

Phosphorus Fertilization and Chemical Root Pruning: Effects on Root Traits During the Nursery Stage in Two Mediterranean Species from Central Chile
 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results
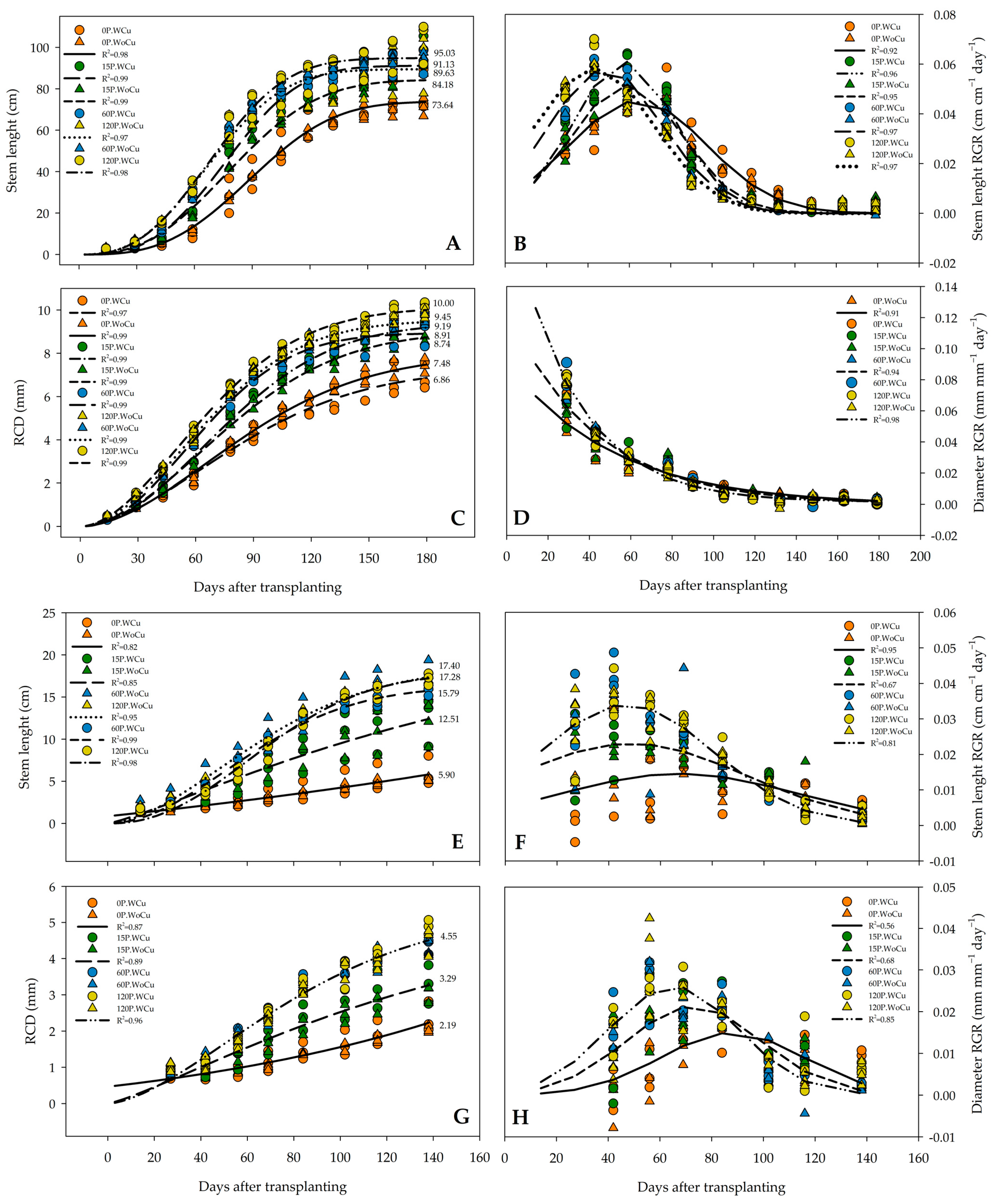
2.1. Morphological Attributes
2.2. Nutrient Concentration and Content Analysis
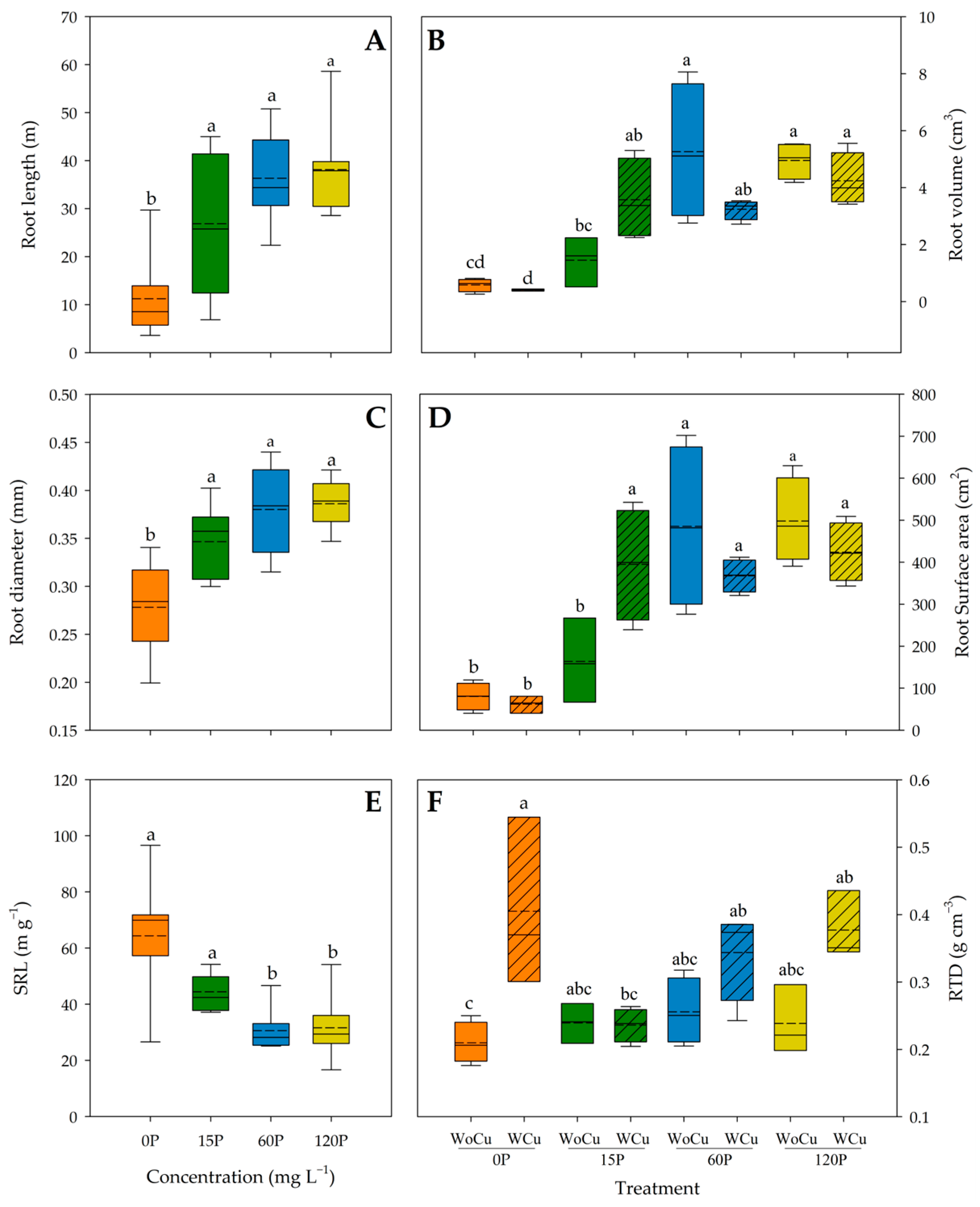
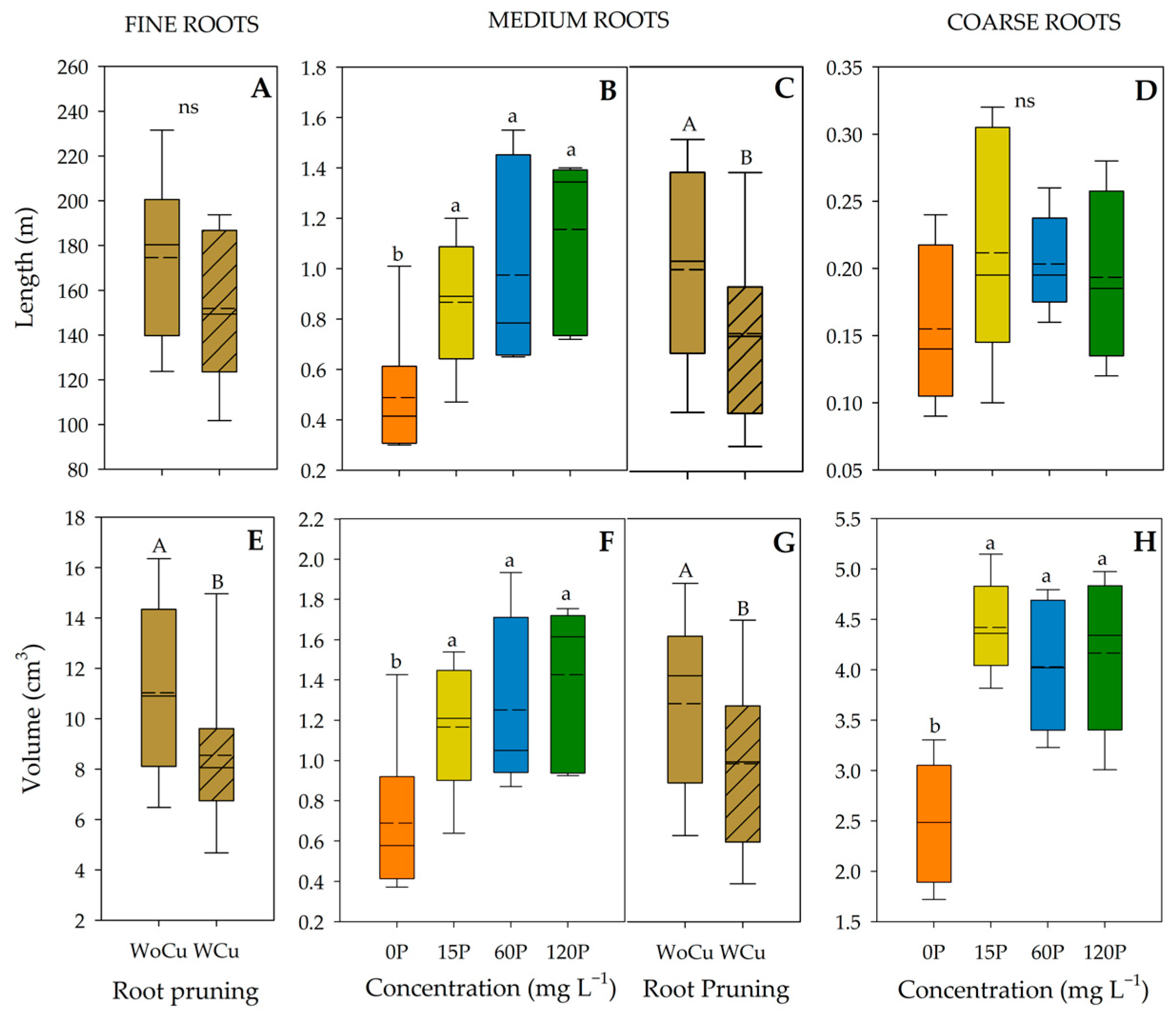
2.3. Root Architecture Traits
3. Discussion
4. Materials and Methods
4.1. Seedling Establishment in the Nursery
4.2. Fertilization
4.3. Morpho-Physiological Evaluations
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez, D.; Chadwick, C.; Plaza-Aguilar, A. The Time of Emergence (ToE) of the Andean Mediterranean Sclerophyllous Forest of Quillaja saponaria (Mol.) and Lithraea caustica (Mol.) Hox. & Arn. For. Ecol. Manag. 2023, 544, 121169. [Google Scholar] [CrossRef]
- Schröter, D.; Cramer, W.; Leemans, R.; Prentice, I.C.; Araújo, M.B.; Arnell, N.W.; Bondeau, A.; Bugmann, H.; Carter, T.R.; Gracia, C.A.; et al. Ecosystem Service Supply and Vulnerability to Global Change in Europe. Science 2005, 310, 1333–1337. [Google Scholar] [CrossRef] [PubMed]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterranean Biomes: Evolution of Their Vegetation, Floras, and Climate. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 383–407. [Google Scholar] [CrossRef]
- Miranda, A.; Syphard, A.D.; Berdugo, M.; Carrasco, J.; Gómez-González, S.; Ovalle, J.F.; Delpiano, C.A.; Vargas, S.; Squeo, F.A.; Miranda, M.D.; et al. Widespread Synchronous Decline of Mediterranean-Type Forest Driven by Accelerated Aridity. Nat. Plants 2023, 9, 1810–1817. [Google Scholar] [CrossRef]
- Rojas, M.; Lambert, F.; Ramirez-Villegas, J.; Challinor, A.J. Emergence of Robust Precipitation Changes across Crop Production Areas in the 21st Century. Proc. Natl. Acad. Sci. USA 2019, 116, 6673–6678. [Google Scholar] [CrossRef] [PubMed]
