
Mosaic Evolution of Membrane Transporters in Galdieriales

 ,
,  ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
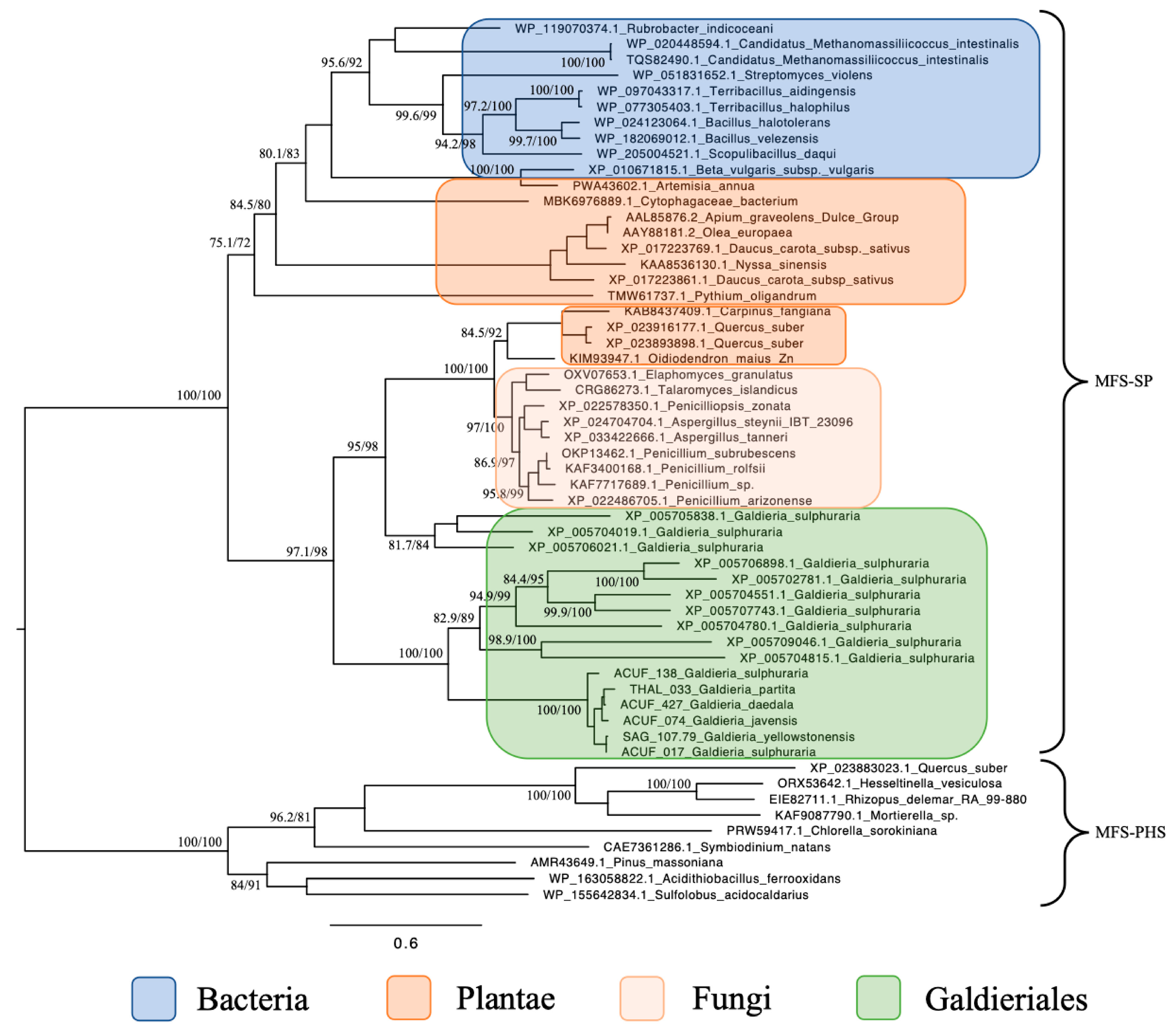
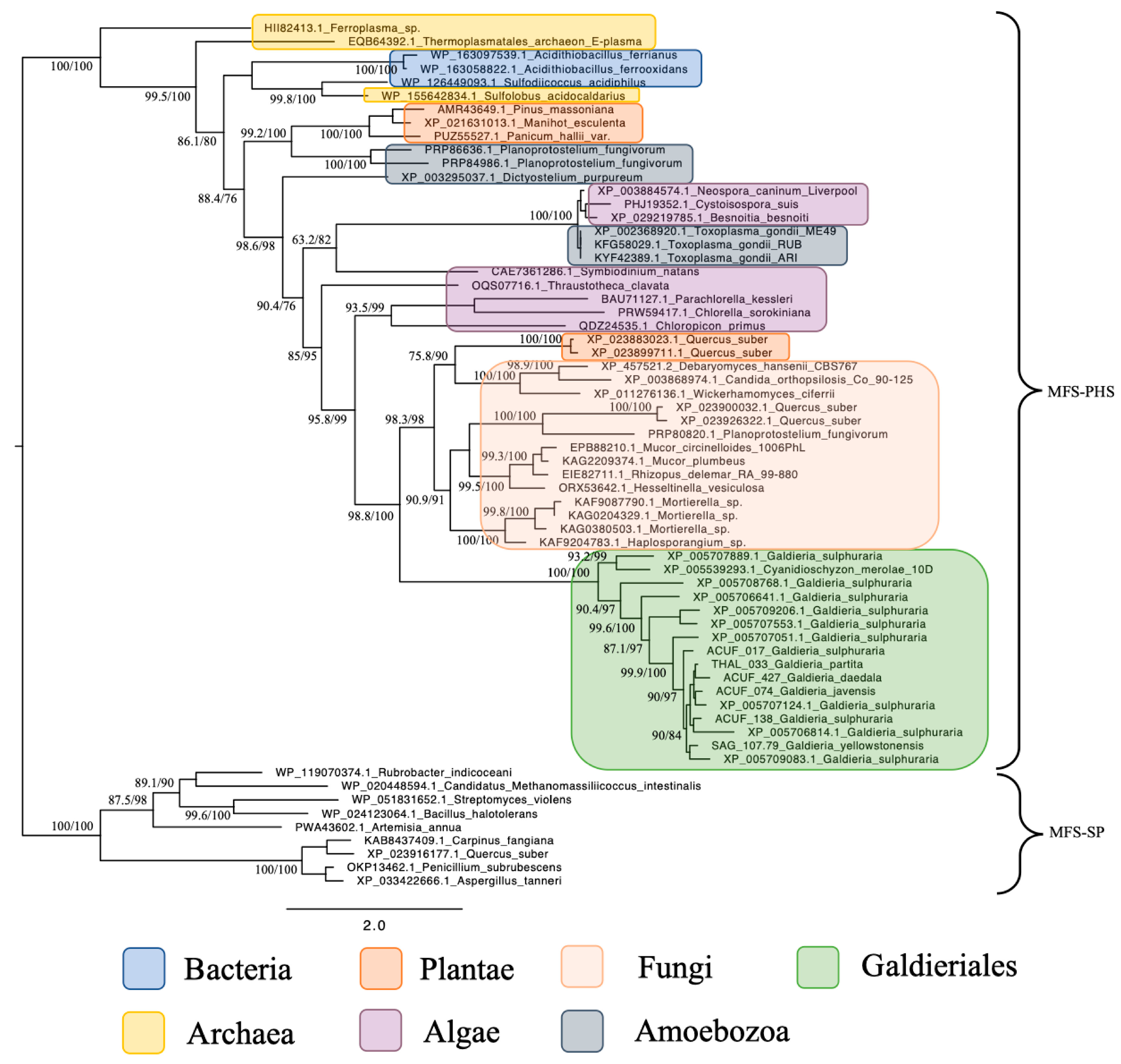
2.1. Major Facilitator Superfamily (MFS) Relationships Reveal Evidence of Polyphyly and Fungal Affinity

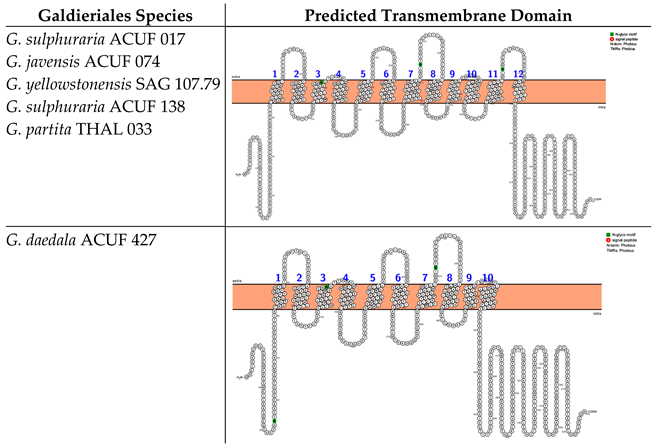
2.2. Amino Acid–Polyamine–Organocation (APC) Family Sequences Clustered Exclusively with Extremophilic Prokaryotes
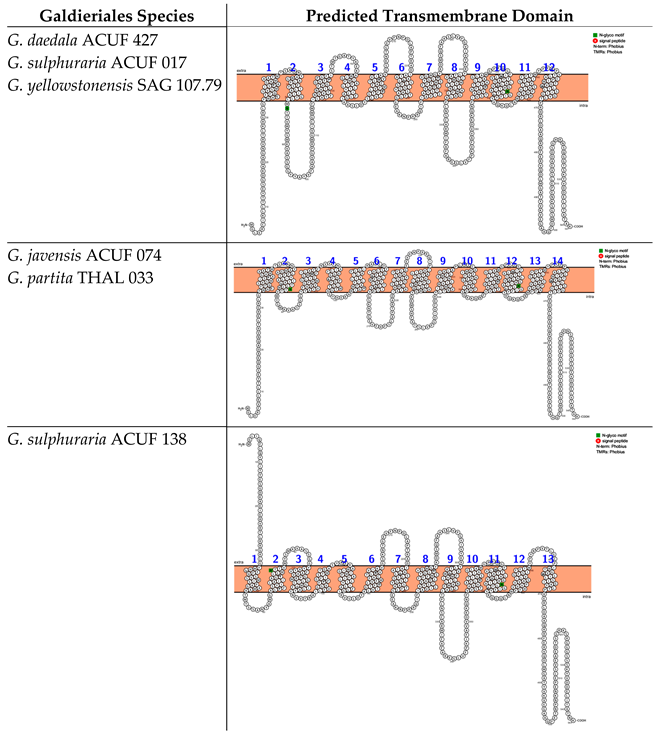
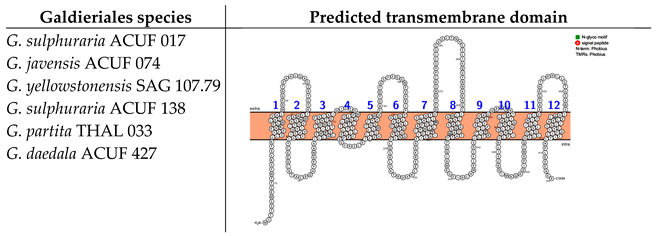
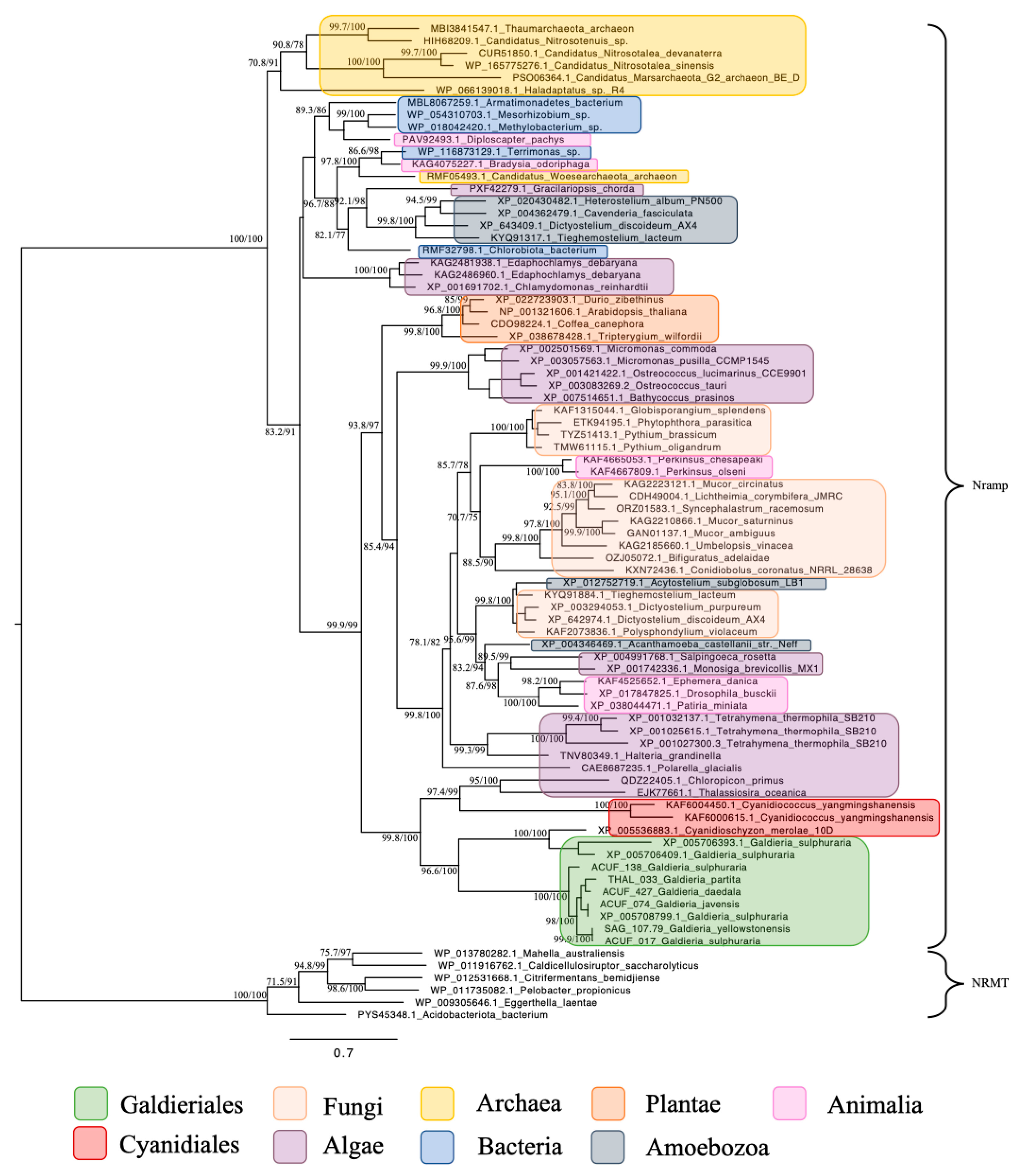
2.3. Nramp (Natural Resistance–Associated Macrophage Protein) Family, Associated with Mn2+ and Fe2+ Transport, Was Widely Distributed Across Eukaryotic and Prokaryotic Lineages
2.4. Protein Structure Prediction and Confidence Assessment
3. Discussion
4. Materials and Methods
4.1. Selection of Target Genes
4.2. Intra-Genus Homolog Discovery
4.3. Prediction of Transmembrane (TM) Domains
4.4. Protein Structure Prediction
4.5. Retrieval of Orthologs from Diverse Taxa
4.6. Sequence Alignment and Curation
4.7. Phylogenetic Tree Reconstruction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, N. Structural Advances for the Major Facilitator Superfamily (MFS) Transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef]
- Jack, D.L.; Paulsen, I.T.; Saier, J. The Amino Acid/Polyamine/Organocation (APC) Superfamily of Transporters Specific for Amino Acids, Polyamines and Organocations. Microbiology 2000, 146, 1797–1814. [Google Scholar] [CrossRef]
- Lanquar, V.; Lelièvre, F.; Barbier-Brygoo, H.; Thomine, S. Regulation and Function of AtNRAMP4 Metal Transporter Protein. Soil. Sci. Plant Nutr. 2004, 50, 1141–1150. [Google Scholar] [CrossRef]
