
Advances in Genetic Diversity of Germplasm Resources, Origin and Evolution of Turnip Rape (Brassica rapa L.)

 ,
,
Abstract
1. Introduction
2. Results
2.1. The Centuries-Old Cultivation and Historical Trajectory of Brassica rapa in China
2.2. Genetic Diversity in Brassica rapa Germplasm Resources
2.3. Diversity in the Utilization of Brassica rapa
2.4. Advances in the Origin Research of Brassica rapa
2.4.1. Phylogenetic Relationships Among Brassica rapa Crops
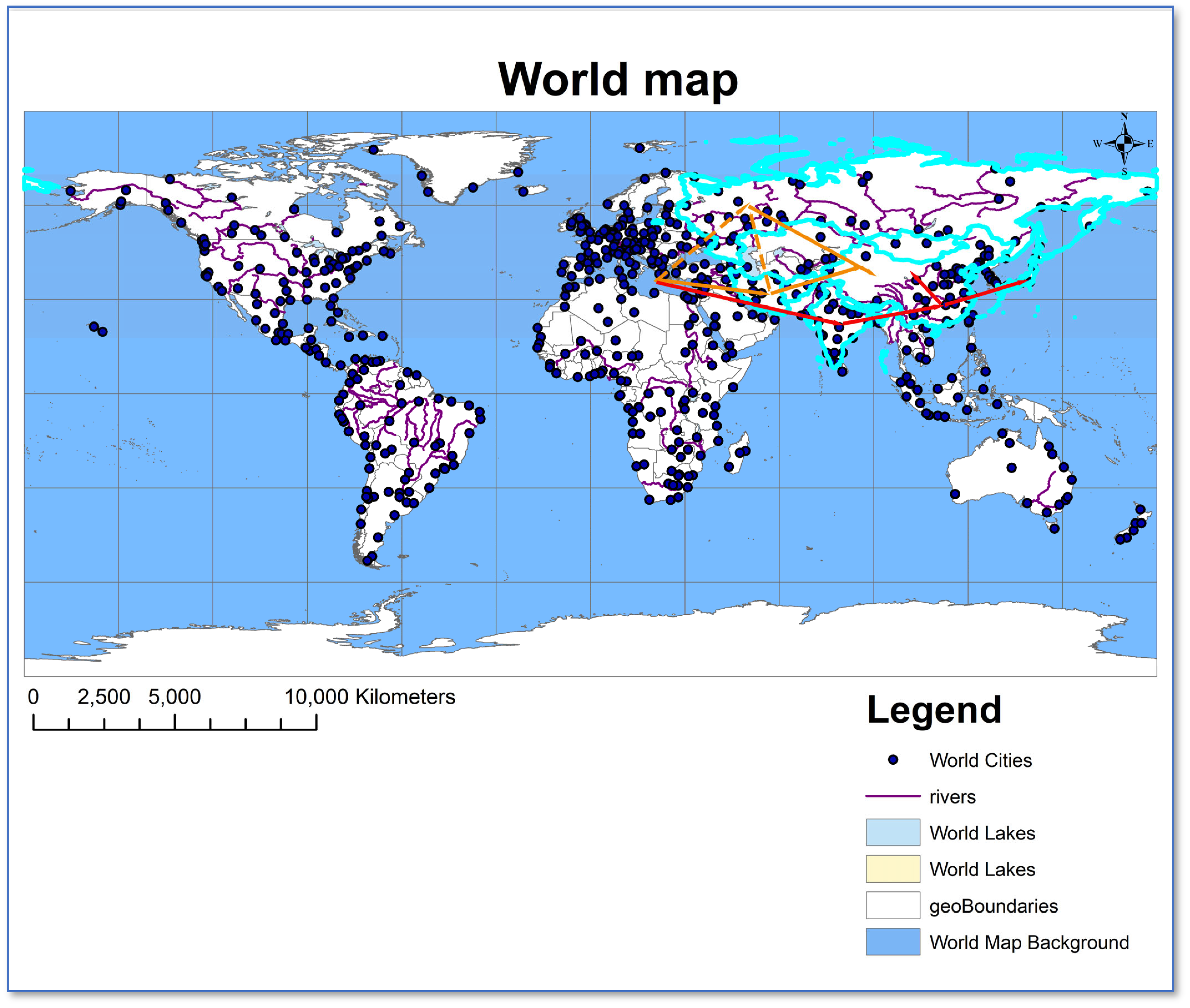
2.4.2. Geographic Origins
2.4.3. Karyotype-Based Analysis
2.4.4. Analysis Based on Genomic Information
High-Throughput Sequencing of Brassica rapa
2.4.5. Advances in Cytoplasmic Genome Research
Progress in Mitochondrial Genome Evolution Studies
Progress in Chloroplast Genome Evolution Studies
3. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, N.N. Study on Cold Resistance and Selecting Indexes Evaluation System of Winter Turnip Rape (Brassica campestris L.). Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2014. [Google Scholar]
- Shang, G.X. Study on the Mechanism of Seed Coat Color Formation in Brassica rapa L. by Multi-Omics Joint Analysis. Master’s Thesis, Qinghai University, Xining, China, 2023. [Google Scholar] [CrossRef]
- He, Y.T.; Chen, B.Y.; Fu, T.D.; Li, D.R.; Tu, J.X. Origin and Evolution of Brassica campestris L. in China. J. Genet. Genom. 2003, 30, 1003–1012. [Google Scholar]
- Laura, P.V.; Miriam, P.M.; Mark, D.W. Brassica rapa CURLY LEAF is a Major H3K27 Methyltransferase Regulating Flowering Time. Planta 2024, 260, 27. [Google Scholar] [CrossRef]
- Li, J.W. The Origin and Variation of Vegetable Crops in China. Sci. Agric. Sin. 1981, 14, 90–95. [Google Scholar]
- Liu, H.L. The Origin and Evolution of Major Brassica Oilseed Crops. Acta Agron. Sin. 1984, 10, 9–18. [Google Scholar]
- Tsunoda, S.; Hinata, K.; Gómez-Campo, C. Brassica Crops and Wild Allies; Japan Scientific Societies Press: Tokyo, Japan, 1980. [Google Scholar]
- Gómez-Campo, C.; Prakash, S. Origin and Domestication. Dev. Plant Genet. Breed. 1999, 4, 33–58. [Google Scholar] [CrossRef]
- Gao, Y.M. Molecular Study on the Classification and Phylogeny of Brassica rapa L. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2009. [Google Scholar]
- Bird, K.A.; An, H.; Gazave, E.; Gore, M.A.; Pires, J.C.; Robertson, L.D.; Labate, J.A. Population Structure and Phylogenetic Relationships in a Diverse Panel of Brassica rapa L. Front. Plant Sci. 2017, 8, 321. [Google Scholar] [CrossRef]
- Su, H.Y. On Relationship Between the First Stage of Dadiwan Culture and the Origin of Chinese Agriculture. J. Northwest AF Univ. (Soc. Sci. Ed.) 2009, 9, 97–101. [Google Scholar]
- Liu, Z.M. Origin and Development of Cultivated Rice in China. J. Genet. Genom. 1975, 23–30. [Google Scholar]
- Wang, J.L.; Luan, Y.F.; Daci, Z.G.; Zhang, Y.Q.; Chang, T.J.; Fang, H.L.; Cheng, H.H.; He, Y. Origin and Evolution of Cultivated Rapeseeds in China. Crop Res. 2006, 199–205. [Google Scholar] [CrossRef]