- Polade, S.D.; Gershunov, A.; Cayan, D.R.; Dettinger, M.D.; Pierce, D.W. Precipitation in a Warming World: Assessing Projected Hydro-Climate Changes in California and Other Mediterranean Climate Regions. Sci. Rep. 2017, 7, 10783. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Sardans, J. Global Change and Forest Disturbances in the Mediterranean Basin: Breakthroughs, Knowledge Gaps, and Recommendations. Forests 2021, 12, 603. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin III, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monog. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Picard, N.; Garavaglia, V. Mediterranean Forests and the United Nations Sustainable Development Goals. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Osem, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 603–616. [Google Scholar] [CrossRef]
- Leon-Lobos, P.; Bustamante-Sanchez, M.A.; Nelson, C.R.; Alarcon, D.; Hasbun, R.; Way, M.; Pritchard, H.W.; Armesto, J.J. Lack of Adequate Seed Supply Is a Major Bottleneck for Effective Ecosystem Restoration in Chile: Friendly Amendment to Bannister et al. (2018). Restor. Ecol. 2020, 28, 277–281. [Google Scholar] [CrossRef]
- Acevedo, M.; Álvarez-Maldini, C.; Dumroese, R.K.; Bannister, J.R.; Cartes, E.; González, M. Native Plant Production in Chile. Is It Possible to Achieve Restoration Goals by 2035? Land 2021, 10, 71. [Google Scholar] [CrossRef]
- Bannister, J.; Vargas-Gaete, R.; Ovalle, J.; Acevedo, M.; Fuentes-Ramirez, A.; Donoso, P.; Promis, A.; Smith-Ramirez, C. Major Bottlenecks for the Restoration of Natural Forests in Chile. Restor. Ecol. 2018, 26, 1039–1044. [Google Scholar] [CrossRef]
