
Nomenclature and Typification of the Goat Grass Aegilops tauschii Coss. (Poaceae: Triticeae): A Key Species for the Secondary Gene Pool of Common Wheat Triticum aestivum

 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Nomenclatural Background
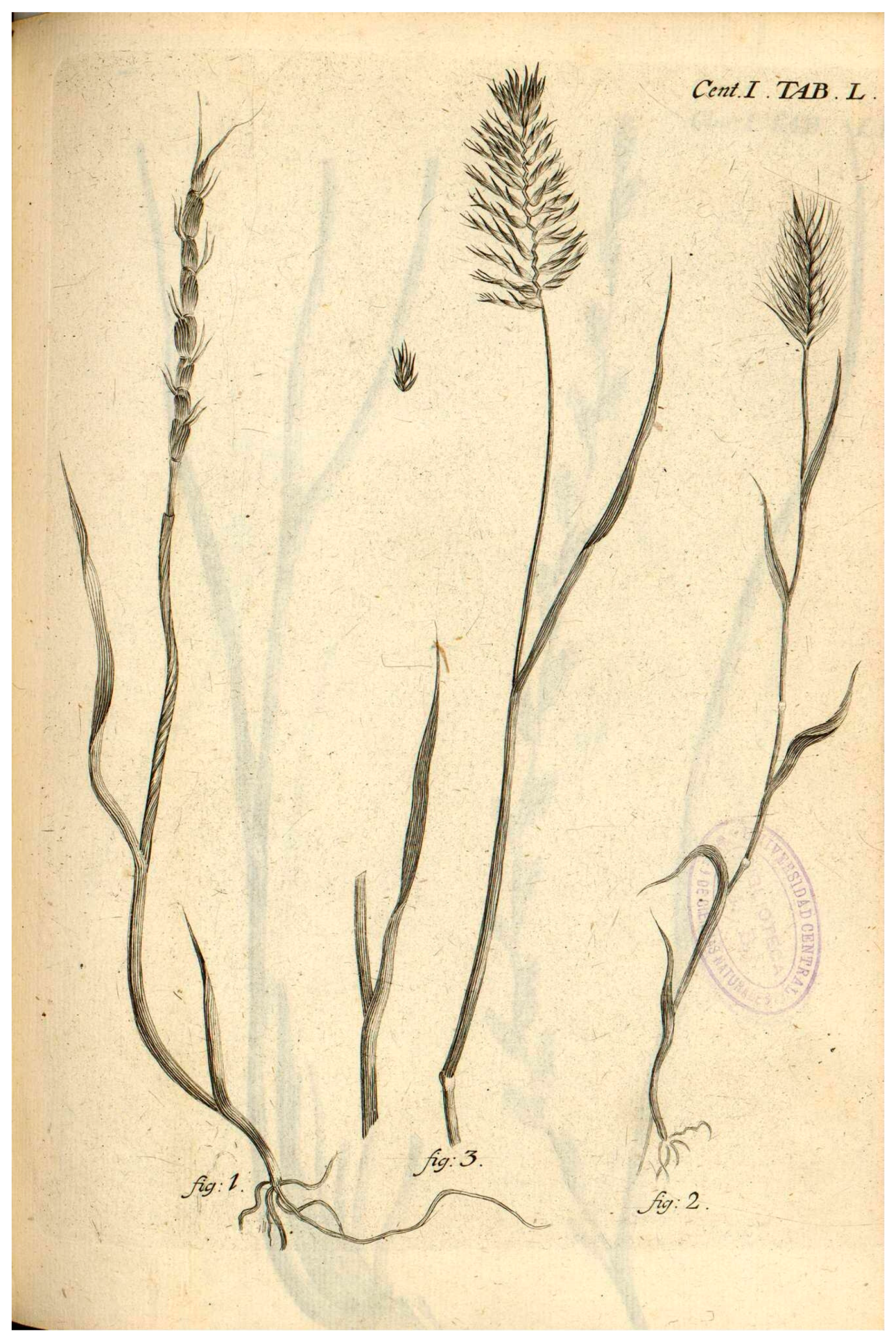
2.2. Typification of the Name
3. Materials and Methods
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, H.T.; Vales, M.I.; Mallory-Smith, C.; Riera-Lizarazu, O. Genetic Structure of Aegilops cylindrica Host in Its Native Range and in the United States of America. Theor. Appl. Genet. 2009, 119, 1013–1025. [Google Scholar] [CrossRef]
- Tzvelev, N.N. The System of the Grasses (Poaceae) and Their Evolution. Bot. Rev. 1989, 55, 193–194. [Google Scholar] [CrossRef]
- Molnár, I.; Vrána, J.; Burešová, V.; Cápal, P.; Farkas, A.; Darkó, É.; Cseh, A.; Kubaláková, M.; Molnár-Láng, M.; Doležel, J. Dissecting the U, M, S and C Genomes of Wild Relatives of Bread Wheat (Aegilops spp.) into Chromosomes and Exploring Their Synteny with Wheat. Plant J. 2016, 88, 452–467. [Google Scholar] [CrossRef]
- Zhao, L.; Ning, S.; Yi, Y.; Zhang, L.; Yuan, Z.; Wang, J.; Zheng, Y.; Hao, M.; Liu, D. Fluorescence in Situ Hybridization Karyotyping Reveals the Presence of Two Distinct Genomes in the Taxon Aegilops tauschii. BMC Genom. 2018, 19, 3. [Google Scholar] [CrossRef]
- Kiani, R.; Arzani, A.; Mirmohammady Maibody, S.A.M. Polyphenols, Flavonoids, and Antioxidant Activity Involved in Salt Tolerance in Wheat, Aegilops cylindrica and Their Amphidiploids. Front. Plant Sci. 2021, 12, 646221. [Google Scholar] [CrossRef]
- Zhao, Y.; Qi, G.; Ren, F.; Wang, Y.; Wang, P.; Wu, X. Analysis of PYL Genes and Their Potential Relevance to Stress Tolerance and Berry Ripening in Grape. J. Am. Soc. Hortic. Sci. 2020, 145, 308–317. [Google Scholar] [CrossRef]
- Wang, X.; Yoo, E.; Lee, S.; Cho, G.-T.; Lee, G.-A.; Yi, J.Y.; Du, X.; Han, S.; Hyun, D.Y.; Ro, N.; et al. Classification of 17 Species Aegilops Using DNA Barcoding and SNPs, Reveals Gene Flow among Aegilops biuncialis, Aegilops juvenalis, and Aegilops columnaris. Front. Plant Sci. 2022, 13, 984825. [Google Scholar] [CrossRef]
- Van Slageren, M.W. Wild Wheats: A Monograph of Aegilops L. and Amblyopyrum (Jaub. & Spach) Eig (Poaceae); Wageningen Agricultural University Papers 94–7; Wageningen Agricultural University: Wageningen, The Netherlands, 1994. [Google Scholar]
- POWO Plant of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:384652-1 (accessed on 22 January 2025).
- Zhukovsky, P.M. A Critical-Systematical Survey of the Species of the Genus Aegilops L. Bull. Appl. Bot. Genet. Plant Breed 1928, 18, 417–609. [Google Scholar]
- Eig, A. Monographisch-Kritische Übersicht Der Gattung Aegilops. Feddes Reper. 1929, 55, 1–228. [Google Scholar]
- Morrison, L.A. Reevaluation of Systematic Relationships in Triticum L. and Aegilops L. Based on Comparative Morphological and Anatomical Investigations of Dispersal Mechanisms. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1994. [Google Scholar]