- He, S.Q. Overview of Pre-Qin Agricultural Archaeology in Gansu. Agric. Archaeol. 1987, 55–61. [Google Scholar]
- Tong, P.Y. Historical Stories of Rapeseed. Agric. Archaeol. 2004, 140–143. [Google Scholar]
- Wang, Y.X. Questions on the Annotations of the Book of Songs. Xinzhu J. (Soc. Sci. Ed.) 1984, 51–57. [Google Scholar]
- Breeding Group of Baoshan Prefecture Agricultural Science Institute. Analysis of Main Technical Experiences in Rapeseed Production. Yunnan Agric. Sci. Technol. 1981, 19–22. [Google Scholar]
- Wei, J.C. Several Issues on Rapeseed Virus Disease. Sci. Agric. Sin. 1961, 20–22. [Google Scholar]
- Qiu, W.F. Current Research Status and Future Directions of Viral Diseases in Cruciferous Crops in China. J. Plant Prot. 1962, 403–408. [Google Scholar]
- Yang, B.; Liu, Z.S.; Xiao, H.G.; Rao, Y.; Tang, R.; Zhang, C.; Wang, L.L. Advances on Research of Distant Hybridizations for Breeding Swede Rapeseed (Brassica napus). J. Plant Genet. Resour. 2021, 22, 593–602. [Google Scholar] [CrossRef]
- Atta, O.F.; Antje, S.H.; Heiko, C.B. Biomass Yield and Heterosis of Crosses Within and Between European Winter Cultivars of Turnip Rape (Brassica rapa L.). J. Appl. Genet. 2011, 53, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Rapeseed Variety Wuyou1. Jiangxi Agric. Sci. Technol. 1977, 24, 48. [Google Scholar]
- Guan, C.X. Breeding of Brassica rapa Should Be Prioritized. J. Anhui Agric. Sci. 1985, 21–23. [Google Scholar]
- Anonymous. Baiyou1. Hubei Agric. Sci. 1972, 48. [Google Scholar]
- Zhang, C.W.; Qin, M.Q.; Lu, X.Q.; Yang, S.J. Breeding of Virus-Resistant Brassica rapa Varieties: Xiezuo1 and Xiezuo2. Plant Prot. 1963, 111–114. [Google Scholar]
- Hao, T.L. Winter Brassica rapa Variety Jinyou 2. Chin. J. Oil Crop Sci. 1985, 79. [Google Scholar]
- Ma, X.G.; Chen, L.H.; Tian, Z.K.; Du, D.Z.; Feng, Q.H.; Yuan, M.Y.; Zhang, Y.C.; Zhang, Z.L.; Sun, H.L. Research on Crop Breeding and Cultivation Techniques. Sci. Technol. Qinghai Agric. 2001, 10–31. [Google Scholar]
- Li, G.M. New Rapeseed Variety: Wanyou 7. Bull. Agric. Sci. Technol. 1993, 32–33. [Google Scholar]
- Sun, W.C.; Zhang, T.; Yang, Y.P. Breeding of Low-Sulfur, Medium-Glucosinolate Spring Brassica rapa Variety Longyou3. Chin. Agric. Sci. Bull. 1998, 69–71. [Google Scholar]
- Zhang, T.; Sun, W.C.; Yang, Y.P. Breeding Report of Double-Low Spring Brassica rapa Variety Longyou4; Gansu Agricultural Science and Technology: Lanzhou, China, 2002; pp. 10–11. [Google Scholar]
- Sun, W.C.; Zhang, C.S.; Tang, S.; Liu, Z.G.; Wu, J.Y.; Zeng, X.C.; Dong, Y.; Wang, J.J.; Geng, Y.G.; Li, X.C.; et al. Breeding of Strong-Cold-Tolerant Winter Rapeseed Variety Longyou7. Scientific Achievements. 2009. Available online: https://kns.cnki.net/kcms2/article/abstract?v=hQuCc5bkPPMeAuXZnS61BJDfJByfCHvYwO6jsEvaclFAZ90dOP-H8oMo4CFge_Q4jbFx2WY8kTDXJaCz5EJxykX7B_4N-zcynqTJ-MixR6VJXhfWL5y9Fig9ukPZCjsVvhmmukf_Q7lKtvHLjTtBnOVynsV_sFFdMS42eHBE8OuOTrOxuXei9skCc9vgQUS5&uniplatform=NZKPT&language=CHS (accessed on 5 June 2025).
