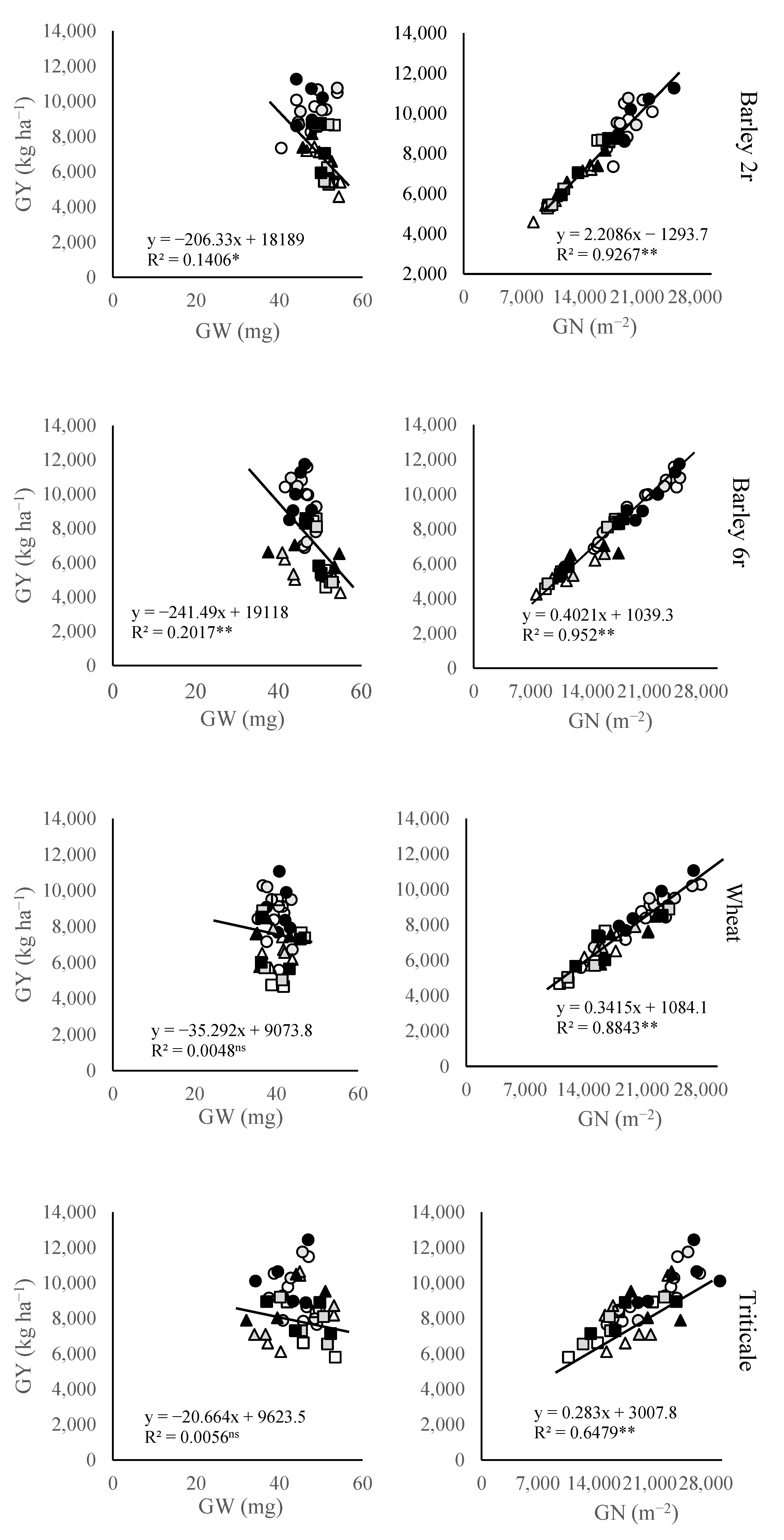
2.2. Grain Yield, Weight, and Number
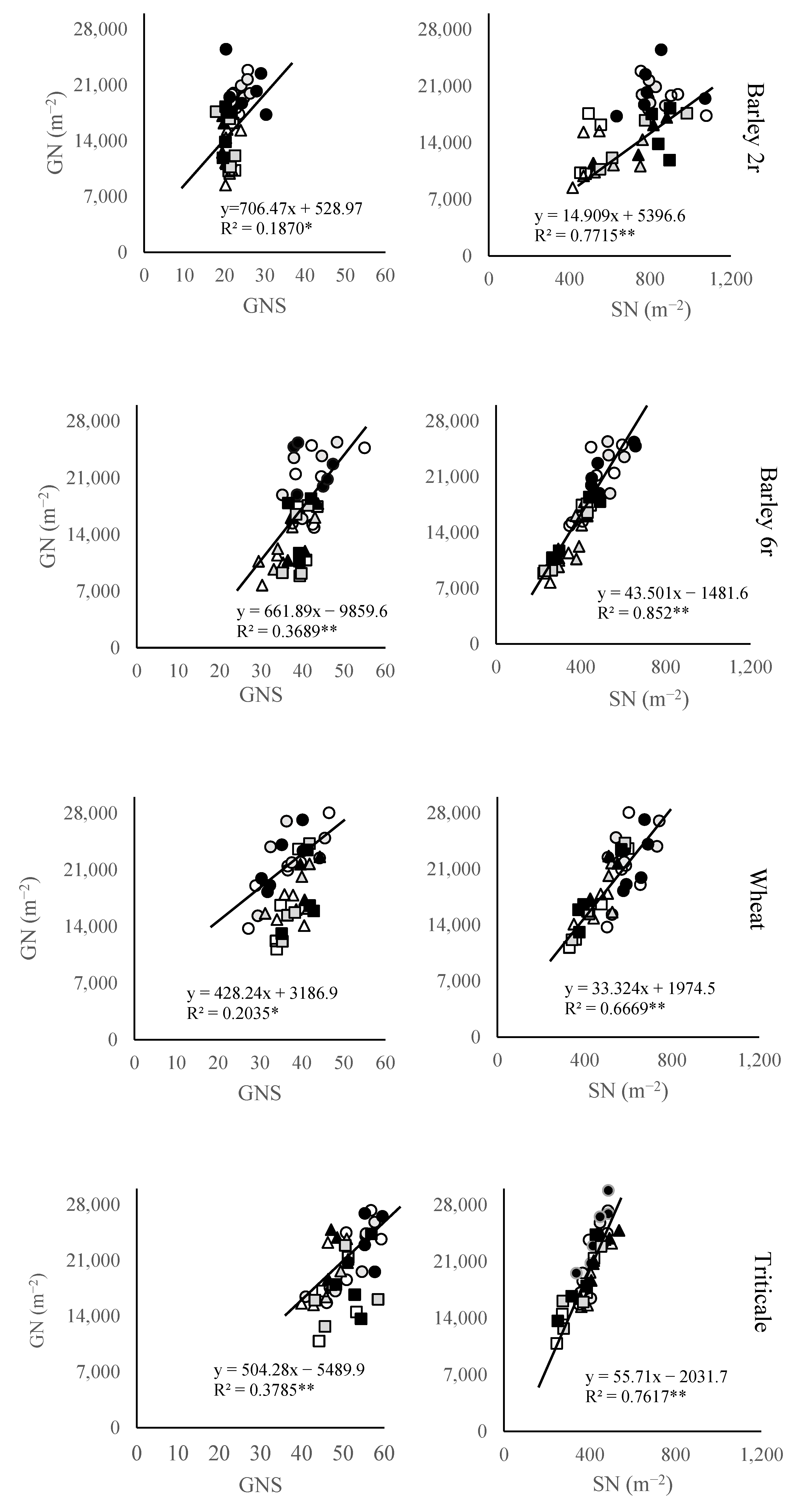
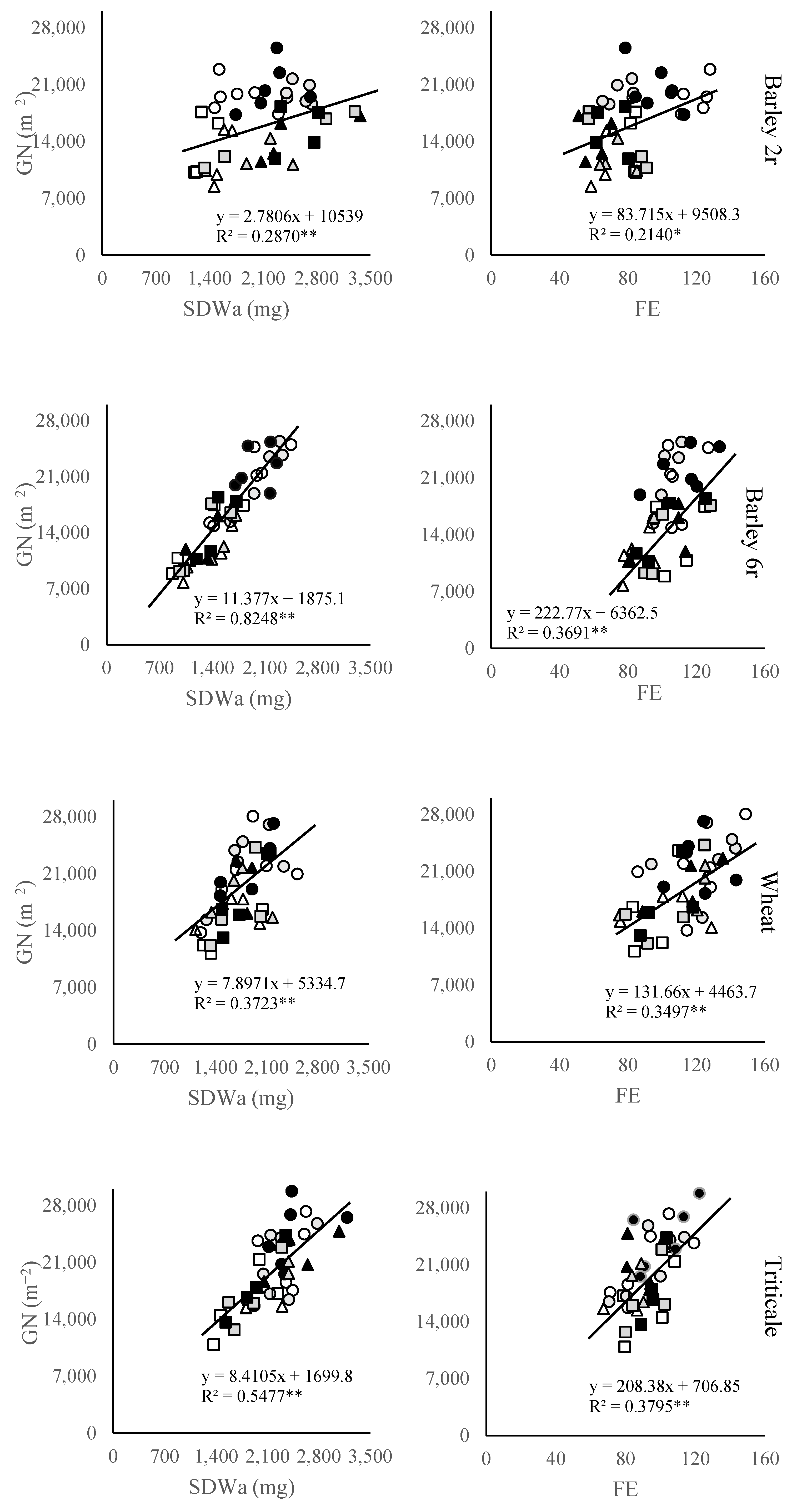
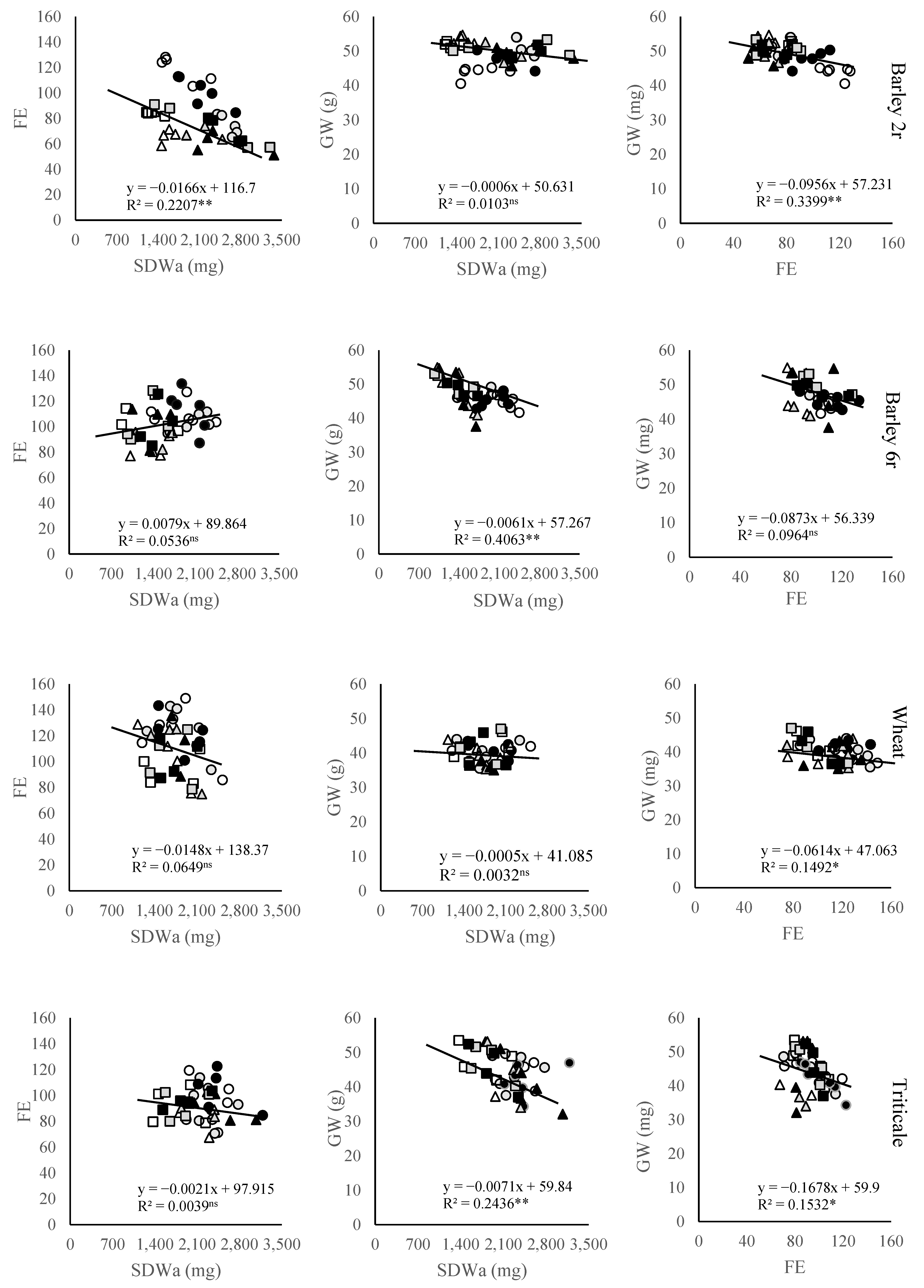
2.3. Grain Number Subcomponents
3. Discussion
3.1. Grain Yield Variation
3.2. Grain Number and Its Components
4. Materials and Methods
4.1. Experimental Site and Design
4.2. Crop Measurements and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GY | Grain yield |
| N | Nitrogen |
| GN | Number of grains per unit area |
| GW | Grain weight |
| SN | Spike number per unit area |
| GNS | Grain number per spike |
| FE | Fruiting efficiency |
| SDWa | Spike dry weight per area at anthesis |
References
- Mirosavljević, M.; Dražić, T.; Mikić, S.; Aćin, V.; Jocković, B.; Brbaklić, L.; Jaćimović, G. Nitrogen use efficiency in modern wheat cultivars affected by sowing dates. Cereal Res. Commun. 2024, 52, 1819–1829. [Google Scholar] [CrossRef]
- Dixon, J.; Braun, H.J.; Crouch, J. Transitioning wheat research to serve the future needs of the developing world. In Wheat Facts and Futures; Dixon, J., Braun, H.J., Kosina, P., Eds.; CIMMYT: Mexico City, Mexico, 2009; pp. 1–19. [Google Scholar]
- Estrada-Campuzano, G.; Slafer, G.A.; Miralles, D.J. Differences in yield, biomass and their components between triticale and wheat grown under contrasting water and nitrogen environments. Field Crops Res. 2012, 128, 167–179. [Google Scholar] [CrossRef]
- Giménez, V.D.; Serrago, R.A.; Abeledo, L.G.; Ciampitti, I.A.; Miralles, D.J. Comparative analysis of wheat and barley yield performance across temperate environments. Field Crops Res. 2024, 310, 109339. [Google Scholar] [CrossRef]
