
Mechanisms of Increase of Winter Wheat Frost Resistance Under Tebuconazole Treatment at Early Stage of Growth: Role of Hormone- and Reactive Oxygen Species-Mediated Signaling Pathways

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Results
2.1. Carotenoids Content
2.2. Growth Parameters Under Control and Cold Hardening
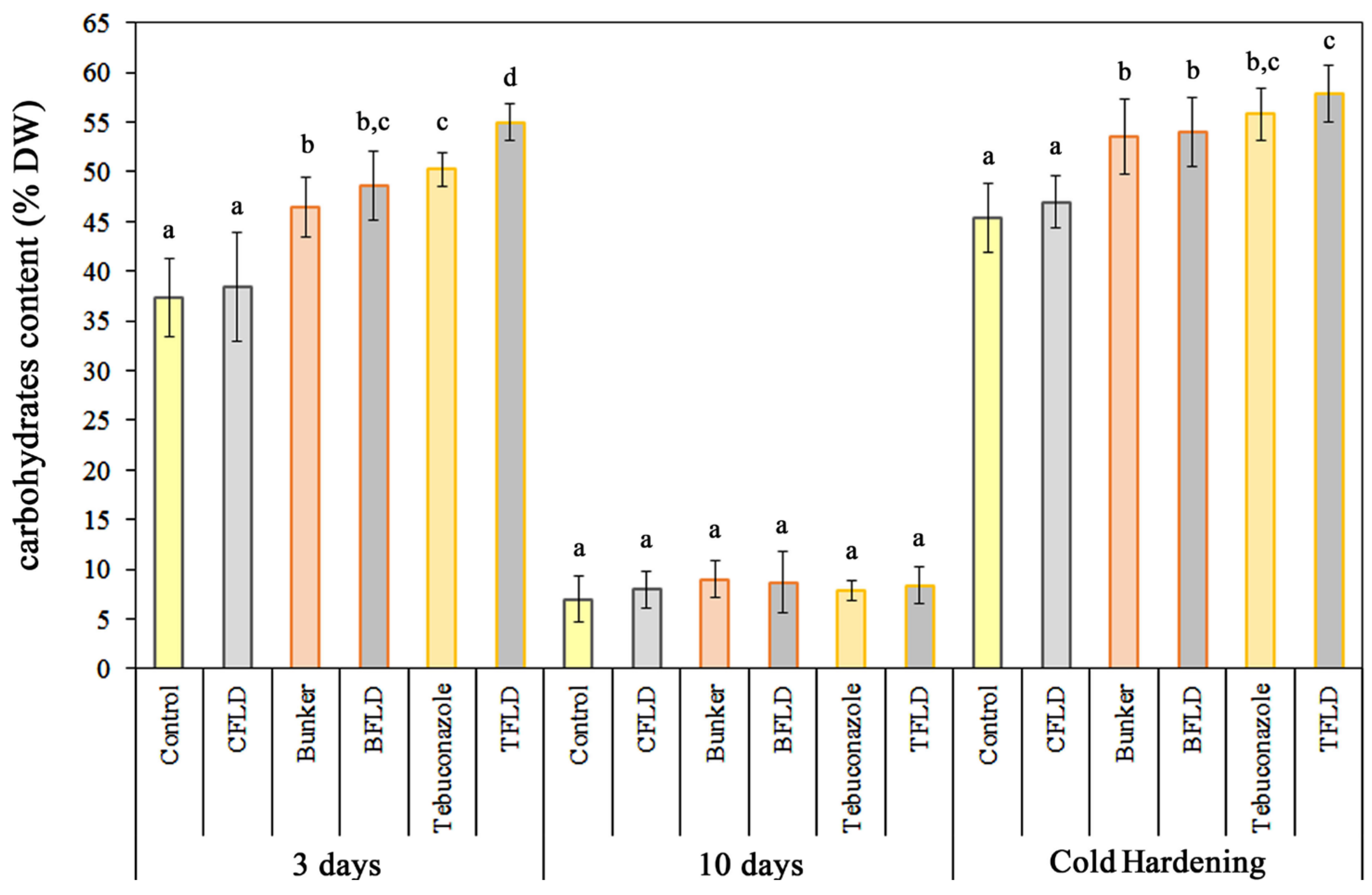
2.3. Water-Soluble Carbohydrate Content in Shoots Under Control and Cold Hardening
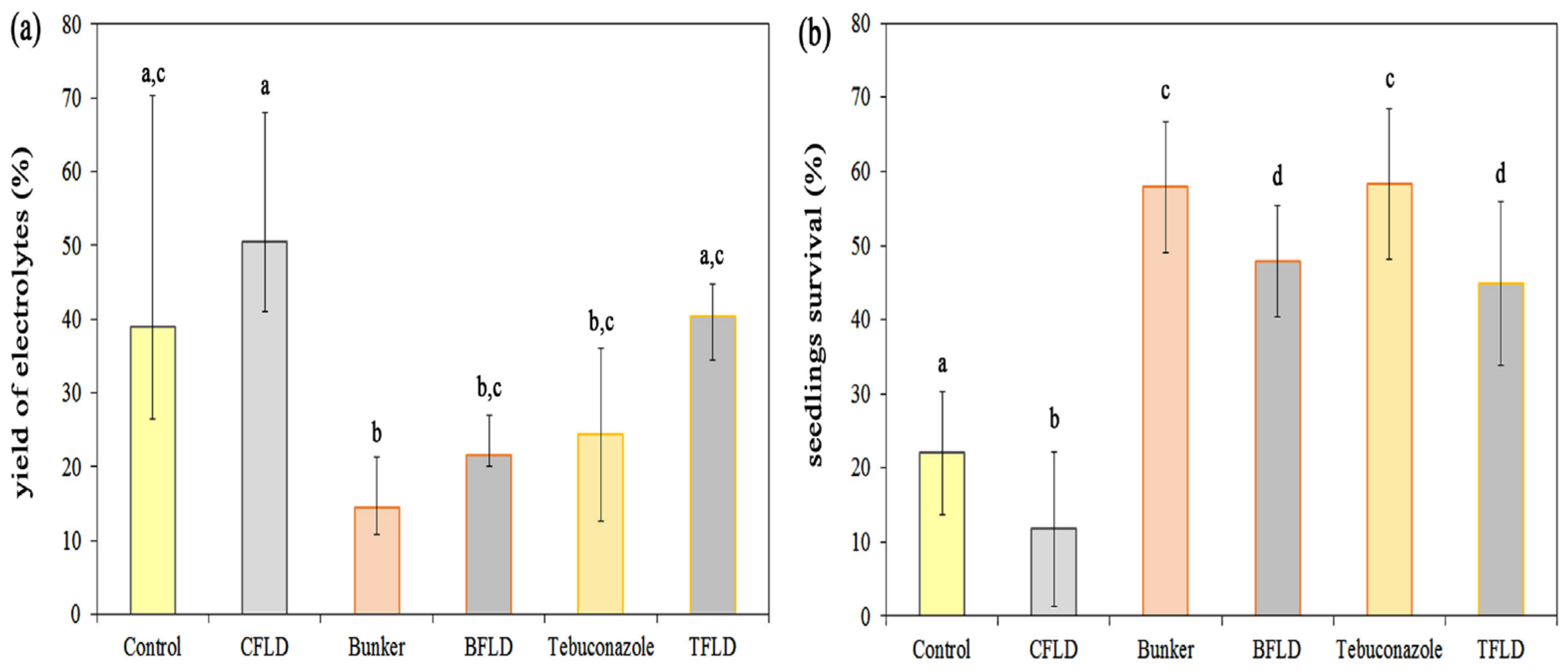
2.4. Frost Resistance of Seedlings
2.5. Hydrogen Peroxide Content, Lipid Peroxidation, and Antioxidant Enzyme Activity Under Control and Cold Hardening
2.6. Endogenous Hormone Content Under Control and Cold Hardening
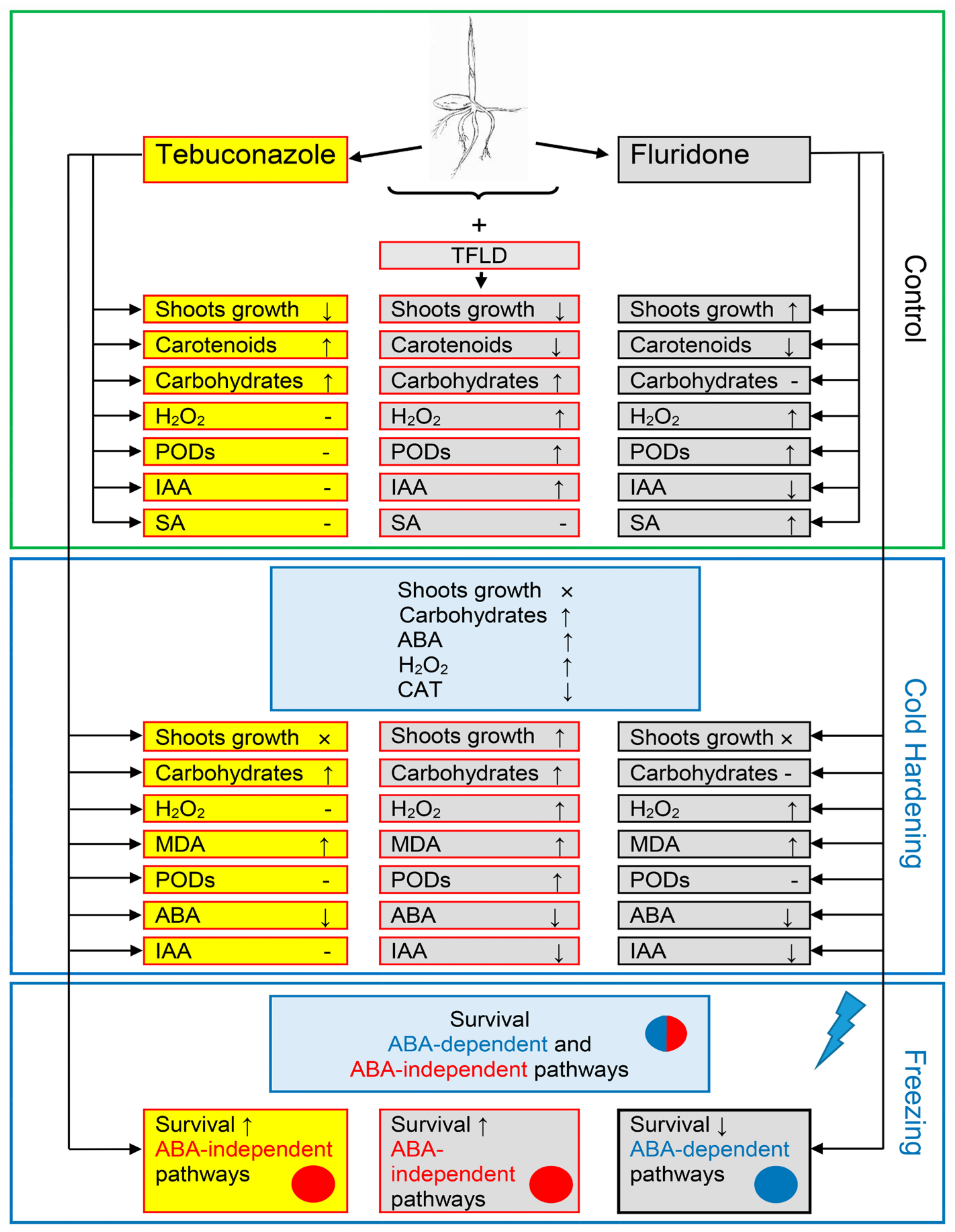
3. Discussion
4. Materials and Methods
4.1. Plant Material, Fungicide and Inhibitor Treatments, and Growth Conditions
4.2. Temperature Treatments and Determination of Frost Resistance of Seedlings
4.3. Determination of Carotenoids Content
4.4. Determination of Water-Soluble Carbohydrate Content
4.5. Determination of Hydrogen Peroxide and Lipid Peroxidation
4.6. Determination of the Catalase and Guaiacol Peroxidase Activity
4.7. Determination of the Content of Endogenous Hormones
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Prášil, I.T.; Vítámvás, P.; Dobrev, P.; Motyka, V.; Floková, K.; Novák, O.; Turečková, V.; Rolčik, J.; Pešek, B.; et al. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J. Plant Physiol. 2012, 169, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Klíma, M.; Prášil, I.T.; Vítámvás, P. COR/LEA proteins as indicators of frost tolerance in Triticeae: A comparison of controlled versus field conditions. Plants 2021, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, I.; Pu, X.; Naseer, M.A.; Bohoussou, Y.N.D.; Liu, Y.; Farooq, M.; Zhang, J.; Zhang, Y.; Wang, Z.; Sun, Z. Cold and drought stresses in wheat: A global meta-analysis of 21st century. J. Plant Growth Regul. 2023, 42, 5379–5395. [Google Scholar] [CrossRef]
- Hameed, A.; Farooq, T. Chapter 7—Triazole-based plant growth-regulating agents: A recent update. In Advances in Triazole Chemistry; Elsevier: Amsterdam, The Netherlands, 2021; pp. 169–185. [Google Scholar] [CrossRef]
- Hof, H. Critical annotations to the use of azole antifungals for plant protection. Antimicrob. Agents Chemother. 2001, 45, 2987–2990. [Google Scholar] [CrossRef]
- Popov, S.Y.; Dorozhkina, L.A.; Kalinin, V.A. Fundamentals of Chemical Plant Protection, 1st ed.; Art-Lion: Moscow, Russia, 2003; pp. 102–107. (In Russian) [Google Scholar]
- Chizhova, S.I.; Pavlova, V.V.; Prusakova, L.D. Abscisic acid content and growth of spring barley plants treated with triazoles. Russ. J. Plant Physiol. 2005, 52, 93–98. [Google Scholar] [CrossRef]
- Vettakkorumakankav, N.N.; Falk, D.; Saxena, P.; Fletcher, R.A. A crucial role for gibberellins in stress protection of plants. Plant Cell Physiol. 1999, 40, 542–548. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef]
