


Altitudinal Variations in Coniferous Vegetation and Soil Carbon Storage in Kalam Temperate Forest, Pakistan

 ,
,  ,
,  ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Growing Stock and Biomass
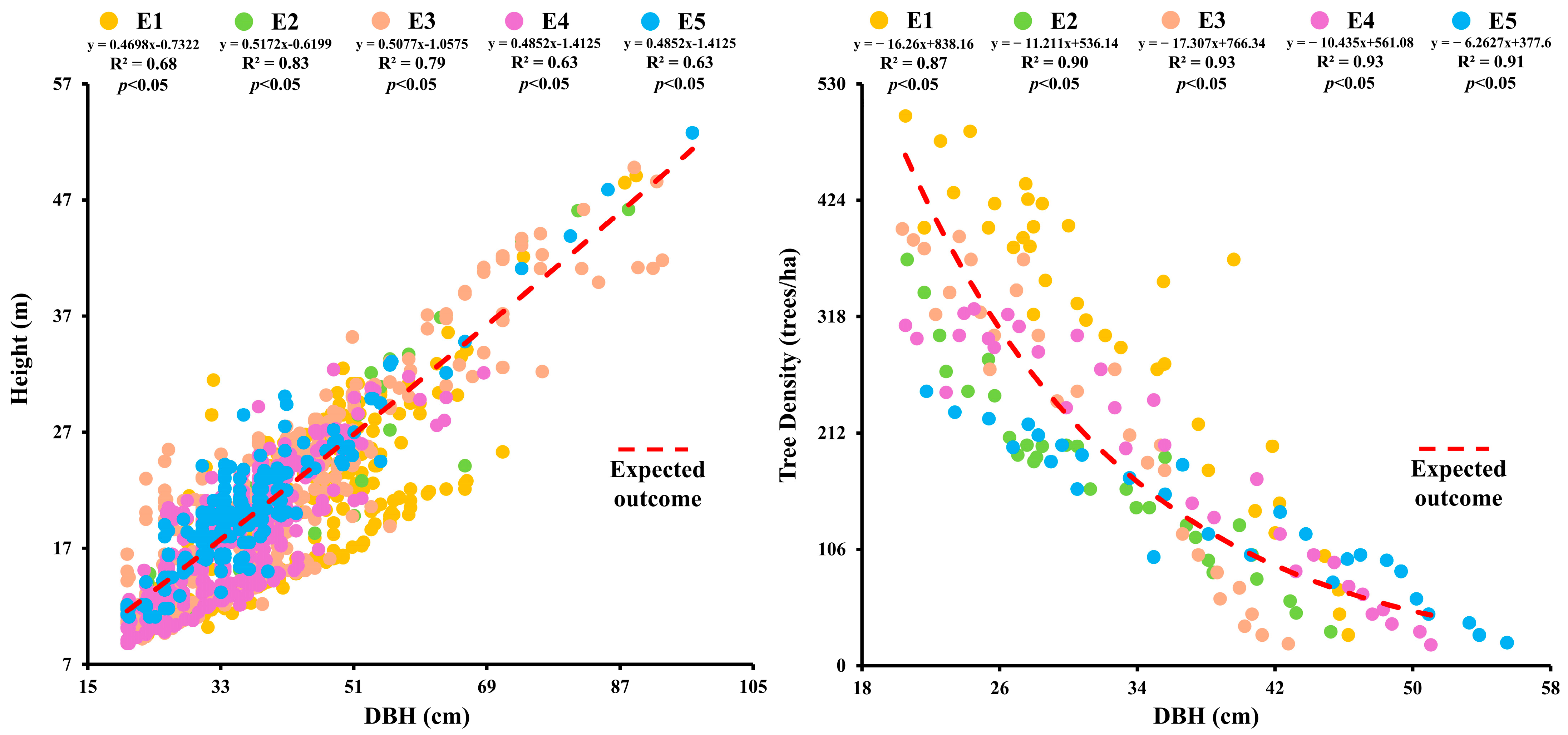
2.2. Association Between DBH, Tree Height, and Stand Density at Different Elevations