- Magni, C.R.; Poch, P.L.; Espinoza, S.E.; Yáñez, M.A.; Martínez, E.E.; Promis, A.A.; Mancilla, G.A. Provenance Influences Seed Germination and Phenotypic Responses to Water Restriction in the Endemic Beilschmiedia miersii (Gay) Kosterm. Front. For. Glob. Chang. 2023, 5, 1039908. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Negri, G.; Scippa, G.S.; Chiatante, D.; Terzaghi, M. Asymmetrical Copper Root Pruning May Improve Root Traits for Reforesting Steep and/or Windy Sites. New For. 2022, 53, 1093–1112. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Pinto, J.R.; Fulgaro, N.; Scippa, G.S.; Chiatante, D. Tree Seedling Response to LED Spectra: Implications for Forest Restoration. Plant Biosyst. 2018, 152, 515–523. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Landis, T.D.; Pinto, J.R.; Haase, D.L.; Wilkinson, K.W.; Davis, A.S. Meeting Forest Restoration Challenges: Using the Target Plant Concept. REFOR 2016, 37–52. [Google Scholar] [CrossRef]
- Cole, R.J.; Holl, K.D.; Keene, C.L.; Zahawi, R.A. Direct Seeding of Late-Successional Trees to Restore Tropical Montane Forest. For. Ecol. Manag. 2011, 261, 1590–1597. [Google Scholar] [CrossRef]
- Landis, T.D.; Dumroese, R.K.; Haase, D.L. The Target Plant Concept. In Container Tree Nursery Manual, Volume 7: Seedling Processing, Storage, and Outplanting; Agricultural Handbook 674; USDA Forest Service: Washington, DC, USA, 2010; pp. 3–15. [Google Scholar]
- Landis, T.D.; Dumroese, R.K. Applying the Target Plant Concept to Nursery Stock Quality. In Plant Quality: A Key to Success in Forest Establishment. Proceeding of the COFORD Conference. Tullow, Co Carlow, Ireland, 20–21 September 2005; MacLennan, L., Fennessy, J., Eds.; National Council for Forest Research and Development: Dublin, Ireland, 2006; pp. 1–10. Available online: https://www.researchgate.net/publication/239932357_Applying_the_target_plant_concept_to_nursery_stock_quality (accessed on 1 September 2024).