- Vastermark, A.; Wollwage, S.; Houle, M.E.; Rio, R.; Saier, M.H. Expansion of the APC Superfamily of Secondary Carriers. Proteins Struct. Funct. Bioinform. 2014, 82, 2797–2811. [Google Scholar] [CrossRef]
- Farcasanu, I.C.; Mizunuma, M.; Hirata, D.; Miyakawa, T. Involvement of Histidine Permease (Hip1p) in Manganese Transport in Saccharomyces Cerevisiae. Mol. General. Genet. 1998, 259, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Nevo, Y.; Nelson, N. The NRAMP Family of Metal-Ion Transporters. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef]
- Eren, A.; Iovinella, M.; Yoon, H.S.; Cennamo, P.; de Stefano, M.; de Castro, O.; Ciniglia, C. Genetic Structure of Galdieria Populations from Iceland. Polar Biol. 2018, 41, 1681–1691. [Google Scholar] [CrossRef]
- Iovinella, M.; Eren, A.; Pinto, G.; Pollio, A.; Davis, S.J.; Cennamo, P.; Ciniglia, C. Cryptic Dispersal of Cyanidiophytina (Rhodophyta) in Non-Acidic Environments from Turkey. Extremophiles 2018, 22, 713–723. [Google Scholar] [CrossRef]
- Park, S.I.; Cho, C.H.; Ciniglia, C.; Huang, T.Y.; Liu, S.L.; Bustamante, D.E.; Calderon, M.S.; Mansilla, A.; McDermott, T.; Andersen, R.A.; et al. Revised Classification of the Cyanidiophyceae Based on Plastid Genome Data with Descriptions of the Cavernulicolales Ord. Nov. and Galdieriales Ord. Nov. (Rhodophyta). J. Phycol. 2023, 59, 444–466. [Google Scholar] [CrossRef]
- Retta, B.; Iovinella, M.; Ciniglia, C. Significance and Applications of the Thermo—Acidophilic Microalga Galdieria sulphuraria (Cyanidiophytina, Rhodophyta). Plants 2024, 13, 1786. [Google Scholar] [CrossRef]