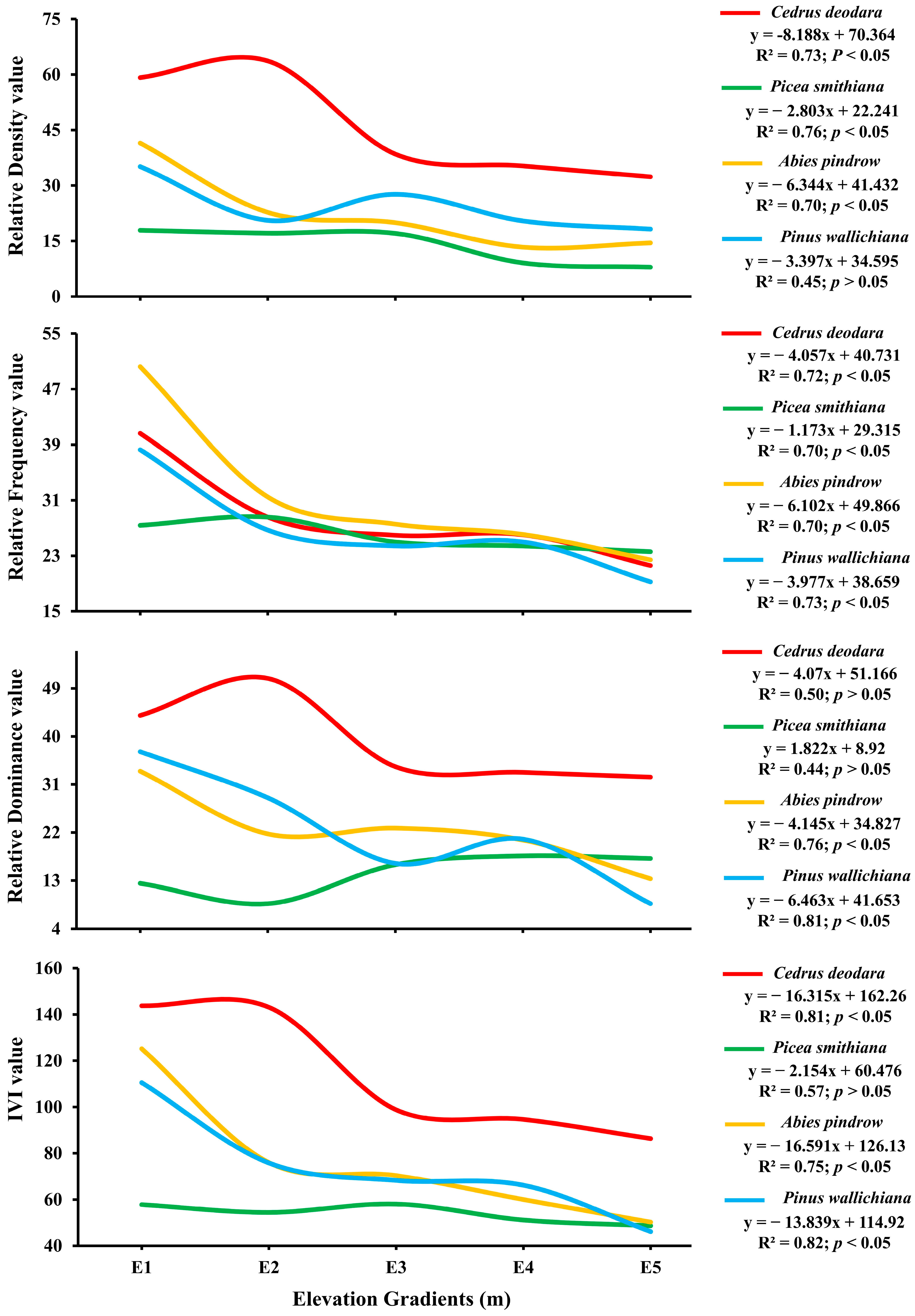
2.3. Association of Species Dominance and Species Diversity with Elevation Gradients
2.4. Association of Litter, Deadwood, and SOCs