- Koppensteiner, L.J.; Kaul, H.P.; Piepho, H.P.; Barta, N.; Euteneuer, P.; Bernas, J.; Klimek-Kopyra, A.; Gronauer, A.; Neugschwandtner, R.W. Yield and yield components of facultative wheat are affected by sowing time, nitrogen fertilization and environment. Eur. J. Agron. 2022, 140, 126591. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B.L.; Fan, J.J.; Sun, M.; Yi, Y.; Guo, W.S.; Voldeng, H.D. Management of nitrogen fertilization to balance reducing lodging risk and increasing yield and protein content in spring wheat. Field Crops Res. 2019, 241, 107584. [Google Scholar] [CrossRef]
- Martínez-Dalmau, J.; Berbel, J.; Ordóñez-Fernández, R. Nitrogen fertilization. A review of the risks associated with the inefficiency of its use and policy responses. Sustainability 2021, 13, 5625. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Jamieson, P.D. Grain number, wheat yield, and bottling beer: An analysis. Field Crops Res. 2006, 98, 60–67. [Google Scholar] [CrossRef]
- Mirosavljević, M.; Momčilović, V.; Dražić, T.; Aćin, V.; Jocković, B.; Mikić, S.; Brbaklić, L.; Živančev, D.; Zorić, M.; Pržulj, N. Genetic progress in grain yield and associated changes in spikelet and grain traits in historical set of Pannonian wheat cultivars. Euphytica 2024, 220, 10. [Google Scholar] [CrossRef]
- Molero, G.; Joynson, R.; Pinera-Chavez, F.J.; Gardiner, L.J.; Rivera-Amado, C.; Hall, A.; Reynolds, M.P. Elucidating the genetic basis of biomass accumulation and radiation use efficiency in spring wheat and its role in yield potential. Plant Biotechnol. J. 2019, 17, 1276–1288. [Google Scholar] [CrossRef]
- Mirosavljević, M.; Momčilović, V.; Čanak, P.; Trkulja, D.; Mikić, S.; Jocković, B.; Pržulj, N. Grain filling variation in winter wheat, barley and triticale in Pannonian environments. Cereal Res. Commun. 2018, 46, 697–706. [Google Scholar] [CrossRef]