- Lee, J.; Ghosh, S.; Saier, M.H. Comparative Genomic Analyses of Transport Proteins Encoded within the Red Algae Chondrus Crispus, Galdieria sulphuraria, and Cyanidioschyzon Merolae11. J. Phycol. 2017, 53, 503–521. [Google Scholar] [CrossRef] [PubMed]
- Schönknecht, G.; Chen, W.; Ternes, C.M.; Barbier, G.G.; Shrestha, R.P.; Stanke, M.; Bräutigam, A.; Baker, B.J.; Banfield, J.F.; Garavito, R.M.; et al. Gene Transfer from Bacteria and Archaea Facilitated Evolution of an Extremophilic Eukaryote. Science 2013, 339, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Hanikenne, M.; Krämer, U.; Demoulin, V.; Baurain, D. A Comparative Inventory of Metal Transporters in the Green Alga Chlamydomonas Reinhardtii and the Red Alga Cyanidioschizon merolae. Plant Physiol. 2005, 137, 428–446. [Google Scholar] [CrossRef]
- Henderson, S.W.; Wege, S.; Gilliham, M. Plant Cation-Chloride Cotransporters (CCC): Evolutionary Origins and Functional Insights. Int. J. Mol. Sci. 2018, 19, 492. [Google Scholar] [CrossRef]
- Ramanadane, K.; Straub, M.S.; Dutzler, R.; Manatschal, C. Structural and Functional Properties 1 of a Magnesium Transporter of the SLC11/NRAMP Family. eLife 2022, 11, e74589. [Google Scholar] [CrossRef]
- Sirakov, M.; Palmieri, M.; Iovinella, M.; Davis, S.J.; Petriccione, M.; di Cicco, M.R.; De Stefano, M.; Ciniglia, C. Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold. Plants 2021, 10, 2367. [Google Scholar] [CrossRef] [PubMed]
- Minoda, A.; Miyashita, S.; Fujii, S.; Inagaki, K.; Takahashi, Y. Cell Population Behavior of the Unicellular Red Alga Galdieria sulphuraria during Precious Metal Biosorption. J. Hazard. Mater. 2022, 432, 128576. [Google Scholar] [CrossRef]