- Wang, F.-X.; Wang, Z.-Y.; Lee, J.H.W. Acceleration of Vegetation Succession on Eroded Land by Reforestation in a Subtropical Zone. Ecol. Eng. 2007, 31, 232–241. [Google Scholar] [CrossRef]
- Luoranen, J.; Rikala, R.; Konttinen, K.; Smolander, H. Summer Planting of Picea abies Container-Grown Seedlings: Effects of Planting Date on Survival, Height Growth and Root Egress. For. Ecol. Manag. 2006, 237, 534–544. [Google Scholar] [CrossRef]
- Hostetler, A.N.; Morais De Sousa Tinoco, S.; Sparks, E.E. Root Responses to Abiotic Stress: A Comparative Look at Root System Architecture in Maize and Sorghum. J. Exp. Bot. 2024, 75, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Villar-Salvador, P.; Puértolas, J.; Cuesta, B.; Peñuelas, J.L.; Uscola, M.; Heredia-Guerrero, N.; Rey Benayas, J.M. Increase in Size and Nitrogen Concentration Enhances Seedling Survival in Mediterranean Plantations. Insights from an Ecophysiological Conceptual Model of Plant Survival. New For. 2012, 43, 755–770. [Google Scholar] [CrossRef]
- Ovalle, J.F.; Ginocchio, R.; Arellano, E.C.; Valenzuela, P. Root Adaptive Management for Improving Plant Quality and Field Performance under Drought: Experiences with Native Tree Species from a South American Mediterranean-Type Ecosystem. Plant Sociol. 2017, 54, 19–27. [Google Scholar] [CrossRef]
- Cuesta, B.; Villar-Salvador, P.; Puértolas, J.; Jacobs, D.F.; Rey Benayas, J.M. Why Do Large, Nitrogen Rich Seedlings Better Resist Stressful Transplanting Conditions? A Physiological Analysis in Two Functionally Contrasting Mediterranean Forest Species. For. Ecol. Manag. 2010, 260, 71–78. [Google Scholar] [CrossRef]
- Luis, V.C.; Puértolas, J.; Climent, J.; Peters, J.; González-Rodríguez, Á.M.; Morales, D.; Jiménez, M.S. Nursery Fertilization Enhances Survival and Physiological Status in Canary Island Pine (Pinus canariensis) Seedlings Planted in a Semiarid Environment. Eur. J. For. Res. 2009, 128, 221–229. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; López-Arredondo, D. Phosphorus: The Underrated Element for Feeding the World. Trends Plant Sci. 2016, 21, 461–463. [Google Scholar] [CrossRef]
- Landis, T.D.; van Steenis, E. Macronutrients—Phosphorus. In Forest Nursery Notes, Summer 2004; USDA Forest Service, Pacific Northwest Region, State and Private Forestry Cooperative Programs: Portland, OR, USA, 2004; R6-CP-TP-07-04; pp. 6–14. Available online: https://rngr.net/publications/fnn/2004-summer/articles/macronutrients-phosphorus (accessed on 12 October 2024).
- Wu, C.; Wei, X.; Sun, H.; Wang, Z. Phosphate Availability Alters Lateral Root Anatomy and Root Architecture of Fraxinus Mandshurica Rupr. Seedlings. J. Integr. Plant Biol. 2005, 47, 292–301. [Google Scholar] [CrossRef]
- Song, C.J.; Ma, K.M.; Qu, L.Y.; Liu, Y.; Xu, X.L.; Fu, B.J.; Zhong, J.F. Interactive Effects of Water, Nitrogen and Phosphorus on the Growth, Biomass Partitioning and Water-Use Efficiency of Bauhinia faberi Seedlings. J. Arid Environ. 2010, 74, 1003–1012. [Google Scholar] [CrossRef]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Root Architecture and Hydraulic Conductance in Nutrient Deprived Pistacia lentiscus L. Seedlings. Oecologia 2012, 170, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J.; Rodà, F. Plasticity of Leaf Morphological Traits, Leaf Nutrient Content, and Water Capture in the Mediterranean Evergreen Oak Quercus ilex Subsp. Ballota in Response to Fertilization and Changes in Competitive Conditions. Ecoscience 2006, 13, 258–270. [Google Scholar] [CrossRef]
- Oliet, J.A.; Planelles, R.; Artero, F.; Jacobs, D.F. Nursery Fertilization and Tree Shelters Affect Long-Term Field Response of Acacia salicina Lindl. Planted in Mediterranean Semiarid Conditions. For. Ecol. Manag. 2005, 215, 339–351. [Google Scholar] [CrossRef]
- Baesso, B.; Chiatante, D.; Terzaghi, M.; Zenga, D.; Nieminen, K.; Mahonen, A.P.; Siligato, R.; Helariutta, Y.; Scippa, G.S.; Montagnoli, A. Transcription Factors PRE 3 and WOX 11 Are Involved in the Formation of New Lateral Roots from Secondary Growth Taproot in A. thaliana. Plant Biol. J. 2018, 20, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Miao, J.; Yumoto, E.; Yokota, T.; Asahina, M.; Watahiki, M. YUCCA9-Mediated Auxin Biosynthesis and Polar Auxin Transport Synergistically Regulate Regeneration of Root Systems Following Root Cutting. Plant Cell Physiol. 2017, 58, 1710–1723. [Google Scholar] [CrossRef] [PubMed]
- Wenny, D.L.; Liu, Y.; Dumroese, R.K.; Osborne, H.L. First Year Field Growth of Chemically Root Pruned Containerized Seedlings. New For. 1988, 2, 111–118. [Google Scholar] [CrossRef]
- Ruehle, J.L. The Effect of Cupric Carbonate on Root Morphology of Containerized Mycorrhizal Pine Seedlings. Can. J. For. Res. 1985, 15, 586–592. [Google Scholar] [CrossRef]