- Barraclough, P.B.; Howarth, J.R.; Jones, J.; Lopez-Bellido, R.; Parmar, S.; Shepherd, C.E.; Hawkesford, M.J. Nitrogen efficiency of wheat: Genotypic and environmental variation and prospects for improvement. Eur. J. Agron. 2010, 33, 1–11. [Google Scholar] [CrossRef]
- Beral, A.; Girousse, C.; Le Gouis, J.; Allard, V.; Slafer, G.A. Physiological bases of cultivar differences in average grain weight in wheat: Scaling down from plot to individual grain in elite material. Field Crops Res. 2022, 289, 108713. [Google Scholar] [CrossRef]
- Cossani, C.M.; Slafer, G.A.; Savin, R. Yield and biomass in wheat and barley under a range of conditions in a Mediterranean site. Field Crops Res. 2009, 112, 205–213. [Google Scholar] [CrossRef]
- Slafer, G.A.; Savin, R. Comparative performance of barley and wheat across a wide range of yielding conditions. Does barley outyield wheat consistently in low-yielding conditions? Eur. J. Agron. 2023, 143, 126689. [Google Scholar] [CrossRef]
- Prado, S.A.; Gallardo, J.M.; Kruk, B.C.; Miralles, D.J. Strategies for yield determination of bread wheat and two-row barley growing under different environments: A comparative study. Field Crops Res. 2017, 203, 94–105. [Google Scholar] [CrossRef]
- Mirosavljević, M.; Momčilović, V.; Denčić, S.; Mikić, S.; Trkulja, D.; Pržulj, N. Grain number and grain weight as determinants of triticale, wheat, two-rowed and six-rowed barley yield in the Pannonian environment. Span. J. Agric. Res. 2018, 16, e0903. [Google Scholar] [CrossRef]