- Saito, S.; Okamoto, M.; Shinoda, S.; Kushiro, T.; Koshiba, T.; Kamiya, Y.; Hirai, N.; Todoroki, Y.; Sakata, K.; Nambara, E.; et al. A plant growth retardant, uniconazole, is a potent inhibitor of ABA catabolism in Arabidopsis. Biosci. Biotechnol. Biochem. 2006, 70, 1731–1739. [Google Scholar] [CrossRef]
- Soumya, P.R.; Kumar, P.; Pal, M. Paclobutrazol: A novel plant growth regulator and multi-stress ameliorant. Indian J. Plant Physiol. 2017, 22, 267–278. [Google Scholar] [CrossRef]
- Tafazoli, E.; Bey, C. Changes in endogenous abscisic acid and cold hardiness in Actinidia treated with triazole growth retardants. J. Plant Growth Regul. 1993, 12, 79–83. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, Q.; Zhang, B.; Zhang, F.; Liu, P.; Zhou, S.; Liu, X. Hormonal and enzymatic responses of maize seedlings to chilling stress as affected by triazoles seed treatments. Plant Physiol. Biochem. 2020, 148, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Geng, R.; Kong, X.; Li, L.; Zhang, Z.; Liu, R. Transcriptomic and physiological properties reveal the tolerance mechanism to difenoconazole toxicity in wheat (Triticum aestivum L.). Ecotoxicol. Environ. Saf. 2023, 255, 114787. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, S.M.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Fujita, M. Exogenous tebuconazole and trifloxystrobin regulates reactive oxygen species metabolism toward mitigating salt-induced damages in cucumber seedling. Plants 2019, 8, 428. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Nahar, K.; Hossain, S.; Bhuyan, B.; Parvin, K.; Fujita, M. Tebuconazole and trifloxystrobin regulate the physiology, antioxidant defense and methylglyoxal detoxification systems in conferring salt stress tolerance in Triticum aestivum L. Physiol. Mol. Biol. Plants 2020, 26, 1139–1154. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Hossain, S.; Fujita, M. Protective role of tebuconazole and trifloxystrobin in wheat (Triticum aestivum L.) under cadmium stress via enhancement of antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2021, 27, 1043–1057. [Google Scholar] [CrossRef]
- Morrison, M.J.; Andrews, C.J. Variable increases in cold hardiness induced in winter rape by plant growth regulators. J. Plant Growth Regul. 1992, 11, 113–117. [Google Scholar] [CrossRef]
- Namjoyan, S.; Rajabi, A.; Sorooshzadeh, A.; AghaAlikhani, M. The potential of tebuconazole for mitigating oxidative stress caused by limited irrigation and improving sugar yield and root quality traits in sugar beet. Plant Physiol. Biochem. 2021, 162, 547–555. [Google Scholar] [CrossRef]
- Pobezhimova, T.P.; Korsukova, A.V.; Dorofeev, N.V.; Grabelnych, O.I. Physiological effects of triazole fungicides in plants. Proc. Univ. Appl. Chem. Biotechnol. 2019, 9, 461–476. (In Russian) [Google Scholar] [CrossRef]
- Upadhyaya, A.; Davis, T.D.; Walser, R.H.; Galbraith, A.B.; Sankhla, N. Uniconazole-induced alleviation of low-temperature damage in relation to antioxidant activity. Hort. Sci. 1989, 24, 955–957. [Google Scholar] [CrossRef]