- Tsakaldimi, M.N.; Ganatsas, P.P. Effect of Chemical Root Pruning on Stem Growth, Root Morphology and Field Performance of the Mediterranean Pine Pinus halepensis Mill. Sci. Hortic. 2006, 109, 183–189. [Google Scholar] [CrossRef]
- Aguilera-Rodríguez, M.; Aldrete, A.; Vargas-Hernández, J.J.; López-Upton, J.; López-López, M.Á.; Ordaz-Chaparro, V.M. Morphology and Root Growth Potential of Pinus patula Produced in Trays with Root Pruning. Agrociencia 2021, 55, 81–97. [Google Scholar]
- Dumroese, R.K.; Sung, S.-J.S.; Pinto, J.R.; Ross-Davis, A.; Scott, D.A. Morphology, Gas Exchange, and Chlorophyll Content of Longleaf Pine Seedlings in Response to Rooting Volume, Copper Root Pruning, and Nitrogen Supply in a Container Nursery. New For. 2013, 44, 881–897. [Google Scholar] [CrossRef]
- Burdett, A.N. Physiological Processes in Plantation Establishment and the Development of Specifications for Forest Planting Stock. Can. J. For. Res. 1990, 20, 415–427. [Google Scholar] [CrossRef]
- Liu, J.; Bloomberg, M.; Li, G.; Liu, Y. Effects of Copper Root Pruning and Radicle Pruning on First-Season Field Growth and Nutrient Status of Chinese Cork Oak Seedlings. New For. 2016, 47, 715–729. [Google Scholar] [CrossRef]
- Guerrero, P.C.; Bustamante, R.O. Abiotic Alterations Caused by Forest Fragmentation Affect Tree Regeneration: A Shade and Drought Tolerance Gradient in the Remnants of Coastal Maulino Forest. Rev. Chil. Hist. Nat. 2009, 82, 413–429. [Google Scholar] [CrossRef]
- Fernández, M.P.; Preller, C.; Fischer, S.; Espinoza, C.; Peña-Rojas, K.; Menéndez-Miguélez, M. Maqui (Aristotelia chilensis [Molina] Stuntz): The Most Antioxidant Wild Berry towards Agricultural Production. Fruits 2019, 74, 214–226. [Google Scholar] [CrossRef]
- Magni, C.; Espinoza, S.; Poch, P.; Abarca, B.; Grez, I.; Martínez, E.; Yáñez, M.; Santelices, R.; Cabrera, A. Growth and Biomass Partitioning of Nine Provenances of Quillaja saponaria Seedlings to Water Stress. South. For. 2019, 81, 103–109. [Google Scholar] [CrossRef]
- Pelah, D.; Abramovich, Z.; Markus, A.; Wiesman, Z. The Use of Commercial Saponin from Quillaja saponaria Bark as a Natural Larvicidal Agent against Aedes aegypti and Culex pipiens. J. Ethnopharmacol. 2002, 81, 407–409. [Google Scholar] [CrossRef]
- López, M.; Abarca, B.; Espinoza, S.; Rojas, A.; Martínez-Herrera, E.; Yáñez, M.; Magni, C.R. A Proposed Methodology for the Determination of Seed Sources for Tree Native Species Based on Environmental Variables: The Case of Quillaja saponaria Mol. New For. 2024, 55, 1–13. [Google Scholar] [CrossRef]
- Razaq, M.; Zhang, P.; Shen, H.-L.; Salahuddin. Influence of Nitrogen and Phosphorous on the Growth and Root Morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [PubMed]
- Andivia, E.; Fernández, M.; Vázquez-Piqué, J. Autumn Fertilization of Quercus ilex ssp. Ballota (Desf.) Samp. Nursery Seedlings: Effects on Morpho-Physiology and Field Performance. Ann. For. Sci. 2011, 68, 543–553. [Google Scholar] [CrossRef]
- Constantino, V.; Motta, A.C.V.; Barbosa, J.Z.; Dolinski, M.A.; Zanette, F.; Prior, S.A. Initial Growth of Araucaria angustifolia Rootstock in Response to Fertilization with Nitrogen, Phosphorus and Potassium. RF 2018, 49, 099. [Google Scholar] [CrossRef]
- Grossnickle, S. Importance of Root Growth in Overcoming Planting Stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Grossnickle, S. Seedling Size and Reforestation Success. How Big Is Big Enough? In The Thin Green Line: A Symposium on the State-of-the-Art in Reforestation, Thunder Bay, ON, Canada, 26–28 Jul 2005; Colombo, S.J. (Compiler) Ontario Ministry of Natural Resources: Sault Ste. Marie, ON, Canada, 2005; Forest Research Information Paper 160; pp. 138–143. Available online: https://rngr.net/publications/tgl/seedling-size-and-reforestation-success-how-big-is-big-enough (accessed on 1 October 2024).
- Qu, L.; Quoreshi, A.M.; Koike, T. Root Growth Characteristics, Biomass and Nutrient Dynamics of Seedlings of Two Larch Species Raised under Different Fertilization Regimes. Plant Soil 2003, 255, 293–302. [Google Scholar] [CrossRef]
- Gleeson, S.K.; Good, R.E. Root Allocation and Multiple Nutrient Limitation in the New Jersey Pinelands. Ecol. Lett. 2003, 6, 220–227. [Google Scholar] [CrossRef]
- Singh, D.K.; Sale, P.W.G.; Pallaghy, C.K.; Mckenzie, B.M. Phosphorus Concentrations in the Leaves of Defoliated White Clover Affect Abscisic Acid Formation and Transpiration in Drying Soil. New Phytol. 2000, 146, 249–259. [Google Scholar] [CrossRef]
- Marchioretto, L.D.R.; De Rossi, A.; Conte, E.D. Chemical Root Pruning Improves Quality and Nutrient Uptake of Cape Gooseberry (Physalis peruviana) Seedlings. Sci. Hortic. 2020, 261, 108948. [Google Scholar] [CrossRef]
- Graciano, C.; Goya, J.F.; Frangi, J.L.; Guiamet, J.J. Fertilization with Phosphorus Increases Soil Nitrogen Absorption in Young Plants of Eucalyptus grandis. Forest Ecol. Manag. 2006, 236, 202–210. [Google Scholar] [CrossRef]