- Méndez-Espinoza, A.M.; Romero-Bravo, S.; Estrada, F.; Garriga, M.; Lobos, G.A.; Castillo, D.; Matus, I.; Aranjuelo, I.; Del Pozo, A. Exploring agronomic and physiological traits associated with the differences in productivity between triticale and bread wheat in Mediterranean environments. Front. Plant Sci. 2019, 10, 404. [Google Scholar] [CrossRef]
- Motzo, R.; Pruneddu, G.; Virdis, A.; Giunta, G. Triticale vs. durum wheat: A performance comparison in a Mediterranean environment. Field Crops Res. 2015, 180, 63–71. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Savin, R.; Slafer, G.A.; Cossani, C.M.; Abeledo, L.G.; Sadras, V.O. Cereal yield in Mediterranean-type environments: Challenging the paradigms on terminal drought, the adaptability of barley vs. wheat and the role of nitrogen fertilization. In Crop Physiology; Academic Press: Cambridge, MA, USA, 2015; pp. 141–158. [Google Scholar]
- Pagnani, G.; Lorenzo, A.; Occhipinti, N.; Antonucci, L.; D’Egidio, S.; Stagnari, F.; Pisante, M. Effect of Flowering Shading on Grain Yield and Quality of Durum Wheat in a Mediterranean Environment. Plants 2024, 14, 76. [Google Scholar] [CrossRef]
- Roques, S.E.; Kindred, D.R.; Clarke, S. Triticale out-performs wheat on range of UK soils with a similar nitrogen requirement. J. Agric. Sci. 2017, 155, 261–281. [Google Scholar] [CrossRef]
- Bassu, S.; Asseng, S.; Richards, R. Yield benefits of triticale traits for wheat under current and future climates. Field Crops Res. 2011, 124, 14–24. [Google Scholar] [CrossRef]
- Asseng, S.; Turner, N.C. Modelling genotype × environment × management interactions to improve yield, water use efficiency and grain protein in wheat. In Scale and Complexity in Plant Systems Research; Springer: Dordrecht, The Netherlands, 2007; pp. 93–103. [Google Scholar]