- Rademacher, W. Plant growth regulators: Backgrounds and uses in plant production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Ijaz, M.; Mahmood, K.; Honermeier, B. Interactive role of fungicides and plant growth regulator (Trinexapac) on seed yield and oil quality of winter rapeseed. Agronomy 2015, 5, 435–446. [Google Scholar] [CrossRef]
- Matysiak, K.; Kaczmarek, S. Effect of chlorocholine chloride and triazoles—Tebuconazole and flusilazole on winter oilseed rape (Brassica napus var. oleifera L.) in response to the application term and sowing density. J. Plant Prot. Res. 2013, 53, 79–88. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Zhang, X.; Chen, C.-J.; Zhou, M.-G.; Wang, H.-C. Effects of fungicides JS399-19, azoxystrobin, tebuconazloe, and carbendazim on the physiological and biochemical indices and grain yield of winter wheat. Pestic. Biochem. Physiol. 2010, 98, 151–157. [Google Scholar] [CrossRef]
- Korsukova, A.V.; Borovik, O.A.; Grabelnych, O.I.; Voinikov, V.K. The tebuconazole-based protectant of seeds “Bunker” induces the synthesis of dehydrins during cold hardening and increases the frost resistance of wheat seedlings. J. Stress Physiol. Biochem. 2015, 11, 118–127. (In Russian) [Google Scholar]
- Korsukova, A.V.; Grabelnych, O.I.; Borovik, O.A.; Pobezhimova, T.P.; Voinikov, V.K.; Dorofeev, N.V. The influence of the treatment of seeds by tebuconazole on the carbohydrates content and frost resistance of winter wheat and winter rye. Agric. Chem. 2016, 7, 52–58. (In Russian) [Google Scholar]
- Pobezhimova, T.P.; Korsukova, A.V.; Borovik, O.A.; Zabanova, N.S.; Dorofeev, N.V.; Grabelnych, O.I.; Voinikov, V.K. The influence of tebuconazole and tebuconazole-based protectant “Bunker” on the functioning of winter wheat mitochondria. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2020, 37, 224–234. (In Russian) [Google Scholar] [CrossRef]
- Lazukin, A.; Pinchuk, M.; Korsukova, A.; Nikiforov, A.; Romanov, G.; Stepanova, O.; Grabelnych, O. Comparison of presowing wheat treatments by low-temperature plasma, electric field, cold hardening, and action of tebuconazole-based protectant. Appl. Sci. 2022, 12, 6447. [Google Scholar] [CrossRef]
- Gusta, L.V.; Trischuk, R.; Weiser, C.J. Plant cold acclimation: The role of abscisic acid. J. Plant Growth Regul. 2005, 24, 308–318. [Google Scholar] [CrossRef]
- Gamble, P.E.; Mullet, J.E. Inhibition of carotenoid accumulation and abscisic acid biosynthesis in fluridone-treated dark-grown barley. Eur. J. Biochem. 1986, 160, 117–121. [Google Scholar] [CrossRef]
- Stetsenko, L.A.; Vedenicheva, N.P.; Likhnevsky, R.V.; Kuznetsov, V.V. Influence of abscisic acid and fluridone on the content of phytohormones and polyamines and the level of oxidative stress in plants of Mesembryanthemum crystallinum L. under salinity. Biol. Bull. 2015, 42, 98–107. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Pál, M. Salicylic acid signalling in plants. Int. J. Mol. Sci. 2020, 21, 2655. [Google Scholar] [CrossRef] [PubMed]