- Seabra, C.E.B.C.; Osiecka, A.; Tucci, C.A.F.; Minogue, P.J.; Pereira, B.F.F.; Andersen, P.C. Influence of Phosphorus Limitations on the Growth, Nutrient Partitioning and Physiology of Mahogany (Swietenia macrophylla King) Seedlings. J. Plant Nutr. 2018, 41, 358–370. [Google Scholar] [CrossRef]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Plant Morphology and Root Hydraulics Are Altered by Nutrient Deficiency in Pistacia lentiscus (L.). Trees 2006, 20, 334–339. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Sun, X.; Song, D.; Chen, W.; Zhang, A.; et al. Phosphorous Application Improves Drought Tolerance of Phoebe zhennan. Front. Plant Sci. 2017, 8, 1561. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.; Faustino, L.I.; Graciano, C. The Spatial Distribution of Phosphate in the Root System Modulates N Metabolism and Growth in Eucalyptus grandis Young Plants. Trees 2017, 31, 247–257. [Google Scholar] [CrossRef]
- Tariq, A.; Graciano, C.; Pan, K.; Olatunji, O.A.; Li, Z.; Sadia, S.; Zhang, Z.; Ismoilov, K.; Ahmed, Z.; Ullah, A.; et al. Phosphorus Fertilization of Phoebe zhennan Seedlings under Drought Reduces Nitrogen Assimilation. J. Plant Nutr. 2022, 45, 2228–2238. [Google Scholar] [CrossRef]
- Heydari, M.M.; Brook, R.M.; Jones, D.L. The Role of Phosphorus Sources on Root Diameter, Root Length and Root Dry Matter of Barley (Hordeum vulgare L.). J. Plant Nutr. 2019, 42, 1–15. [Google Scholar] [CrossRef]
- Fernandes, A.M.; Soratto, R.P.; Gonsales, J.R. Root Morphology and Phosphorus Uptake by Potato Cultivars Grown under Deficient and Sufficient Phosphorus Supply. Sci. Hortic. 2014, 180, 190–198. [Google Scholar] [CrossRef]
- Herdler, S.; Kreuzer, K.; Scheu, S.; Bonkowski, M. Interactions between Arbuscular Mycorrhizal Fungi (Glomus intraradices, Glomeromycota) and Amoebae (Acanthamoeba castellanii, Protozoa) in the Rhizosphere of Rice (Oryza sativa). Soil Biol. Biochem. 2008, 40, 660–668. [Google Scholar] [CrossRef]
- Jin, J.; Wang, G.; Liu, X.; Pan, X.; Herbert, S.J. Phosphorus Application Affects the Soybean Root Response to Water Deficit at the Initial Flowering and Full Pod Stages. Soil Sci. Plant Nutr. 2005, 51, 953–960. [Google Scholar] [CrossRef]
- Toro, M.; Azcon, R.; Barea, J. Improvement of Arbuscular Mycorrhiza Development by Inoculation of Soil with Phosphate-Solubilizing Rhizobacteria To Improve Rock Phosphate Bioavailability ((Sup32)P) and Nutrient Cycling. Appl. Environ. Microbiol. 1997, 63, 4408–4412. [Google Scholar] [CrossRef]
- Huang, B.; Nobel, P.S. Hydraulic Conductivity and Anatomy for Lateral Roots of Agave Deserti During Root Growth and Drought-Induced Abscission. J. Exp. Bot. 1992, 43, 1441–1449. [Google Scholar] [CrossRef]
- Fitter, A.H.; Stickland, T.R.; Harvey, M.L.; Wilson, G.W. Architectural Analysis of Plant Root Systems 1. Architectural Correlates of Exploitation Efficiency. New Phytol. 1991, 118, 375–382. [Google Scholar] [CrossRef]
- Toca, A.; Moler, E.; Nelson, A.; Jacobs, D.F. Environmental Conditions in the Nursery Regulate Root System Development and Architecture of Forest Tree Seedlings: A Systematic Review. New For. 2022, 53, 1113–1143. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root Traits are Multidimensional: Specific Root Length Is Independent from Root Tissue Density and the Plant Economic Spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Laliberté, E.; Lambers, H.; Burgess, T.I.; Wright, J. Phosphorus Limitation, Soil-Borne Pathogens and the Coexistence of Plant Species in Hyperdiverse Forests and Shrublands. New Phytol. 2015, 206, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.D.; Rosas, J.C.; Brown, K.M.; Lynch, J.P. Root Architectural Tradeoffs for Water and Phosphorus Acquisition. Funct. Plant Biol. 2005, 32, 737. [Google Scholar] [CrossRef]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific Root Length as an Indicator of Environmental Change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- Xu, B.; Niu, F.; Duan, D.; Xu, W.-Z.; Huang, J. Root Morphological Characteristics of Lespedeza davurica (L.) Intercropped with Bothriochloa ischaemum (L.) Keng under Water Stress and P Application Conditions. Pak. J. Bot. 2012, 44, 1857–1864. [Google Scholar]
- Ji, L.; Attaullah, K.; Wang, J.; Yu, D.; Yang, Y.; Yang, L.; Lu, Z. Root Traits Determine Variation in Nonstructural Carbohydrates (NSCs) under Different Drought Intensities and Soil Substrates in Three Temperate Tree Species. Forests 2020, 11, 415. [Google Scholar] [CrossRef]
- Padilla, F.M.; Ortega, R.; Sánchez, J.; Pugnaire, F.I. Rethinking Species Selection for Restoration of Arid Shrublands. Basic Appl. Ecol. 2009, 10, 640–647. [Google Scholar] [CrossRef]
- Wright, I.J.; Westoby, M. Differences in Seedling Growth Behaviour among Species: Trait Correlations across Species, and Trait Shifts along Nutrient Compared to Rainfall Gradients. J. Ecol. 1999, 87, 85–97. [Google Scholar] [CrossRef]