- Lachutta, K.; Jankowski, K.J. An agronomic efficiency analysis of winter wheat at different sowing strategies and nitrogen fertilizer rates: A case study in northeastern Poland. Agriculture 2024, 14, 442. [Google Scholar] [CrossRef]
- Serrago, R.A.; García, G.A.; Savin, R.; Miralles, D.J.; Slafer, G.A. Determinants of grain number responding to environmental and genetic factors in two-and six-rowed barley types. Field Crops Res. 2023, 302, 109073. [Google Scholar] [CrossRef]
- Slafer, G.A.; García, G.A.; Serrago, R.A.; Miralles, D.J. Physiological drivers of responses of grains per m2 to environmental and genetic factors in wheat. Field Crops Res. 2022, 285, 108593. [Google Scholar] [CrossRef]
- Mirosavljević, M.; Momčilović, V.; Mikić, S.; Abicić, I.; Pržulj, N. Breeding progress in grain filling and grain yield components of six-rowed winter barley. Zemdirb.-Agric. 2020, 107, 271–278. [Google Scholar] [CrossRef]
- Gauley, A.; Boden, S.A. Genetic pathways controlling inflorescence architecture and development in wheat and barley. J. Integr. Plant Biol. 2019, 61, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Schnurbusch, T. A genetic playground for enhancing grain number in cereals. Trends Plant Sci. 2012, 17, 91–101. [Google Scholar] [CrossRef]
- Mirosavljević, M.; Mikić, S.; Kondić-Špika, A.; Župunski, V.; Zhou, R.; Abdelhakim, L.; Ottosen, C.O. The effect of heat stress on some main spike traits in 12 wheat cultivars at anthesis and mid-grain filling stage. Plant Soil Environ. 2021, 67, 71–76. [Google Scholar] [CrossRef]
- Arisnabarreta, S.; Miralles, D.J. Floret development and grain setting in near isogenic two-and six-rowed barley lines (Hordeum vulgare L.). Field Crops Res. 2006, 96, 466–476. [Google Scholar] [CrossRef]