- Rock, C.D.; Sakata, Y.; Quatrano, R.S. Stress signaling I: The role of abscisic acid (ABA). In Abiotic Stress Adaptation in Plants, 1st ed.; Pareek, A., Sopory, S.K., Bohnert, H.J., Govindjee, G., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 33–73. [Google Scholar] [CrossRef]
- Xue-Xuan, X.; Hong-Bo, S.; Yuan-Yuan, M.; Gang, X.; Jun-Na, S.; Dong-Gang, G.; Cheng-Jiang, R. Biotechnological implications from abscisic acid (ABA) roles in cold stress and leaf senescence as an important signal for improving plant sustainable survival under abiotic-stressed conditions. Crit. Rev. Biotechnol. 2010, 30, 222–230. [Google Scholar] [CrossRef]
- Deepthi, V.P. Triazoles as plant growth regulators and stress protectants in fruit crops: A review. Int. J. Agric. Sci. Vet. Med. 2016, 4, 53–64. [Google Scholar]
- Ahmad, I.; Kamran, M.; Ali, S.; Cai, T.; Bilegjargal, B.; Liu, T.; Han, Q. Seed filling in maize and hormones crosstalk regulated by exogenous application of uniconazole in semiarid regions. Environ. Sci. Pollut. Res. 2018, 25, 33225–33229. [Google Scholar] [CrossRef]
- Grabelnych, O.I.; Polyakova, E.A.; Korsukova, A.V.; Zabanova, N.S.; Berezhnaya, E.V.; Lyubushkina, I.V.; Fedotova, O.A.; Stepanov, A.V.; Pobezhimova, T.P.; Dorofeev, N.V. Differently directional effects of tebuconazole-based protectant of seeds “Bunker” on the growth of winter wheat shoots and roots. Bull. Irkutsk. State Univ. Ser. Biol. Ecol. 2020, 34, 3–19. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gampala, S.S.L.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell. 2002, 14, 15–45. [Google Scholar] [CrossRef]
- Liu, L.; Cang, J.; Yu, J.; Wang, X.; Huang, R.; Wang, J.; Lu, B. Effects of exogenous abscisic acid on carbohydrate metabolism and the expression levels of correlative key enzymes in winter wheat under low temperature. Biosci. Biotechnol. Biochem. 2013, 77, 516–525. [Google Scholar] [CrossRef]
- Borovik, O.A.; Pomortsev, A.V.; Korsukova, A.V.; Polyakova, E.A.; Fomina, E.A.; Zabanova, N.S.; Grabelnych, O.I. Effect of cold acclimation and deacclimation on the content of soluble carbohydrates and dehydrins in the leaves of winter wheat. J. Stress Physiol. Biochem. 2019, 15, 62–67. [Google Scholar]
- Bakht, J.; Bano, A.; Dominy, P. The role of abscisic acid and low temperature in chickpea (Cicer arietinum) cold tolerance. II. Effects on plasma membrane structure and function. J. Exp. Bot. 2006, 57, 3707–3715. [Google Scholar] [CrossRef]
- Chen, W.P.; Li, P.H. Membrane stabilization by abscisic acid under cold aids proline in alleviating chilling injury in maize (Zea mays L.) cultured cells. Plant Cell Environ. 2002, 25, 955–962. [Google Scholar] [CrossRef]
- Allagulova, C.R.; Maslennikova, D.R.; Avalbaev, A.M.; Fedorova, K.A.; Yuldashev, R.A.; Shakirova, F.M. Influence of 24-epibrassinolide on growth of wheat plants and the content of dehydrins under cadmium stress. Russ. J. Plant Physiol. 2015, 62, 465–471. [Google Scholar] [CrossRef]