- Birouste, M.; Zamora-Ledezma, E.; Bossard, C.; Pérez-Ramos, I.M.; Roumet, C. Measurement of Fine Root Tissue Density: A Comparison of Three Methods Reveals the Potential of Root Dry Matter Content. Plant Soil 2014, 374, 299–313. [Google Scholar] [CrossRef]
- Rewald, B.; Rechenmacher, A.; Godbold, D.L. It’s Complicated: Intraroot System Variability of Respiration and Morphological Traits in Four Deciduous Tree Species. Plant Physiol. 2014, 166, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Olmo, M.; Lopez-Iglesias, B.; Villar, R. Drought Changes the Structure and Elemental Composition of Very Fine Roots in Seedlings of Ten Woody Tree Species. Implications for a Drier Climate. Plant Soil 2014, 384, 113–129. [Google Scholar] [CrossRef]
- King, J.S.; Albaugh, T.J.; Allen, H.L.; Buford, M.; Strain, B.R.; Dougherty, P. Below-Ground Carbon Input to Soil Is Controlled by Nutrient Availability and Fine Root Dynamics in Loblolly Pine. New Phytol. 2002, 154, 389–398. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How Tree Roots Respond to Drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef]
- Acevedo, M.; Álvarez, C.; Cartes, E.; Dumroese, R.K.; González, M. Production and Establishment Techniques for the Restoration of Nothofagus alessandrii, an Endangered Keystone Species in a Mediterranean Forest. New For. 2020, 51, 159–174. [Google Scholar] [CrossRef]
- González, M.; Ríos, D.; Peña Rojas, K.; García, E.; Acevedo, M.; Cartes, E.; Sánchez Olate, M. Efecto de la concentración de fósforo y calcio sobre atributos morfo-fisiológicos y potencial de crecimiento radical en plantas de Aextoxicon punctatum producidas a raíz cubierta en la etapa de endurecimiento. Bosque 2020, 41, 137–146. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Montville, M.E.; Pinto, J.R. Using Container Weights to Determine Irrigation Needs: A Simple Method. Native Plants J. 2015, 16, 67–71. [Google Scholar] [CrossRef]
- Alvarez-Maldini, C.; Acevedo, M.; Dumroese, R.K.; González, M.; Cartes, E. Intraspecific Variation in Drought Response of Three Populations of Cryptocarya alba and Persea lingue, Two Native Species from Mediterranean Central Chile. Front. Plant Sci. 2020, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Galmés, J.; Cifre, J.; Medrano, H.; Flexas, J. Modulation of Relative Growth Rate and Its Components by Water Stress in Mediterranean Species with Different Growth Forms. Oecologia 2005, 145, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Bergerud, W.A. Introduction to Logistic Regression Models with Worked Forestry Examples: Biometrics Information Handbook No.7; Ministry of Forests Research Program: Victoria, BC, Canada, 1996.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Aristotelia chilensis | Quillaja saponaria | |||||
|---|---|---|---|---|---|---|---|
| Phosphorous (P) | Root Pruning (RP) | P × RP | Phosphorous (P) | Root Pruning (RP) | P × RP | ||
| Morphological | Leaf area | ˂0.0001 | 0.2788 | 0.9136 | ˂0.0001 | 0.0057 | 0.4347 |
| Leaf biomass | ˂0.0001 | 0.1892 | 0.8667 | ˂0.0001 | 0.0029 | 0.3596 | |
| Stem biomass | ˂0.0001 | 0.0045 | 0.0006 | ˂0.0001 | 0.0004 | 0.4996 | |
| Root biomass | 0.0404 | 0.0086 | 0.3849 | ˂0.0001 | 0.1606 | 0.5951 | |
| R:S | ˂0.0001 | 0.0041 | 0.8804 | 0.0522 | 0.0228 | 0.8139 | |
| Nutritional | N concentration | ˂0.0001 | 0.0006 | 0.1396 | ˂0.0001 | 0.0001 | 0.0131 |
| P concentration | ˂0.0001 | 0.0124 | 0.1630 | 0.0016 | 0.0012 | 0.0074 | |
| K concentration | ˂0.0001 | 0.1950 | 0.8049 | 0.0043 | 0.9467 | 0.1298 | |
| N content | ˂0.0001 | 0.1172 | 0.8296 | ˂0.0001 | 0.3275 | 0.8368 | |
| P content | ˂0.0001 | 0.5542 | 0.9462 | ˂0.0001 | 0.2370 | 0.7441 | |
| K content | 0.0552 | 0.8510 | 0.7362 | ˂0.0001 | 0.0249 | 0.4022 | |
| Root architecture | Root length | 0.2742 | 0.2417 | 0.4801 | ˂0.0001 | 0.1532 | 0.1353 |
| Root diameter | 0.1095 | 0.1468 | 0.5625 | ˂0.0001 | 0.2709 | 0.3179 | |
| Root volume | 0.0785 | 0.0364 | 0.4313 | ˂0.0001 | 0.8007 | 0.0099 | |
| Root surface area | 0.1601 | 0.0616 | 0.4043 | ˂0.0001 | 0.4938 | 0.0140 | |
| SRL | 0.4362 | 0.1305 | 0.5325 | 0.0002 | 0.0350 | 0.0862 | |
| RTD | 0.1140 | 0.8988 | 0.3191 | 0.1142 | 0.0001 | 0.0263 | |
| Biomass (g) | |||||||
|---|---|---|---|---|---|---|---|
| Leaf Area (cm2) | Leaf | Stem | Root | R:S | |||
| WoCu | WCu | ||||||
| A. chilensis | Phosphorous (P) | ||||||
| 0P | 383.15 ± 102.21 b | 4.03 ± 0.45 b | 5.63 ± 0.61 d | 5.05 ± 0.53 d | 3.48 ± 1.87 b | 0.36 ± 0.08 a | |
| 15P | 759.53 ± 217.65 a | 7.66 ± 1.52 a | 12.3 ± 0.10 c | 16.00 ± 2.62 ab | 4.28 ± 2.27 ab | 0.20 ± 0.05 b | |
| 60P | 781.03 ± 218.31 a | 7.75 ± 1.28 a | 14.62 ± 1.28 abc | 14.98 ± 2.53 abc | 3.85 ± 2.05 ab | 0.18 ± 0.03 b | |
| 120P | 803.55 ± 157.65 a | 8.26 ± 1.15 a | 14.35 ± 2.39 bc | 18.08 ± 1.81 a | 4.40 ± 2.32 a | 0.19 ± 0.04 b | |