- Gao, B.Z.; Liu, B.; Li, S.K.; Liang, J.L.; Cheng, F.; Wang, X.W.; Wu, J. Genome-Wide Association Studies for Flowering Time in Brassica rapa. Sci. Agric. Sin. 2017, 50, 3375–3390. [Google Scholar]
- Huang, C. Evaluation of Cold Resistance and Transcriptome Analysis of Cabbage-Type Oilseed Rape in Tibet. Master’s Thesis, Tibet Agricultural and Animal Husbandry University, Linzhi, China, 2024. [Google Scholar] [CrossRef]
- Guan, Y. Use of Brassica rapa. Crop Res. 2013, 27, 526–528. [Google Scholar]
- Tanhuanpää, P.; Erkkilä, M.; Tenhola, R.T.; Tanskanen, J.; Manninen, O.; Bell, J.B. SNP Diversity Within and Among Brassica rapa Accessions Reveals No Geographic Differentiation. Genome 2016, 59, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Osborn, T.C.; Williams, P.H. Brassica Taxonomy Based on Nuclear Restriction Fragment Length Polymorphisms (RFLPs). Theor. Appl. Genet. 1990, 79, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.M.; Zou, Y.; Liu, Z.G.; Mi, W.B.; Xu, M.X.; Dong, X.Y.; Cao, X.D.; Zheng, G.Q.; Fang, X.L. Physiological and Biochemical Characteristics of Low Temperature Vernalization of Germinating Seeds of Brassica rapa. Sci. Agric. Sin. 2020, 53, 929–941. [Google Scholar]
- Miao, Y.; Li, Y.F.; Kong, J.P.; Rexidan, A.M.T.; Wei, H.P.; Wang, X.G.; Ye, X.J.; Xing, Y.P.; Zeng, X.H. Effects of Different Sowing Dates on Growth Period and Yield of Winter Rapeseed Longyou7. Rural. Sci. Technol. 2014, 8–9. [Google Scholar]
- Pan, Q.Y. Winter Turnip in Gansu and Its Utilization Value. Chin. J. Oil Crop Sci. 1987, 79–81. [Google Scholar]
- Chen, B.Y.; Xu, K.; Gao, G.Z.; Yan, G.X.; Li, J.; Wu, X.M. Analysis of Phenotypic Diversity in Chinese Collection of Brassica rapa L. Chin. J. Oil Crop Sci. 2012, 34, 25–32. [Google Scholar]
- Hu, S.Y.; Wang, J.L.; Luan, Y.F.; Dan, B.; Ni, M.Z.M.; Zhuo, G.; Meng, X.; Tang, L. Geographical Distribution and Phenotypic Differentiation of Tibetan Brassica rapa (Turnip Rape) Resources. Tibet. Sci. Technol. 2002, 44–46+40. [Google Scholar]
- Chen, J.; Tao, G.X. Identification of Native Rape Germplasm Resources of Baicai Type in Guizhou; Guizhou Agricultural Sciences: Guiyang, China, 1999; pp. 25–26. [Google Scholar]
- Yang, G.H.; Zhao, C.X.; Wang, J.X.; Tang, L. Analysis and Evaluation of Growth Period and Yield Traits in Rapeseed Germplasm Resources. Xizang Sci. Technol. 2022, 3–7, 15. [Google Scholar]
- Li, P.; Ma, L.; Xu, F.; Liu, L.J.; Yao, Y.L.; Pu, Y.Y.; Wang, W.T.; Li, X.C.; Fang, Y.; Sun, W.C.; et al. Comparison of Morphological and Physiological Responses of Two Different Species of Northern Winter Rapeseed Before Overwintering. Agric. Res. Arid. Areas 2022, 40, 42–51. [Google Scholar]
- Ma, L.; Lian, Y.T.; Li, S.Y.; Fahim, A.M.; Hou, X.F.; Liu, L.J.; Pu, Y.Y.; Yang, G.; Wang, W.T.; Wu, J.Y.; et al. Integrated transcriptome and metabolome analysis revealed molecular regulatory mechanism of saline-alkali stress tolerance and identified bHLH142 in winter rapeseed (Brassica rapa). Int. J. Biol. Macromol. 2025, 295, 139542. [Google Scholar] [CrossRef]
- Liu, H.Q.; Sun, W.C.; Liu, Z.G.; Wu, J.Y.; Yang, J.S.; Qian, W.; Zhao, Y.N.; Liu, L.B. Analysis of Cold Hardness Adaptability of Winter Rapeseed (Brassica rapa L.) in the Cold and Arid Regions of Northern China. Acta Agric. Boreali-Occident. Sin. 2014, 23, 109–117. [Google Scholar]
- Li, Q.; Gu, Y.G.; Jia, D.H.; Chen, Y.H. Study on Introduction Possibility of Winter Rapeseed to Dry and Cold Areas in Xinjiang China. Acta Agric. Boreali-Occident. Sin. 2011, 20, 106–111. [Google Scholar]
- Li, G.Q. Dominant Factors Analysis of the Major Influential Factors to the Yield of B. Campestris in East Heilongjiang and Its Adapted Varieties. Heilongjiang Agric. Sci. 2000, 23–25. [Google Scholar]