- de San Celedonio, R.P.; Abeledo, L.G.; Miralles, D.J. Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant Soil 2014, 378, 265–277. [Google Scholar] [CrossRef]
- Koppolu, R.; Schnurbusch, T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. J. Int. Plant Biol. 2019, 61, 278–295. [Google Scholar] [CrossRef]
- Sadras, V.O.; Slafer, G.A. Environmental modulation of yield components in cereals: Heritabilities reveal a hierarchy of phenotypic plasticities. Field Crops Res. 2012, 127, 215–224. [Google Scholar] [CrossRef]
- Ferrante, A.; Savin, R.; Slafer, G.A. Floret development of durum wheat in response to nitrogen availability. J. Exp. Bot. 2010, 61, 4351–4359. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Wu, Y.; Zheng, X.; Guo, X.; Sun, W.; Sun, Z.; Wang, Z.; Zhang, Y. A dynamic regulation of nitrogen on floret primordia development in wheat. Crop J. 2024, 12, 271–280. [Google Scholar] [CrossRef]
- Slafer, G.A.; Elia, M.; Savin, R.; García, G.A.; Terrile, I.I.; Ferrante, A.; Miralles, D.J.; González, F.G. Fruiting efficiency: An alternative trait to further rise wheat yield. Food Energy Secur. 2015, 4, 92–109. [Google Scholar] [CrossRef]
- Pretini, N.; Alonso, M.P.; Vanzetti, L.S.; Pontaroli, A.C.; González, F.G. The physiology and genetics behind fruiting efficiency: A promising spike trait to improve wheat yield potential. J. Exp. Bot. 2021, 72, 3987–4004. [Google Scholar] [CrossRef]
- Gerard, G.S.; Alqudah, A.; Lohwasser, U.; Börner, A.; Simón, M.R. Uncovering the genetic architecture of fruiting efficiency in bread wheat: A viable alternative to increase yield potential. Crop Sci. 2019, 59, 1853–1869. [Google Scholar] [CrossRef]