- Palmieri, M.; Iovinella, M.; Davis, S.J.; Rosa, M.; Lubritto, C.; Race, M.; Papa, S.; Fabbricino, M.; Ciniglia, C. Galdieria sulphuraria ACUF427 Freeze-Dried Biomass as Novel Biosorbent for Rare Earth Elements. Microorganisms 2022, 10, 2138. [Google Scholar] [CrossRef]
- Iovinella, M.; Lombardo, F.; Ciniglia, C.; Palmieri, M.; di Cicco, M.R.; Trifuoggi, M.; Race, M.; Manfredi, C.; Lubritto, C.; Fabbricino, M.; et al. Bioremoval of Yttrium (III), Cerium (III), Europium (III), and Terbium (III) from Single and Quaternary Aqueous Solutions Using the Extremophile Galdieria sulphuraria (Galdieriaceae, Rhodophyta). Plants 2022, 11, 1376. [Google Scholar] [CrossRef]
- Iovinella, M.; Palmieri, M.; Papa, S.; Auciello, C.; Ventura, R.; Lombardo, F.; Race, M.; Lubritto, C.; di Cicco, M.R.; Davis, S.J.; et al. Biosorption of Rare Earth Elements from Luminophores by G. sulphuraria (Cyanidiophytina, Rhodophyta). Environ. Res. 2023, 239, 117281. [Google Scholar] [CrossRef]
- Adams, E.; Maeda, K.; Kamemoto, Y.; Hirai, K.; Apdila, E.T.; Tarakhovskaya, R.; Adams, E.; Maeda, K.; Kamemoto, Y.; Hirai, K.; et al. Contribution to a Sustainable Society: Biosorption of Precious Metals Using the Microalga Galdieria. Int. J. Mol. Sci. 2024, 25, 704. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, C.; Amoruso, A.J.; Ciniglia, C.; Iovinella, M.; Palmieri, M.; Lubritto, C.; El Hassanin, A.; Davis, S.J.; Trifuoggi, M. Selective Biosorption of Lanthanides onto Galdieria sulphuraria. Chemosphere 2023, 317, 137818. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, A.W.; Price, D.C.; Seger, M.; Lyska, D.; Lammers, P.; Bhattacharya, D.; Weber, A.P.M. The Genomes of Polyextremophilic Cyanidiales Contain 1% Horizontally Transferred Genes with Diverse Adaptive Functions. eLife 2019, 8, e45017. [Google Scholar] [CrossRef]
- Chaloupka, R.; Courville, P.; Veyrier, F.; Knudsen, B.; Tompkins, T.A.; Cellier, M.F.M. Identification of Functional Amino Acids in the Nramp Family by a Combination of Evolutionary Analysis and Biophysical Studies of Metal and Proton Cotransport in Vivo. Biochemistry 2005, 44, 726–733. [Google Scholar] [CrossRef]
- Bozzi, A.T.; Zimanyi, C.M.; Nicoludis, J.M.; Lee, B.K.; Zhang, C.H.; Gaudet, R. Structures in Multiple Conformations Reveal Distinct Transition Metal and Proton Pathways in an Nramp Transporter. eLife 2019, 8, e41124. [Google Scholar] [CrossRef]
- Valdés, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R.; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans Metabolism: From Genome Sequence to Industrial Applications. BMC Genom. 2008, 9, 597. [Google Scholar] [CrossRef]
- Cho, C.H.; Park, S.I.; Huang, T.Y.; Lee, Y.; Ciniglia, C.; Yadavalli, H.C.; Yang, S.W.; Bhattacharya, D.; Yoon, H.S. Genome-Wide Signatures of Adaptation to Extreme Environments in Red Algae. Nat. Commun. 2023, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.; Cho, C.H.; Yoon, H.S.; Bhattacharya, D. Extremophilic Red Algae as Models for Understanding Adaptation to Hostile Environments and the Evolution of Eukaryotic Life on the Early Earth. Semin. Cell Dev. Biol. 2022, 134, 4–13. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive Protein Feature Visualization and Integration with Experimental Proteomic Data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. A Combined Transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting Modern Challenges in Visualization and Analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent Developments in the MAFFT Multiple Sequence Alignment Program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-Based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Yuan, L.; Lu, H.; Li, F.; Nielsen, J.; Kerkhoven, E.J. HGTphyloDetect: Facilitating the Identification and Phylogenetic Analysis of Horizontal Gene Transfer. Brief. Bioinform. 2023, 24, bbad035. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciniglia, C.; Pollio, A.; Pozzuoli, E.; Licata, M.; Nappi, N.; Davis, S.J.; Iovinella, M. Mosaic Evolution of Membrane Transporters in Galdieriales. Plants 2025, 14, 2043. https://doi.org/10.3390/plants14132043
Ciniglia C, Pollio A, Pozzuoli E, Licata M, Nappi N, Davis SJ, Iovinella M. Mosaic Evolution of Membrane Transporters in Galdieriales. Plants. 2025; 14(13):2043. https://doi.org/10.3390/plants14132043
Chicago/Turabian StyleCiniglia, Claudia, Antonino Pollio, Elio Pozzuoli, Marzia Licata, Nunzia Nappi, Seth J. Davis, and Manuela Iovinella. 2025. "Mosaic Evolution of Membrane Transporters in Galdieriales" Plants 14, no. 13: 2043. https://doi.org/10.3390/plants14132043
APA StyleCiniglia, C., Pollio, A., Pozzuoli, E., Licata, M., Nappi, N., Davis, S. J., & Iovinella, M. (2025). Mosaic Evolution of Membrane Transporters in Galdieriales. Plants, 14(13), 2043. https://doi.org/10.3390/plants14132043