- McAlvay, A.C.; Ragsdale, A.P.; Mabry, M.E.; Qi, X.; Bird, K.A.; Velasco, P.; An, H.; Pires, J.C.; Emshwiller, E. Brassica rapa Domestication: Untangling Wild and Feral Forms and Convergence of Crop Morphotypes. Mol. Biol. Evol. 2021, 38, 3358–3372. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.P.; Liu, X.L.; Li, S.Q.; Liu, X.J.; Hu, Y.Q.; Chen, T. Obtaining and genetic analysis of Hybrid Progeny Between Brassica napus cv. Xiangyou15 and Brassica campestris ssp. pekinensis. Acta Agric. Zhejiangensis 2021, 33, 1170–1176. [Google Scholar]
- Luo, Y.X.; Du, D.Z.; Yu, Q.L. Inbreeding Depression of Qinghai Dahuang Rape. J. Qinghai Univ. 2007, 46–48. [Google Scholar]
- Yuan, Y.; Guo, S.P.; De, Q.; De, Q.P.C.; Yang, G.H.; Deng, G.Z.; Dan, B. A Preliminary Study on Self-Compatibility of Tibetan Cabbage-type Yellow Seed Rape. J. Plateau Agric. 2020, 4, 454–458. [Google Scholar] [CrossRef]
- Wang, J.Y.; Yang, W.X.; Bian, L.; Liu, M.M.; Fan, T.T.; Wu, J.Y.; Pu, Y.Y.; Liu, L.J.; Ma, L.; Wang, W.T.; et al. Characterization of Main Feature in Self-Compatibility Strong Winter Rapeseed (Brassica rapa L.). Chin. J. Oil Crop Sci. 2025, 47, 289–301. [Google Scholar] [CrossRef]
- Yu, Y.L. Mutant Construction of Candidate Genes in Multilocular Silique of Brassica napus L. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2023. [Google Scholar] [CrossRef]
- Luo, B.; Zhao, W.G.; Li, B.J.; Zhao, Y.J.; Zhu, Y.T.; Wang, H. Research Progress on Cold Tolerance of Winter-Type Brassica rapa and Brassica napus in China. Shaanxi J. Agric. Sci. 2017, 63, 80–83. [Google Scholar]
- Wang, L.P.; Yang, G.; Sun, W.C.; Zhang, Z.L.; Wang, Y.; Dong, H.Y.; Zhao, Y.N.; Kong, D.J.; Lu, M.R.; Yang, N.N.; et al. Cold Resistance and Botanical Characteristics of the Hybrid Prgeny between Winter and Spring Turnip Rapes (Brassica campestris L.). Agric. Res. Arid. Areas 2015, 33, 291–296. [Google Scholar]
- Cheng, H.H. Study on Brassica campestris L. Evolution in Tibet. Master’s Thesis, Tibet University, Lhasa, China, 2008. [Google Scholar]
- Zhao, H.P.; Li, Y.L.; Ma, Q.R.; Xiao, L. Quantitative Genetic Analysis of Yield-Related Traits in Brassica rapa. Jiangsu Agric. Sci. 2023, 51, 92–97. [Google Scholar] [CrossRef]
- An, J.K.; Xu, J.H.; Li, M.H.; He, Y.; Chang, X.; Wang, J.L.; Dan, B. Analysis and Comparisons on Genetic Resources Between Wild and Cultivated Rapeseed in Tibetan. Seed 2016, 35, 55–58. [Google Scholar] [CrossRef]
- Liu, H.L. Progress in Rapeseed Breeding Research in China. Chin. Agric. Sci. Bull. 1988, 18–22. [Google Scholar]
- Zhang, Q.; Zhao, Y.Y.; Tan, Y.F.; Chang, X.K.; Yao, Q.J.; Yuan, Y.X.; Wei, X.C.; Wang, Z.Y.; Zhang, X.W. Study on the Brassica napus Resynthesis via Interspecific Hybrid between ‘Yellow Sarson’ and Brassica albograbra. Mol. Plant Breed. 2019, 17, 210–216. [Google Scholar] [CrossRef]
- Qian, X.Z. Research Status and Future Prospects of Rapeseed Germplasm Resources in China. China Seed Ind. 1984, 9–11. [Google Scholar] [CrossRef]
- Zhang, H.C. Some Characteristics of the Spring Rape in Xi Zang. J. Yangzhou Univ. (Agric. Life Sci. Ed.) 1981, 43–46+61. [Google Scholar] [CrossRef]
- Liu, X.Y. Preliminary Analysis of Protein Content in Chinese Rapeseed Germplasm Resources. China Seed Ind. 1990, 21–22. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate Structures in Evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Sun, D.Z.; Ren, D.D.; Xie, Y.F.; Yang, Y.Y.; Wang, X.Y. Simultaneous Determination of 13 kinds of Glucosinolates in Rapeseeds by Ultra Performance Liquid Chromatography-Tandem Mass Spectrometry. J. Food Saf. Qual. 2022, 13, 1895–1902. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Tian, Y.L.; Kang, L.; Li, X.P. Research Status of Glucosinolates. Agric. Prod. Process. 2020, 86–88. [Google Scholar] [CrossRef]
- Wang, Y. CRISPR/Cas9-mediated Editing of Thioside Transporter Gene BnaGTRs in Brassica napus. Master’s Thesis, Yangzhou University, Yangzhou, China, 2022. [Google Scholar] [CrossRef]