- Gupta, P.K.; Baum, B.R. Stable Classification and Nomenclature in the Triticeae: Desirability, Limitations and Prospects. Euphytica 1989, 41, 191–197. [Google Scholar] [CrossRef]
- Jia, J.; Zhao, S.; Kong, X.; Li, Y.; Zhao, G.; He, W.; Appels, R.; Pfeifer, M.; Tao, Y.; Zhang, X.; et al. Aegilops tauschii Draft Genome Sequence Reveals a Gene Repertoire for Wheat Adaptation. Nature 2013, 496, 91–95. [Google Scholar] [CrossRef]
- Cavalet-Giorsa, E.; González-Muñoz, A.; Athiyannan, N.; Holden, S.; Salhi, A.; Gardener, C.; Quiroz-Chávez, J.; Rustamova, S.M.; Elkot, A.F.; Patpour, M.; et al. Origin and Evolution of the Bread Wheat D Genome. Nature 2024, 633, 848–855. [Google Scholar] [CrossRef]
- Tsunewaki, K. Plasmon Analysis in the Triticum-Aegilops Complex. Breed. Sci. 2009, 470, 455–470. [Google Scholar] [CrossRef]
- Kihara, H. Discovery of the DD-Analyser, One of the Ancestors of Triticum vulgare. Agric. Hort. 1944, 19, 13–14. [Google Scholar]
- Mcfadden, E.S.; Sears, E.R. The Origin of Triticum spelta and Its Free-Threshing Hexaploid Relatives. J. Hered. 1946, 37, 81–89. [Google Scholar] [CrossRef]
- Petersen, G.; Seberg, O.; Yde, M.; Berthelsen, K. Phylogenetic Relationships of Triticum and Aegilops and Evidence for the Origin of the A, B, and D Genomes of Common Wheat (Triticum aestivum). Mol. Phylogenet. Evol. 2006, 39, 70–82. [Google Scholar] [CrossRef]
- Wang, J.; Luo, M.-C.; Chen, Z.; You, F.M.; Wei, Y.; Zheng, Y.; Dvorak, J. Aegilops tauschii Single Nucleotide Polymorphisms Shed Light on the Origins of Wheat D-Genome Genetic Diversity and Pinpoint the Geographic Origin of Hexaploid Wheat. New Phytol. 2013, 198, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, M.S.; Rao, M.V. Macro-Mutations and Subspecific Differentiation in Triticum. Wheat Inf. Serv. 1961, 13, 9–11. [Google Scholar]
- Dvorak, J.; Luo, M.-C.; Yang, Z.-L.; Zhang, H.-B. The Structure of the Aegilops tauschii Genepool and the Evolution of Hexaploid Wheat. Theor. Appl. Genet. 1998, 97, 657–670. [Google Scholar] [CrossRef]
- Goncharov, N.P.; Golovnina, K.A.; Kondratenko, E.Y. Taxonomy and Molecular Phylogeny of Natural and Artificial Wheat Species. Breed. Sci. 2009, 498, 492–498. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of Polyploid Triticum Wheats under Cultivation: The Role of Domestication, Natural Hybridization and Allopolyploid Speciation in Their Diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, T.; Rodriguez, J.C.; Deal, K.R.; Dubcovsky, J.; McGuire, P.E.; Lux, T.; Spannagl, M.; Mayer, K.F.X.; Baldrich, P.; et al. Aegilops tauschii Genome Assembly Aet v5.0 Features Greater Sequence Contiguity and Improved Annotation. G3 2021, 11, jkab325. [Google Scholar] [CrossRef]