- Kondhare, K.R.; Hedden, P.; Kettlewell, P.S.; Farrell, A.D.; Monaghan, J.M. Use of the hormone-biosynthesis inhibitors fluridone and paclobutrazol to determine the effects of altered abscisic acid and gibberellin levels on pre-maturity a-amylase formation in wheat grains. J. Cereal. Sci. 2014, 60, 210–216. [Google Scholar] [CrossRef]
- Hoffmann-Benning, S.; Kende, H. On the role of abscisic acid and gibberellin in the regulation of growth in rice. Plant Physiol. 1992, 99, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, N.; Yan, X.; Shi, J.; Zhang, M.; Wang, Z.; Yuan, H. Microencapsulation of seed-coating tebuconazole and its effects on physiology and biochemistry of maize seedlings. Colloids Surf. B Biointerfaces 2014, 114, 241–246. [Google Scholar] [CrossRef]
- Shaki, F.; Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Role of triazolic compounds in underlying mechanisms of plant stress tolerance: A Review. Iran. J. Plant Physiol. 2022, 12, 3943–3954. [Google Scholar]
- Emenecker, R.J.; Strader, L.C. Auxin-abscisic acid interactions in plant growth and development. Biomolecules 2020, 10, 281. [Google Scholar] [CrossRef]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef]
- Eremina, M.; Rozhon, W.; Poppenberger, B. Hormonal control of cold stress responses in plants. Cell Mol. Life Sci. 2016, 73, 797–810. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, S.; Saha, B.; Awasthi, J.; Dey, M.; Panda, S.K.; Sahooet, L. Crosstalk between salt, drought, and cold stress in plants: Toward genetic engineering for stress tolerance. In Abiotic Stress in Plants; Wiley: Hoboken, NJ, USA, 2016; pp. 55–86. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kidokoro, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulatory networks in plant responses to drought and cold stress. Plant Physiol. 2024, 195, 170–189. [Google Scholar] [CrossRef]
- Richter, R.; Bastakis, E.; Schwechheimer, C. Cross-repressive interactions between SOC1 and the GATAs GNC and GNL/CGA1 in the control of greening, cold tolerance, and flowering time in Arabidopsis. Plant Physiol. 2013, 162, 1992–2004. [Google Scholar] [CrossRef] [PubMed]
- Richter, R.; Behringer, C.; Müller, I.K.; Schwechheimer, C. The GATA-type transcription factors GNC and GNL/CGA1 repress gibberellin signaling downstream from DELLA proteins and PHYTOCHROME-INTERACTING FACTORS. Genes Dev. 2010, 24, 2093–2104. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Liu, C.; Ren, W.; Li, B.; Lü, Y.; Pan, Z.; Cao, W. Flower and pod development, grain-setting characteristics and grain yield in Chinese milk vetch (Astragalus sinicus L.) in response to pre-anthesis foliar application of paclobutrazol. PLoS ONE 2021, 16, e0245554-1. [Google Scholar] [CrossRef]
- Zhang, S.-W.; Li, C.-H.; Cao, J.; Zhang, Y.-C.; Zhang, S.-Q.; Xia, Y.-F.; Sun, D.-Y.; Sun, Y. Altered Architecture and Enhanced Drought Tolerance in Rice via the Down-Regulation of Indole-3-Acetic Acid by TLD1/OsGH3.13 Activation. Plant Physiol. 2009, 151, 1889–1901. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Q.; Li, Z.; Staswick, P.E.; Wang, M.; Zhu, Y.; He, Z. Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-Pseudomonas syringae interaction. Plant Physiol. 2007, 145, 450–464. [Google Scholar] [CrossRef]