- Ma, Y.H.; Pu, Y.Y.; Wang, J.X.; Wu, J.Y.; Yang, G.; Zhao, C.X.; Ma, L.; Liu, L.J.; Wang, W.T.; Miao, C.Q.; et al. Analysis of Glucosinolate Content and Component in Brassica rapa L. Sci. Agric. Sin. 2024, 57, 4308–4327. [Google Scholar]
- Feng, X.G.; Yi, J.M.; Shi, T.Y. Isolation, Identification, and Molluscicidal Activity of 2-Hydroxy-3-butenyl Glucosinolate from Rapeseed. Chin. J. Parasitol. Parasit. Dis. 2001, 62–63. [Google Scholar]
- Liang, Q.; Xiong, W.; Zhou, Q.; Cui, C.; Xu, X.; Zhao, L.; Xuan, P.; Yao, Y. Glucosinolates or erucic acid, which one contributes more to volatile flavor of fragrant rapeseed oil? Food Chem. 2023, 412, 135594. [Google Scholar] [CrossRef]
- Ai, X.Y.; El-Badri, A.M.; Batool, M.; Lou, H.; Gao, G.; Bai, C.; Wang, Z.; Jiang, C.; Zhao, X.; Wang, B.; et al. Morpho-Physiochemical Indices and Transcriptome Analysis Reveal the Role of Glucosinolate and Erucic Acid in Response to Drought Stress during Seed Germination of Rapeseed. Int. J. Mol. Sci. 2024, 25, 3308. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, J.W.; Liu, J.; Zhen, C.H.; Li, Q.G.; Song, X.H.; Shen, J.; Zhu, D.Y.; Liu, S.Y.; Liu, K.J. Correlation between Oil Content and Oil Composition Factors in Brassica rapa. South-Cent. Agric. Sci. Technol. 2023, 44, 11–14+21. [Google Scholar]
- Wang, X.F.; Sun, W.C.; Li, X.Z.; Wu, J.Y.; Liu, H.X.; Zeng, C.W.; Pu, Y.Y.; Zhang, P.F.; Zhang, J.J. Wind Erosion-Resistance of Fields Planted with Winter Rapeseed in the Wind Erosion Region of Northern China. Acta Ecol. Sin. 2009, 29, 6572–6577. [Google Scholar]
- Wang, J.Y.; Liu, Y.X.; Zhou, J.H.; Song, H.X.; Tai, S.H.; Li, L. Integrated Management Technologies and Practices for Seasonal Bare Farmlands in Beijing Suburbs. Crops 2011, 1–4. [Google Scholar] [CrossRef]
- Wang, C.L.; Qiang, C.; Wang, J.P.; Cao, C.H.; Zhang, Z.; Chen, W.J.; Yang, J.L. Variety screening of greening manure rapeseed and the mulching effect on soil moisture conservation and fertility in dryland orchard of Loess Plateau. J. Plant Nutr. Fertil. 2023, 29, 1843–1852. [Google Scholar]
- Xu, W.L.; Wang, B. Effects of Different Green Manure Crops Returning to the Field on Subsequent Maize Growth and Yield. China Agric. Technol. Ext. 2024, 40, 89–91. [Google Scholar]
- Zhang, B.S.; Li, G.Q.; Pei, X.Q.; Ju, Q.X.; Zhang, J.L.; Yang, Z.; Niu, S.J.; Cao, S.Q. Effects of Green Fertilizer Winter Rape Multi-cropped in Leisure Period on Nitrogen and Phosphorus Utilization Efficiencies and Yields of Spring Wheat in Dryland Areas of Central Gansu. J. Cold Arid. Agric. Sci. 2024, 3, 926–930. [Google Scholar]
- Sun, W.C.; Pan, Q.Y.; Liu, Z.G.; Meng, Y.X.; Zhang, T.; Wang, H.L.; Zeng, X.C. Genetic resources of oilseed Brassica and related species in Gansu Province, China. Plant Genet. Resour. Charact. Util. 2004, 2, 167–173. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, J.Y.; Zhou, G.C.; Hang, Y.Y.; Sun, X.Q. Cloning and functional analysis on erucic acid synthesis gene FAE1-closest in Brassicaceae plants. J. Plant Resour. Environ. 2023, 32, 45–51. [Google Scholar]
- Lü, L.L.; Zhao, X.B.; Li, X.G.; Liu, D.H. Promising perspects for planting oilseed rape and developing biodiesel industry in the Tibetan Plateau. Biotechnol. Bus. 2019, 32–38. [Google Scholar]
- Guo, Y.; Chen, S.; Li, Z.; Cowling, W.A. Center of Origin and Centers of Diversity in an Ancient Crop, Brassica rapa (Turnip Rape). J. Hered. 2014, 105, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Sun, R.F.; Hou, X.L.; Zheng, H.K.; Zhang, F.L.; Zhang, Y.Y.; Liu, B.; Liang, J.L.; Zhuang, M.; Liu, Y.X.; et al. Subgenome Parallel Selection Is Associated with Morphotype Diversification and Convergent Crop Domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Y. Nomenclature, Classification, and Origin of Cultivated Rapeseed in China. Stud. Hist. Nat. Sci. 1989, 8, 158–165. [Google Scholar]