- Xu, X.; Liu, S.; Meng, F.; Zhang, X.; Zhao, J.; Qu, W.; Shi, Y.; Zhao, C. Grain yield formation and nitrogen utilization efficiency of different winter wheat varieties under rainfed conditions in the Huang-Huai-Hai Plain. Agronomy 2023, 13, 915. [Google Scholar] [CrossRef]
- Benincasa, P.; Reale, L.; Cerri, M.; Tedeschini, E.; Tosti, G.; Falcinelli, B.; Rosati, A. Nitrogen fertilization levels and timing affect the plasticity of yield components in bread wheat (Triticum aestivum L.). Field Crops Res. 2022, 289, 108734. [Google Scholar] [CrossRef]
- Ferrante, A.; Savin, R.; Slafer, G.A. Differences in yield physiology between modern, well adapted durum wheat cultivars grown under contrasting conditions. Field Crops Res. 2012, 136, 52–64. [Google Scholar] [CrossRef]
- Ferrante, A.; Cartelle, J.; Savin, R.; Slafer, G.A. Yield determination, interplay between major components and yield stability in a traditional and a contemporary wheat across a wide range of environments. Field Crops Res. 2017, 203, 114–127. [Google Scholar] [CrossRef]
- Elía, M.; Savin, R.; Slafer, G.A. Fruiting efficiency in wheat: Physiological aspects and genetic variation among modern cultivars. Field Crops Res. 2016, 191, 83–90. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Wehrmann, J.; Scharpf, H.C. Mineral Nitrogen Concentration of Soil as a Gauge of Need for Nitrogen-Fertilizer (N-Min Method). Plant Soil 1979, 52, 109–126. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Novi Sad | Sombor | Sremska Mitrovica | ||||
|---|---|---|---|---|---|---|---|
| Coordinate | 45°20′ N, 19°51′ E | 45°48′ N, 19°07′ E | 44°56′ N, 19°49′ E | ||||
| Growing season | 2019/20 | 2020/21 | 2021/22 | 2019/20 | 2020/21 | 2019/20 | 2020/21 |
| Sowing date | 14 October | 18 October | 27 October | 21 October | 15 October | 9 October | 23 October |