- Payne, P.I. Genetics of Wheat Storage Proteins and the Effect of Allelic Variation on Bread-Making Quality. Annu. Rev. Plant Biol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Chepurnov, G.Y.; Ovchinnikova, E.S.; Blinov, A.G.; Chikida, N.N.; Belousova, M.K.; Goncharov, N.P. Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss. Plants 2023, 12, 3596. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Raupp, W.J.; Sharma, H.C.; Browder, L.E.; Hatchett, J.H.; Cook, D.L.S.; Hetz, J.H.W.L.; Anderson, M.M. Resistance in Aegilops squarrosa to Wheat Leaf Rust, Wheat Powdery Mildew, Greenbug, and Hessian Fly. Plant Dis. 1986, 70, 553–556. [Google Scholar] [CrossRef]
- Zohary, D.; Feldman, M. Hybridization between Amphidiploids and the Evolution of Polyploids in the Wheat (Aegilops-Triticum) Group. Evolution 1962, 16, 44–61. [Google Scholar] [CrossRef]
- Goncharov, N.P. Genus Triticum L. Taxonomy: The Present and the Future. Plant Syst. Evol. 2011, 295, 1–11. [Google Scholar] [CrossRef]
- Goncharov, N.P. Comparative Genetics of Wheats and Their Related Species; Publishing House of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2002. (In Russian) [Google Scholar]
- Goncharov, N.P. Opredelitel’ Raznovidnostey Myagkoy I Tverdoy Pshenits; Manual Book of Common and Hard Wheat Varieties; Publishing House of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2009. (In Russian) [Google Scholar]
- MacKey, J. Wheat: Its Concept, Evolution, and Taxonomy, 1st ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Kislev, M.E. Triticum parvicoccum sp. Nov., the Oldest Naked Wheat. Isr. J. Bot. 1979, 28, 95–107. [Google Scholar] [CrossRef]
- Udachin, R.A. On the Possible Current Existence of Triticum antiquorum Heer. Nauch.-Tekhn. Byul. 1982, 119, 72–73. [Google Scholar]
- Udachin, R.A.N.I. Vavilov and Studies of Wheats from Central Asia. Tr. Prikl. Bot. Genet. Sel. 1991, 140, 47–57. [Google Scholar]
- Goncharov, N.P.; Gaidalenok, R.F. Localization of Genes Controlling Spherical Grain and Compact Ear in Triticum antiquorum Heer Ex Udacz. Russ. J. Genet. 2005, 41, 1262–1267. [Google Scholar] [CrossRef]
- Rivera, D.; Ferrer-Gallego, P.P.; Obón, C.; Alcaraz, F.; Laguna, E.; Goncharov, N.P. Exploring the Origins of Hexaploid Wheats: Typification of Archaeological Triticum vulgare Var. Antiquorum and Description of Modern Triticum sphaerococcum subsp. Antiquorum (Poaceae: Triticeae). Taxonomy 2024, 4, 780–794. [Google Scholar] [CrossRef]
- Nesbitt, M. When and Where Did Domesticated Cereals First Occur in Southwest Asia? In The Dawn of Farming in the Near East; Cappers, R.T.J., Bottema, S., Eds.; Ex Oriente: Berlin, Germany, 2002. [Google Scholar]
- Arzani, A.; Ashraf, M. Cultivated Ancient Wheats (Triticum spp.): A Potential Source of Health-Beneficial Food Products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K. Vorarbeiten Zur Monographischen Darstellung von Wildpflanzensortimenten: Aegilops L.—Resistenzuntersuchungen. Die Kult. 1985, 33, 123–131. [Google Scholar] [CrossRef]
- Lubbers, E.L.; Gill, K.S.; Cox, T.S.; Gill, B.S. Variation of Molecular Markers among Geographically Diverse Accessions of Triticum tauschii. Genome 1991, 34, 354–361. [Google Scholar] [CrossRef]
- Dudnikov, A.J.; Goncharov, N.P. Allozyme Variation in Aegilops squarrosa. Hereditas 1993, 119, 117–122. [Google Scholar] [CrossRef]