- Song, K.M.; Osborn, T.C.; Williams, P.H. Brassica Taxonomy Based on Nuclear Restriction Fragment Length Polymorphisms (RFLPs). 2. Preliminary Analysis of Subspecies Within, B. rapa (syn. Campestris) and B. oleracea. Theor. Appl. Genet. 1988, 76, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Wang, X.W.; Deng, B.; Lou, P.; Wu, J.; Sun, R.F.; Xu, Z.Y.; Vromans, J.; Koornneef, M.; Bonnerma, G. Genetic Relationships Within Brassica rapa as Inferred from AFLP Fingerprints. Theor. Appl. Genet. 2005, 110, 1301–1314. [Google Scholar] [CrossRef]
- Li, P.R. The Cytoplasmic Genomes Analysis of Brassica Species. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2017. [Google Scholar] [CrossRef]
- Liu, H.L. Origin and Evolution of Rapeseed. Gansu Agric. Sci. Technol. 1984, 2–5. [Google Scholar]
- Huang, X.Q.; Ma, R.C.; Li, L.; Chen, J.; Dong, Y.P. Genetic Diversity Analysis on Brassica Campestris, L. from Guizhou Province in China. Southwest China J. Agric. Sci. 2009, 22, 271–276. [Google Scholar] [CrossRef]
- Wang, J.L.; He, Y.; Luan, Y.F.; Da, C.Z.G. Multivariate Statistical Analysis of Morphological and Ecological Characteristics of Wild Rapeseed in Tibet. Crop Res. 2006, 223–226. [Google Scholar] [CrossRef]
- Wang, J.L.; Luan, Y.F.; Da, C.Z.G.; Chang, T.J. Geographical distribution and biological characters of wild rapeseed in Tibet. Chin. J. Oil Crop Sci. 2006, 134–137. [Google Scholar]
- Song, K.M.; Suzuki, J.Y.; Slocum, M.K.; Williams, P.M.; Osborn, T.C. A linkage map of Brassica rapa (syn. campestris) based on restriction fragment length polymorphism loci. Theor. Appl. Genet. 1991, 82, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.G. A Multidisciplinary Study of the Taxonomy and Origin of Brassica Crops. BioScience 1977, 27, 35–40. [Google Scholar] [CrossRef]
- Chen, Y.P.; Cao, J.S.; Ying, M.; Ye, W.Z. Analysis of Genetic Polymorphism in Vegetable Crops of Brassica Campestris by RAPD Markers. J. Zhejiang Univ. (Agric. Life Sci.) 2000, 26, 131–136. [Google Scholar]
- Guo, J.X.; Zhou, N.Y.; Ma, R.C.; Cao, M.Q. Genetic Diversity in Brassica rapa Revealed by AFLP Molecular Markers. J. Agric. Biotechnol. 2002, 10, 138–143. [Google Scholar]
- Li, M.H.; Ci, R.B.Z.; An, K.J.; He, Y.; Dan, B. Difference on Karyotype Analysis of 2 Local Rape Varieties (Brassica rapa L.) in Tibet. Anhui Agric. Sci. Bull. 2016, 22, 21–23, 146. [Google Scholar] [CrossRef]
- Li, M.X. Size Variation and Evolution of Plant Chromosomes. Bull. Biol. 1985, 14–16. [Google Scholar]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971; pp. 87–90. [Google Scholar]
- Levitzky, G.A. The Karyotype in Systematics. Bull. Appl. Bot. Genet. Plant Breed. 1931, 27, 220–240. [Google Scholar]
- Li, M.H.; Ci, R.B.Z.; An, K.J.; He, Y.; Dan, B. Karyotype study of yellow-seed rape landraces (Brassica rapa L.) of Tibet China. Chin. J. Oil Crop Sci. 2016, 38, 549–554. [Google Scholar]
- Dong, C.J.; Li, X.J.; Sun, X.W. Research Progress on Genetic Diversity and Origin Evolution of Carp Populations in China. J. Fish. China 2020, 44, 1046–1062. [Google Scholar]
- Wu, J.Y.; Xu, X.D.; Liu, L.J.; Ma, L.; Pu, Y.Y.; Wang, W.T.; Hua, X.Y.; Song, J.M.; Liu, K.; Lu, G. A Chromosome-Level Genome Assembly of a Winter Turnip Rape (Brassica rapa, L.) to Explore the Genetic Basis of Cold Tolerance. Front. Plant Sci. 2022, 13, 936958. [Google Scholar] [CrossRef]
- Wang, X.W.; Wang, H.Z.; Wang, J.; Sun, R.F.; Wu, J.; Liu, S.Y.; Bai, Y.Q.; Mun, J.H.; Bancroft, L.; Cheng, F.; et al. The Genome of the Mesopolyploid Crop Species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Akter, A.; Kakizaki, T.; Itabashi, E.; Kunita, K.; Shimizu, M.; Akter, M.A.; Mehraj, H.; Okazaki, K.; Dennis, E.S.; Fujimoto, R. Characterization of FLOWERING LOCUS C 5 in Brassica rapa L. Mol. Breed. 2023, 43, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Belser, C.; Istace, B.; Denis, E.; Dubarry, M.; Baurens, F.C.; Falentin, C.; Genete, M.; Berrabah, W.; Chèvre, A.M.; Delourme, R. Chromosome-Scale Assemblies of Plant Genomes Using Nanopore Long Reads and Optical Maps. Nat. Plants 2018, 4, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.S.; An, H.; Ragsdale, A.P.; Gutenkunst, R.N.; Pires, J.C.; Barker, M.S. Genomic Inferences of Domestication Events are Corroborated by Written Records in Brassica rapa. Mol. Ecol. 2017, 26, 3373–3388. [Google Scholar] [CrossRef] [PubMed]