- Du, W.; Lu, Y.; Li, Q.; Luo, S.; Shen, S.; Li, N.; Chen, X. TIR1/AFB proteins: Active players in abiotic and biotic stress signaling. Sec. Plant Abiotic Stress 2022, 13, 1083409. [Google Scholar] [CrossRef]
- Calderón Villalobos, L.I.A.; Lee, S.; De Oliveira, C.; Ivetac, A.; Brandt, W.; Armitage, L.; Sheard, L.B.; Tan, X.; Parry, G.; Mao, H.; et al. A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 2012, 8, 477–485. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef]
- Wojtaczka, P.; Ciarkowska, A.; Starzynska, E.; Ostrowski, M. The GH3 amidosynthetases family and their role in metabolic crosstalk modulation of plant signaling compounds. Phytochemistry 2022, 194, 113039. [Google Scholar] [CrossRef]
- Ng, S.; De Clercq, I.; Van Aken, O.; Law, S.R.; Ivanova, A.; Willems, P.; Giraud, E.; Van Breusegem, F.; Whelan, J. Anterograde and retrograde regulation of nuclear genes encoding mitochondrial proteins during growth, development, and stress. Mol. Plant 2014, 7, 1075–1093. [Google Scholar] [CrossRef]
- Van Aken, O. Mitochondrial redox systems as central hubs in plant metabolism and signaling. Plant Physiol. 2021, 186, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.; Law, S.R.; Narsai, R.; Duncan, O.; Lee, J.H.; Zhang, B.; Van Aken, O.; Radomiljac, J.D.; Van der Merwe, M.; Yi, K.; et al. Functional antagonistic relationship between auxin and mitochondrial retrograde signaling regulates Alternative Oxidase1a expression in Arabidopsis. Plant Physiol. 2014, 165, 1233–1254. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Baloch, A.M. Overview of sustainable plant growth and differentiation and the role of hormones in controlling growth and development of plants under various stresses. Recent Pat. Food Nutr. Agric. 2020, 11, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fu, X.; Feng, Y.; Zhang, X.; Bi, H.; Ai, X. Abscisic acid mediates salicylic acid induced chilling tolerance of grafted cucumber by activating H2O2 biosynthesis and accumulation. Int. J. Mol. Sci. 2022, 23, 16057. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Hu, H.; Fan, B.; Zhu, C.; Chen, Z. Biosynthesis and roles of salicylic acid in balancing stress response and growth in plants. Int. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Jardim-Messender, D.; Margis-Pinheiro, M.; Sachetto-Martins, G. Salicylic acid and adenine nucleotides regulate the electron transport system and ROS production in plant mitochondria. Biochem. Biophys. Acta 2022, 1863, 148559. [Google Scholar] [CrossRef]
- Quinlan, C.L.; Orr, A.L.; Perevoshchikova, I.V.; Treberg, J.R.; Ackrell, B.A.; Brand, M.D. Mitochondrial complex II can generate reactive oxygen species at high rates in both the forward and reverse reactions. J. Biol. Chem. 2012, 287, 27255–27264. [Google Scholar] [CrossRef]
- Song, W.; Shao, H.; Zheng, A.; Zhao, L.; Xu, Y. Advances in roles of salicylic acid in plant tolerance responses to biotic and abiotic stresses. Plants 2023, 12, 3475. [Google Scholar] [CrossRef]