- Dvorak, J.; Luo, M.; Akhunov, E.D.N.I. Vavilov’s Theory of Centres of Diversity in the Light of Current Understanding of Wheat Diversity, Domestication and Evolution. Czech J. Genet. Plant Breed. 2011, 47, S20–S27. [Google Scholar] [CrossRef]
- Kihara, H.; Tanaka, M. Morphological and Physiological Variation among Aegilops squarrosa Strains Collected in Pakistan, Afganistan and Iran. Preslia 1958, 30, 241–251. [Google Scholar]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of Polyploid Wheat Genomic Diversity Using a High-Density 90,000 Single Nucleotide Polymorphism Array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef]
- Hammer, K. Vorarbeiten Zur Monographischen Darstellung von Wildpflanzensortimenten: Aegilops L. Die Kult. 1980, 28, 33–180. [Google Scholar] [CrossRef]
- Kihara, H.; Yamashita, K.; Tanaka, M. Morphological, Physiological, Genetic and Cytological Studies in Aegilops and Triticum Collected from Pakistan, Afghanistan and Iran. In Results of the Kyoto University Scientific Expedition to the Karakoram and Hindukush; Yamashita, K., Ed.; Kyoto University: Kyoto, Japan, 1965; pp. 1–118. [Google Scholar]
- Tzvelev, N.N.; Probatova, N.S. Grasses of Russia; KMK: Moscow, Russia, 2019. (In Russian) [Google Scholar]
- Matsuoka, Y.; Nishioka, E.; Kawahara, T.; Takumi, S. Genealogical Analysis of Subspecies Divergence and Spikelet-Shape Diversification in Central Eurasian Wild Wheat Aegilops tauschii Coss. Plant Syst. Evol. 2009, 279, 233–244. [Google Scholar] [CrossRef]
- Reza, A.; Mohammad, T.Æ.; Naghavi, R. Distribution and Diversity of Aegilops tauschii in Iran. Genet. Resour. Crop Evol. 2008, 55, 341–349. [Google Scholar] [CrossRef]
- Jaaska, V. Electrophoretic Survey of Seedling Esterases in Wheats in Relation to Their Phylogeny. Theor. Appl. Genet. 1980, 56, 273–284. [Google Scholar] [CrossRef]
- Nishikawa, K.; Furuta, Y.; Wada, T. Genetics Studies on Alpha-Amylase Isozymes in Wheat. III. Jpn. J. Genet. 1980, 55, 325–336. [Google Scholar] [CrossRef]
- Nakai, Y. Isozyme Variations in Aegilops and Triticum, IV. The Origin of the Common Wheats Revealed from the Study on Esterase Isozymes in Synthesized Hexaploid Wheats. Jpn. J. Genet. 1979, 54, 175–189. [Google Scholar] [CrossRef]
- Dvorak, J.; Deal, K.R.; Luo, M.-C.; You, F.M.; von Borstel, K.; Dehghani, H. The Origin of Spelt and Free-Threshing Hexaploid Wheat. J. Hered. 2012, 103, 426–441. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, K. Alpha-Amylase Isozymes and Phylogeny of Hexaploid Wheat. In Proceedings of the 4th International Wheat Genetics Symposium; Sears, E.R., Sears, L.M., Eds.; Missouri Agricultural Experiment Station: Columbia, MO, USA, 1973; pp. 851–855. [Google Scholar]
- Lelley, T.; Stachel, M.; Grausgruber, H.; Vollmann, J. Analysis of Relationships between Aegilops tauschii and the D Genome of Wheat Utilizing Microsatellites. Genome 2000, 43, 661–668. [Google Scholar] [CrossRef]