- Li, C.C. Revealing the Genetic Basis of Male Sterility Based on the Evolution of Citrus Cytoplasmic Genome. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2023. [Google Scholar] [CrossRef]
- Xiao, W.Y.; Wu, X.; Zhou, X.Y.; Zhang, J.; Huang, J.H.; Dai, X.C.; Ren, H.L.; Xu, D.L. Assembly and Comparative Analysis of the First Complete Mitochondrial Genome of Zicaitai (Brassica rapa var. Purpuraria): Insights into Its Genetic Architecture and Evolutionary Relationships. Front. Plant Sci. 2024, 15, 1475064. [Google Scholar] [CrossRef] [PubMed]
- Handa, H. The Complete Nucleotide Sequence and RNA Editing Content of the Mitochondrial Genome of Rapeseed (Brassica napus, L.): Comparative Analysis of the Mitochondrial Genomes of Rapeseed and Arabidopsis Thaliana. Nucleic Acids Res. 2003, 31, 5907–5916. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Guan, R.Z.; Chang, S.X.; Du, T.Q.; Zhang, H.S.; Han, X.; Zhang, X. Substoichiometrically Different Mitotypes Coexist in Mitochondrial Genomes of Brassica napus L. PLoS ONE 2011, 6, e17662. [Google Scholar] [CrossRef]
- Chang, S.X.; Yang, T.T.; Du, T.Q.; Huang, Y.J.; Chen, J.M.; Yan, J.Y.; He, J.B.; Guan, R.Z. Mitochondrial Genome Sequencing Helps Show the Evolutionary Mechanism of Mitochondrial Genome Formation in Brassica. BMC Genom. 2011, 12, 497–508. [Google Scholar] [CrossRef]
- Wu, Z.X.; Hu, K.N.; Yan, M.J.; Song, L.P.; Wen, J.; Ma, C.Z.; Shen, J.X.; Fu, T.D.; Yi, B.; Tu, J.X. Mitochondrial Genome and Transcriptome Analysis of Five Alloplasmic Male-Sterile Lines in Brassica juncea. BMC Genom. 2019, 20, 348–362. [Google Scholar] [CrossRef]
- Shao, D.K.; Ma, Y.D.; Li, X.J.; Ga, S.; Ren, Y.J. The Sequence Structure and Phylogenetic Analysis by Complete Mitochondrial Genome of Kohlrabi (Brassica Oleracea var. Gongylodes L.). Mitochondrial DNA Part B 2021, 6, 2714–2716. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Tanaka, Y.; Terachi, T. Complete Mitochondrial Genome Sequence of Black Mustard (Brassica nigra; BB) and Comparison with Brassica oleracea (CC) and Brassica carinata (BBCC). Genome 2014, 57, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Heng, S.P. The Mitochondrial Genomes and Functions of HAU Cytoplasm Male Sterility Gene in Brassica. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2015. [Google Scholar]
- Liu, G.D. Comparative Mitochondrial Genomes Analysis of Brassica and MSH1-Mediated Leaf Senescence. Master’s Thesis, Zhejiang University, Hangzhou, China, 2016. [Google Scholar]
- Lü, P.J. Assessment of Chloroplast DNA Polymorphism in Brassica oilseeds and Its Related Species. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2011. [Google Scholar]
- Wu, J.Y.; Ma, X.C.; Ma, L.; Fang, Y.; Zhang, Y.H.; Liu, L.J.; Li, X.C.; Zeng, R.; Sun, W.C. Complete Chloroplast Genome Sequence and Phylogenetic Analysis of Winter Oil Rapeseed (Brassica rapa L.). Mitochondrial DNA Part B 2021, 6, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.Z.; Wu, X.M.; Hu, Q.; Li, X.Z. Collection, Identification, Conservation, and Utilization of Rapeseed Germplasm Resources. Chin. J. Oil Crop Sci. 1996, 60–63. [Google Scholar]
- He, T.T.; Ma, Z.Q.; Peng, S.D.; Xia, G.Y.; Lin, L.B.; Xu, X.G. Collection and Evaluation Agronomic Traits of Yunnan Wild Rape. Seed 2010, 29, 64–67. [Google Scholar] [CrossRef]

{kind=link}
| Title of Work | Era | Description of Rapeseed |
|---|---|---|
| Xia Xiaozheng [15] | 3000 BCE (Xia Dynasty) | Gather rapeseed in the first lunar month, and it blooms in the second lunar month. |