- Borovik, O.A.; Grabelnych, O.I. Mitochondrial alternative cyanide-resistant oxidase is involved in an increase of heat stress tolerance in spring wheat. J. Plant Physiol. 2018, 231, 310–317. [Google Scholar] [CrossRef]
- Gavrilenko, V.F.; Zhigalova, T.V. Large Workshop on Photosynthesis; Academa: Moscow, Russia, 2003; pp. 54–55. (In Russian) [Google Scholar]
- Lyubushkina, I.V.; Polyakova, M.S.; Pomortsev, A.V.; Korsukova, A.V.; Zabanova, N.S.; Pobezhimova, T.P.; Grabel’nykh, O.I.; Voinikov, V.K. Efficiency of androgenesis in vitro in the culture of isolated winter wheat anthers: 1. The influence of the duration of low-temperature pretreatment of donor plants on respiration, carbohydrate content, and ROS levels in flowers from different parts of the spike. Russian J. Plant Physiol. 2023, 70, 213. [Google Scholar] [CrossRef]
- Bellincampi, D.; Dipierro, N.; Salvi, G.; Cervone, F.; De Lorenzo, G. Extracellular H2O2 induced by oligogalacturonides is not involved in the inhibition of the auxin-regulated rolB gene expression in tobacco leaf explants. J. Plant Physiol. 2000, 122, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Garmash, E.V.; Velegzhaninov, I.O.; Grabelnych, O.I.; Borovik, O.A.; Silina, E.V.; Voinikov, V.K.; Golovko, T.K. Expression profiles of genes for mitochondrial respiratory energy-dissipating systems and antioxidant enzymes in wheat leaves during de-etiolation. J. Plant Physiol. 2017, 215, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Fedoseeva, I.V.; Gamburg, K.Z.; Varakina, N.N.; Rusaleva, T.M.; Tauson, E.L.; Stupnikova, I.V.; Borovskii, G.B.; Stepanov, A.V.; Davydenko, E.A.; Rikhvanov, E.G.; et al. The effect of sodium azide and 2,4-dinitrophenol on the development of thermotolerance and induction of Hsp101 in cultured Arabidopsis thaliana cells. Russ. J. Plant Physiol. 2008, 55, 225–231. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Rudikovskii, A.V.; Stolbikova, A.V.; Rudikovskaya, E.G.; Dudareva, L.V. Role of phytogormones in the formation of dwarf and tall Siberian crabapple (Malus baccata L. Borkh.). Zemdirb.-Agric. 2019, 106, 167–172. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants | Growth Period, Days | Cold Hardening | ||
|---|---|---|---|---|
| 3 | 5 | 10 | ||
| Coleoptile, mm | ||||
| Control | 39 [35; 43] c | 49 [46; 53] d | 46 [43; 48] d | 42 [38; 45] c |
| CFLD | 45 [42; 47] d | 58 [54; 62] e | 50 [46; 53] e | 46 [41; 49] d |
| Bunker | 21 [18; 24] a | 34 [31; 37] a, b | 27 [25; 29] a, b | 22 [20; 25] a |
| BFLD | 24 [22; 27] b | 38 [36; 43] a | 30 [28; 33] c | 25 [24; 29] b |
| Tebuconazole | 19 [13; 22] a | 28 [26; 30] c | 25 [23; 27] a | 22 [17; 24] a |
| TFLD | 27 [23; 29] b | 31 [29; 33] b, c | 30 [25; 32] b, c | 29 [26; 31] b |
| Shoot, mm | ||||
| Control | 37 [31; 40] c | 83 [75; 95] b | 156 [131; 178] a | 40 [34; 45] c |
| CFLD | 39 [36; 43] d | 93 [85; 104] c | 182 [145; 211] b | 36 [30; 41] a |