| Harvest date | 3 July 2020 | 29 June 2021 | 27 June 2022 | 07 July 2020 | 28 June 2021 | 06 July 2020 | 30 June 2021 |
| Some properties of the soil | |||||||
| CaCO3, % | 5.17 | 5.09 | 5.37 | 5.45 | 6.12 | 4.30 | 4.45 |
| P2O5 (Egner–Riehm), mg/100 g | 24.5 | 23.1 | 24.9 | 19.8 | 17.4 | 22.5 | 24.2 |
| K2O (Egner–Riehm), mg/100 g | 18.4 | 23.5 | 20.3 | 30.1 | 28.7 | 23.9 | 21.8 |
| N (N-min method), kg ha−1 | 85 | 77 | 77 | 75 | 83 | 70 | 90 |
| Humus (Turin) | 2.75 | 2.92 | 2.89 | 2.35 | 2.40 | 2.48 | 2.56 |
| pH (KCl) | 7.26 | 7.41 | 7.35 | 7.75 | 7.88 | 7.35 | 7.42 |
| Applied P (kg ha−1) | 70 | 60 | 70 | 80 | 80 | 70 | 60 |
| Applied K (kg ha−1) | 50 | 40 | 50 | 40 | 40 | 40 | 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirosavljević, M.; Momčilović, V.; Aćin, V.; Jocković, B.; Pržulj, N.; Jaćimović, G. Yield Determination in Major Small Grain Crops in Response to Nitrogen Fertilization. Plants 2025, 14, 1017. https://doi.org/10.3390/plants14071017
Mirosavljević M, Momčilović V, Aćin V, Jocković B, Pržulj N, Jaćimović G. Yield Determination in Major Small Grain Crops in Response to Nitrogen Fertilization. Plants. 2025; 14(7):1017. https://doi.org/10.3390/plants14071017
Chicago/Turabian StyleMirosavljević, Milan, Vojislava Momčilović, Vladimir Aćin, Bojan Jocković, Novo Pržulj, and Goran Jaćimović. 2025. "Yield Determination in Major Small Grain Crops in Response to Nitrogen Fertilization" Plants 14, no. 7: 1017. https://doi.org/10.3390/plants14071017
APA StyleMirosavljević, M., Momčilović, V., Aćin, V., Jocković, B., Pržulj, N., & Jaćimović, G. (2025). Yield Determination in Major Small Grain Crops in Response to Nitrogen Fertilization. Plants, 14(7), 1017. https://doi.org/10.3390/plants14071017