| Root Pruning (RP) | |||||||
| WoCu | 667.37 ± 237.28 ns | 7.07 ± 1.98 ns | 11.72 ± 4.04 ns | 4.33 ± 2.25 a | 0.26 ± 0.10 a | ||
| WCu | 700.79 ± 260.75 ns | 6.78 ± 2.13 ns | 13.53 ± 5.46 ns | 3.67 ± 1.95 b | 0.20 ± 0.08 b | ||
| Q. saponaria | Phosphorous (P) | ||||||
| 0P | 13.37 ± 6.17 c | 0.21 ± 0.09 c | 0.06 ± 0.05 c | 0.14 ± 0.08 c | 0.65 ± 0.21 ns | ||
| 15P | 44.55 ± 19.47 b | 0.72 ± 0.36 b | 0.25 ± 0.15 b | 0.69 ± 0.44 b | 0.75 ± 0.15 ns | ||
| 60P | 87.95 ± 18.73 a | 1.49 ± 0.42 a | 0.52 ± 0.13 a | 1.21 ± 0.68 a | 0.61 ± 0.07 ns | ||
| 120P | 109.69 ± 44.19 a | 1.81 ± 0.92 a | 0.71 ± 0.43 a | 1.31 ± 0.76 a | 0.56 ± 0.07 ns | ||
| Root Pruning (RP) | |||||||
| WoCu | 55.84 ± 39.26 b | 0.88 ± 0.66 b | 0.30 ± 0.23 b | 0.77 ± 0.56 ns | 0.70 ± 0.14 a | ||
| WCu | 71.94 ± 50.34 a | 1.23 ± 0.94 a | 0.47 ± 0.41 a | 0.90 ± 0.63 ns | 0.59 ± 0.14 b | ||
| Nutrient Concentration (mg g−1) | Nutrient Content (mg) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| A. chilensis | N | P | K | N | P | K | |||
| Phosphorous (P) | |||||||||
| 0P | 12.15 ± 1.76 b | 0.67 ± 0.12 d | 12.10 ± 2.30 a | 48.17 ± 4.49 b | 2.63 ± 0.22 c | 47.69 ± 4.34 ns | |||
| 15P | 16.47 ± 2.76 a | 1.28 ± 0.15 c | 8.28 ± 0.90 b | 125.22 ± 23.96 a | 9.81 ± 1.80 b | 63.77 ± 14.95 ns | |||
| 60P | 16.58 ± 1.01 a | 1.77 ± 0.12 b | 6.60 ± 0.50 c | 133.75 ± 17.85 a | 14.29 ± 2.39 a | 53.29 ± 7.81 ns | |||
| 120P | 17.60 ± 1.87 a | 2.08 ± 0.23 a | 7.43 ± 0.80 bc | 136.97 ± 14.32 a | 16.52 ± 2.09 a | 58.89 ± 6.43 ns | |||
| Root pruning (RP) | |||||||||
| WoCu | 14.47 ± 2.65 b | 1.51 ± 0.57 a | 8.24 ± 1.97 ns | 105.56 ± 39.07 ns | 10.62 ± 5.66 ns | 55.87 ± 12.91 ns | |||
| WCu | 16.78 ± 2.44 a | 1.39 ± 0.58 b | 8.97 ± 2.95 ns | 116.50 ± 42.79 ns | 11.01 ± 5.95 ns | 55.95 ± 8.31 ns | |||
| Q. saponaria | WoCu | WCu | WoCu | WCu | |||||
| Phosphorous (P) | |||||||||
| 0P | 34.43 ± 4.89 a | 27.33 ± 1.99 b | 2.10 ± 0.26 b | 2.06 ± 0.21 ab | 9.38 ± 1.21 b | 7.09 ± 1.82 c | 0.49 ± 0.16 c | 2.23 ± 0.82 c | |
| 15P | 20.67 ± 2.57 c | 18.27 ± 0.42 cd | 1.83 ± 0.25 a | 1.60 ± 0.17 c | 12.20 ± 1.25 a | 15.94 ± 3.88 b | 1.41 ± 0.40 b | 10.06 ± 2.61 b | |
| 60P | 15.90 ± 0.69 d | 16.20 ± 0.70 d | 1.80 ± 0.17 ab | 1.77 ± 0.06 bc | 10.75 ± 1.19 ab | 24.64 ± 3.92 a | 2.73 ± 0.35 a | 9.38 ± 1.81 b | |
| 120P | 21.07 ± 0.70 c | 15.47 ± 0.49 d | 2.43 ± 0.15 ab | 1.70 ± 0.17 bc | 11.12 ± 1.04 ab | 34.15 ± 8.84 a | 3.80 ± 0.76 a | 12.20 ± 6.83 a | |
| Root pruning (RP) | |||||||||
| WoCu | 23.02 ± 7.59 ns | 2.04 ± 0.32 ns | 10.93 ± 1.87 ns | 19.13 ± 9.78 ns | 2.00 ± 1.28 ns | 10.97 ± 6.50 b | |||
| WCu | 19.32 ± 5.04 ns | 1.78 ± 0.23 ns | 10.79 ± 1.09 ns | 21.78 ± 13.10 ns | 2.22 ± 1.48 ns | 13.97 ± 9.45 a | |||
| Source of Variation | Length | Volume | |||||
|---|---|---|---|---|---|---|---|
| Fine | Medium | Coarse | Fine | Medium | Coarse | ||
| A. chilensis | Phosphorous (P) | 0.2792 | 0.0021 | 0.2954 | 0.0675 | 0.0038 | 0.0004 |
| Root Pruning (RP) | 0.2472 | 0.0380 | 0.0807 | 0.0485 | 0.0382 | 0.9086 | |
| P × RP | 0.4802 | 0.5809 | 0.4547 | 0.5076 | 0.4450 | 0.8502 | |
| Q. saponaria | Phosphorous (P) | 0.0003 | 0.0008 | 0.0146 | 0.0001 | 0.0008 | 0.0035 |
| Root Pruning (RP) | 0.0322 | 0.0264 | 0.1082 | 0.0199 | 0.0428 | 0.0677 | |
| P × RP | 0.0011 | 0.0778 | 0.0899 | 0.0516 | 0.0379 | 0.0343 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderón-Ureña, F.; Álvarez-Maldini, C.; Acevedo, M.; Sánchez-Olate, M.E.; Dumroese, R.K.; Sierra-Olea, A.; Ovalle, J.F.; Esquivel-Segura, E. Phosphorus Fertilization and Chemical Root Pruning: Effects on Root Traits During the Nursery Stage in Two Mediterranean Species from Central Chile. Plants 2025, 14, 195. https://doi.org/10.3390/plants14020195
Calderón-Ureña F, Álvarez-Maldini C, Acevedo M, Sánchez-Olate ME, Dumroese RK, Sierra-Olea A, Ovalle JF, Esquivel-Segura E. Phosphorus Fertilization and Chemical Root Pruning: Effects on Root Traits During the Nursery Stage in Two Mediterranean Species from Central Chile. Plants. 2025; 14(2):195. https://doi.org/10.3390/plants14020195
Chicago/Turabian StyleCalderón-Ureña, Fiorella, Carolina Álvarez-Maldini, Manuel Acevedo, Manuel E. Sánchez-Olate, R. Kasten Dumroese, Antay Sierra-Olea, Juan F. Ovalle, and Edwin Esquivel-Segura. 2025. "Phosphorus Fertilization and Chemical Root Pruning: Effects on Root Traits During the Nursery Stage in Two Mediterranean Species from Central Chile" Plants 14, no. 2: 195. https://doi.org/10.3390/plants14020195
APA StyleCalderón-Ureña, F., Álvarez-Maldini, C., Acevedo, M., Sánchez-Olate, M. E., Dumroese, R. K., Sierra-Olea, A., Ovalle, J. F., & Esquivel-Segura, E. (2025). Phosphorus Fertilization and Chemical Root Pruning: Effects on Root Traits During the Nursery Stage in Two Mediterranean Species from Central Chile. Plants, 14(2), 195. https://doi.org/10.3390/plants14020195