| Bunker | 21 [18; 24] a | 62 [58; 67] a | 138 [121; 149] c | 22 [20; 26] b |
| BFLD | 25 [22; 27] b | 71 [65; 78] d | 165 [148; 181] a, b | 27 [24; 30] d |
| Tebuconazole | 19 [13; 22] a | 63 [52; 71] a | 125 [111; 137] d | 23 [17; 27] b |
| TFLD | 27 [23; 29] b | 79 [69; 85] e | 151 [136; 165] a | 35 [32; 37] a |
| Variant | Growth Period, Days | Cold Hardening | |
|---|---|---|---|
| 3 | 10 | ||
| Fresh Weight, g | |||
| Control | 1.047 ± 0.034 a | 1.624 ± 0.667 a | 1.042 ± 0.025 a |
| CFLD | 1.016 ± 0.046 a | 1.591 ± 0.624 a | 0.965 ± 0.138 a |
| Bunker | 0.573 ± 0.114 b | 1.721 ± 0.359 a | 0.638 ± 0.229 a |
| BFLD | 0.633 ± 0.098 b | 1.734 ± 0.507 a | 0.788 ± 0.280 a |
| Tebuconazole | 0.480 ± 0.142 c | 2.760 ± 0.020 b | 0.616 ± 0.149 b |
| TFLD | 0.823 ± 0.033 d | 2.462 ± 0.393 b | 0.998 ± 0.211 c |
| Dry Weight, g | |||
| Control | 0.0871 ± 0.0036 a | 0.1490 ± 0.0588 a | 0.1200 ± 0.0115 a |
| CFLD | 0.0944 ± 0.0078 a | 0.1510 ± 0.0610 a | 0.1250 ± 0.0281 a |
| Bunker | 0.0591 ± 0.0094 b | 0.1710 ± 0.0354 a | 0.0889 ± 0.0361 a |
| BFLD | 0.0667 ± 0.0069 b | 0.1680 ± 0.0517 a | 0.1080 ± 0.0437 a |
| Tebuconazole | 0.0555 ± 0.0111 c | 0.2750 ± 0.0059 b | 0.0879 ± 0.0139 b |
| TFLD | 0.0842 ± 0.0036 d | 0.2500 ± 0.0412 b | 0.1260 ± 0.0268 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korsukova, A.V.; Lyubushkina, I.V.; Zabanova, N.S.; Berezhnaya, E.V.; Polyakova, E.A.; Pobezhimova, T.P.; Kirichenko, K.A.; Dorofeev, N.V.; Dudareva, L.V.; Grabelnych, O.I. Mechanisms of Increase of Winter Wheat Frost Resistance Under Tebuconazole Treatment at Early Stage of Growth: Role of Hormone- and Reactive Oxygen Species-Mediated Signaling Pathways. Plants 2025, 14, 314. https://doi.org/10.3390/plants14030314
Korsukova AV, Lyubushkina IV, Zabanova NS, Berezhnaya EV, Polyakova EA, Pobezhimova TP, Kirichenko KA, Dorofeev NV, Dudareva LV, Grabelnych OI. Mechanisms of Increase of Winter Wheat Frost Resistance Under Tebuconazole Treatment at Early Stage of Growth: Role of Hormone- and Reactive Oxygen Species-Mediated Signaling Pathways. Plants. 2025; 14(3):314. https://doi.org/10.3390/plants14030314
Chicago/Turabian StyleKorsukova, Anna V., Irina V. Lyubushkina, Natalya S. Zabanova, Ekaterina V. Berezhnaya, Elizaveta A. Polyakova, Tamara P. Pobezhimova, Kuzma A. Kirichenko, Nikolay V. Dorofeev, Lyubov V. Dudareva, and Olga I. Grabelnych. 2025. "Mechanisms of Increase of Winter Wheat Frost Resistance Under Tebuconazole Treatment at Early Stage of Growth: Role of Hormone- and Reactive Oxygen Species-Mediated Signaling Pathways" Plants 14, no. 3: 314. https://doi.org/10.3390/plants14030314
APA StyleKorsukova, A. V., Lyubushkina, I. V., Zabanova, N. S., Berezhnaya, E. V., Polyakova, E. A., Pobezhimova, T. P., Kirichenko, K. A., Dorofeev, N. V., Dudareva, L. V., & Grabelnych, O. I. (2025). Mechanisms of Increase of Winter Wheat Frost Resistance Under Tebuconazole Treatment at Early Stage of Growth: Role of Hormone- and Reactive Oxygen Species-Mediated Signaling Pathways. Plants, 14(3), 314. https://doi.org/10.3390/plants14030314