| The Book of poetry [16] | 11th–6th century BCE | Pluck the turnip and the radish, but do not judge them by their roots alone. |
| Lv’s Spring and Autumn Annals [15] | 3rd century BCE (Warring States Period) | The finest vegetables include the rapeseed from Yanghua. |
| Popular Prose [15] | The late Eastern Han Dynasty (184 CE–220 CE) | Rapeseed is historically termed Barbarian Vegetable. Legend attributes its name to the Yuntai Rong tribe beyond the northern frontier, who were said to have first cultivated this plant. |
| Qimin Yaoshu [15] | 6th century CE (Northern Wei Dynasty) | For planting mustard seeds, Sichuan mustard, and rapeseed, they should all be sown in the second or third month when there is favorable rainfall. In times of drought, plant them in ridged beds with irrigation. They will ripen and be ready for seed harvest in the fifth month. |
| Tiangong Kaiwu [15] | Ming Dynasty (1637 CE, by Song Yingxing) | Rapeseeds are placed into a cauldron and stir-fried over gentle heat until aromatic, then crushed and steamed. |
| The Compendium of Materia Medicais [13] | Song Dynasty (11th century CE, compiled by Su Song) | Rapeseed bears a slight resemblance to Chinese cabbage (Brassica rapa var. pekinensis) in shape, with green leaves featuring fine serrations. |
| Compendium of Materia Medica [15] | Ming Dynasty (1593 CE, by Li Shizhen) | In the bitterly cold lands of Qiang, Long, Di, and Hu, this vegetable is widely cultivated during the winter months for its ability to endure frost and snow. |
| Traits | Winter-Type Brassica rapa | Spring-Type Brassica rapa |
|---|---|---|
| Botanical Traits | Prostrate growth at the seedling stage, relatively rapid growth of underground parts, large root collar diameter, and well-developed taproot; purple heart leaves, no bolting or budding before winter. | Green heart leaves, yellowish-green leaf color, Erect seedling growth habit and strong spring growth type. |
| Agronomic Traits | Exhibit similar and abundant total silique numbers per plant. | Exhibits significantly lower total silique numbers per plant. |
| Exhibit lower thousand-seed weights. | Exhibits significantly higher thousand-seed weights. | |
| Exhibit greater plant height compared to spring ecotypes. | Exhibits significantly reduced plant height. | |
| Develops a greater number of primary branches. | Develops significantly fewer primary branches. |
| Largest Chromosome | Percentage of Chromosomes with Arm Ratio > 2:1 | ||||
|---|---|---|---|---|---|
| Smallest Chromosome | 0.0 | 0.01–0.50 | 0.51–0.99 | 1.0 | |
| <2:1 | 1A | 2A | 3A | 4A | |
| 2:1–4:1 | 1B | 2B | 3B | 4B | |
| >4:1 | 1C | 2C | 3C | 4C | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Zhang, T.; Ma, Y.; Han, C.; Yang, W.; Pu, Y.; Ma, L.; Wu, J.; Yang, G.; Wang, W.; et al. Advances in Genetic Diversity of Germplasm Resources, Origin and Evolution of Turnip Rape (Brassica rapa L.). Plants 2025, 14, 2311. https://doi.org/10.3390/plants14152311
Lu X, Zhang T, Ma Y, Han C, Yang W, Pu Y, Ma L, Wu J, Yang G, Wang W, et al. Advances in Genetic Diversity of Germplasm Resources, Origin and Evolution of Turnip Rape (Brassica rapa L.). Plants. 2025; 14(15):2311. https://doi.org/10.3390/plants14152311
Chicago/Turabian StyleLu, Xiaoming, Tianyu Zhang, Yuanqiang Ma, Chunyang Han, Wenxin Yang, Yuanyuan Pu, Li Ma, Junyan Wu, Gang Yang, Wangtian Wang, and et al. 2025. "Advances in Genetic Diversity of Germplasm Resources, Origin and Evolution of Turnip Rape (Brassica rapa L.)" Plants 14, no. 15: 2311. https://doi.org/10.3390/plants14152311
APA StyleLu, X., Zhang, T., Ma, Y., Han, C., Yang, W., Pu, Y., Ma, L., Wu, J., Yang, G., Wang, W., Fan, T., Liu, L., & Sun, W. (2025). Advances in Genetic Diversity of Germplasm Resources, Origin and Evolution of Turnip Rape (Brassica rapa L.). Plants, 14(15), 2311. https://doi.org/10.3390/plants14152311