- Mizuno, N.; Yamasaki, M.; Matsuoka, Y.; Kawahara, T.; Takumi, S. Population Structure of Wild Wheat D-Genome Progenitor Aegilops tauschii Coss.: Implications for Intraspecific Lineage Diversification and Evolution of Common Wheat. Mol. Ecol. 2010, 19, 999–1013. [Google Scholar] [CrossRef]
- Cosson, E.-C. Notes Sur Quelques Plantes de France Critiques; Librairie de Victor Masson, Place de l’École-de-Médecine 1: Paris, France, 1849. [Google Scholar]
- Davis, P.H. Aegilops. In Flora of Turkey and the East Aegean Islands; Davis, P., Ed.; Edinburgh University Press: Edinburgh, Scotland, 1985; pp. 163, 232–233, 237, 239, 241, 243. [Google Scholar]
- Bor, N.L. Gramineae. In Flora Iranica; Rechinger, K., Ed.; Akademische Druck-u. Verlagsanstalt: Graz, Austria, 1970; pp. 1–573. [Google Scholar]
- Cope, T. Poaceae. In Flora of Pakistan; Nasir, E., Ali, S.I., Eds.; Fakhri Printing Press: Karachi, Pakistan, 1982; pp. 595–596. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code); Koeltz Botanical: Glashütten, Germany, 2018. [Google Scholar]
- Buxbaum, J.C. Plantarum Minus Cognitarum Centuria I Complectens Plantas Circa Byzantium & in Oriente Observatas.; Ex typographia Academiae: Petropoli, Brazil, 1728. [Google Scholar]
- Trinius, C.B. Clavis Agrostographiae Antiquioris. Uebersicht des Zustandes der Agrostographie Bis Auf Linné; Und Versuch Einer Reduction der Alten Synonyme der Gräser Auf Die Heutigen Trivialnahmen; In der Biedermann’schen Hofbuchhandlung: Coburg, Germany, 1822. [Google Scholar]
- Zhu, Y.J. Aegilops. In Flora de China; Wu, Z., Raven, P.H., Eds.; Missouri Botanical Garden: Beijing, China; St. Louis, MO, USA, 2006; p. 444. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2019. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer-Gallego, P.P.; Ferrer-Gallego, R.; Rivera, D.; Obón, C.; Laguna, E.; Goncharov, N.P. Nomenclature and Typification of the Goat Grass Aegilops tauschii Coss. (Poaceae: Triticeae): A Key Species for the Secondary Gene Pool of Common Wheat Triticum aestivum. Plants 2025, 14, 2375. https://doi.org/10.3390/plants14152375
Ferrer-Gallego PP, Ferrer-Gallego R, Rivera D, Obón C, Laguna E, Goncharov NP. Nomenclature and Typification of the Goat Grass Aegilops tauschii Coss. (Poaceae: Triticeae): A Key Species for the Secondary Gene Pool of Common Wheat Triticum aestivum. Plants. 2025; 14(15):2375. https://doi.org/10.3390/plants14152375
Chicago/Turabian StyleFerrer-Gallego, P. Pablo, Raúl Ferrer-Gallego, Diego Rivera, Concepción Obón, Emilio Laguna, and Nikolay P. Goncharov. 2025. "Nomenclature and Typification of the Goat Grass Aegilops tauschii Coss. (Poaceae: Triticeae): A Key Species for the Secondary Gene Pool of Common Wheat Triticum aestivum" Plants 14, no. 15: 2375. https://doi.org/10.3390/plants14152375
APA StyleFerrer-Gallego, P. P., Ferrer-Gallego, R., Rivera, D., Obón, C., Laguna, E., & Goncharov, N. P. (2025). Nomenclature and Typification of the Goat Grass Aegilops tauschii Coss. (Poaceae: Triticeae): A Key Species for the Secondary Gene Pool of Common Wheat Triticum aestivum. Plants, 14(15), 2375. https://doi.org/10.3390/plants14152375