with Elevation Gradients
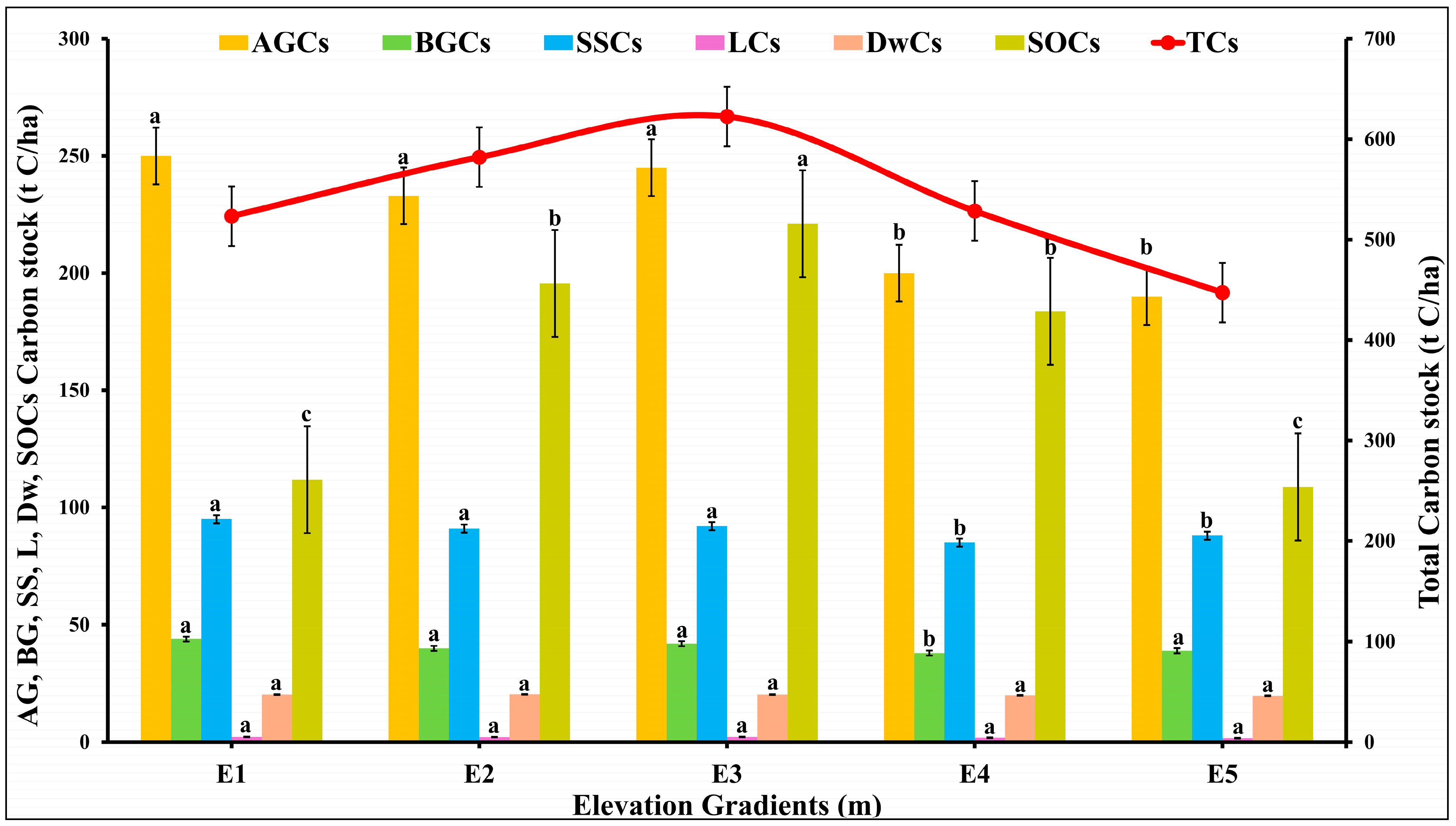
2.5. The Distribution of C Pools over Various Elevations
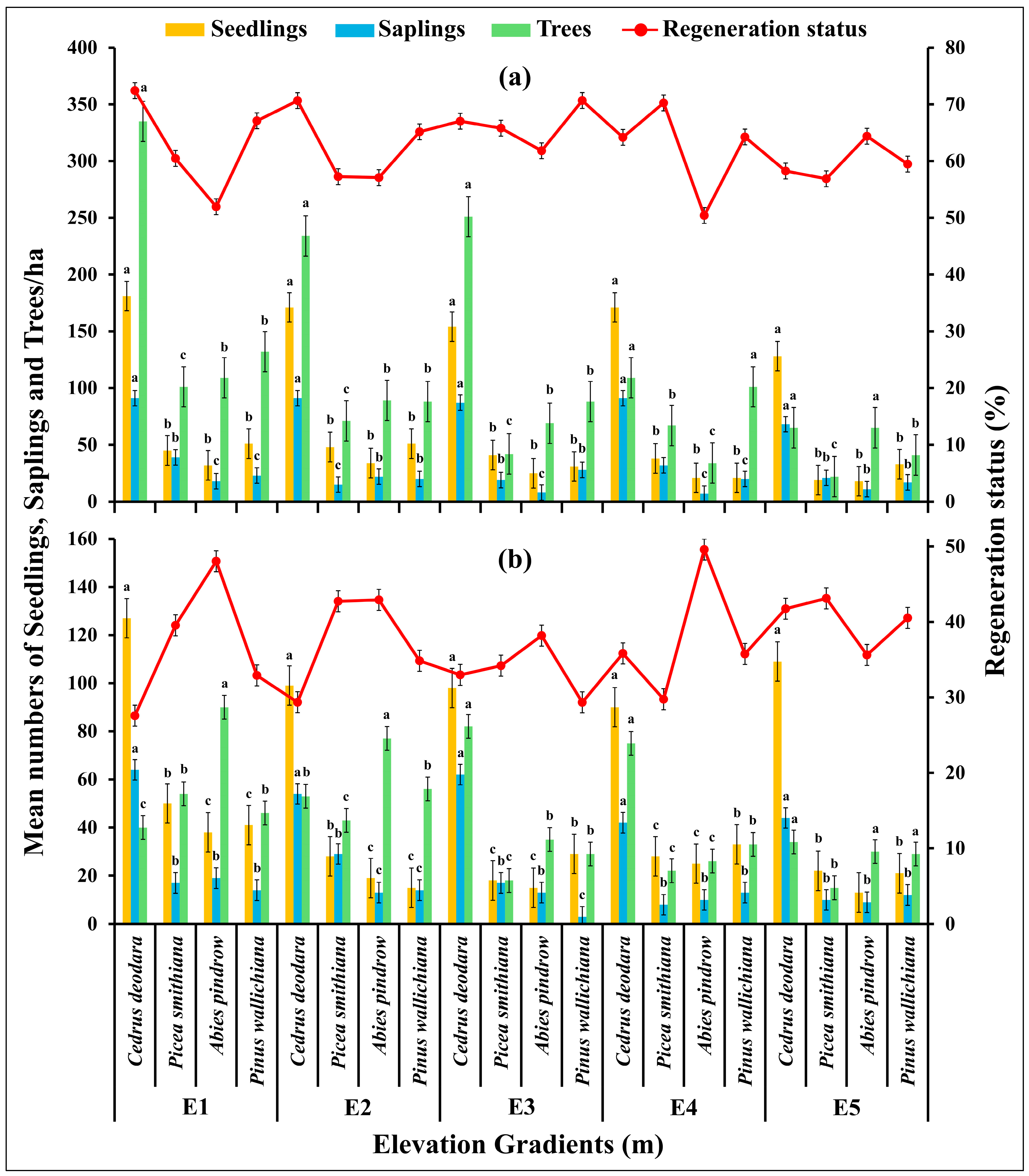
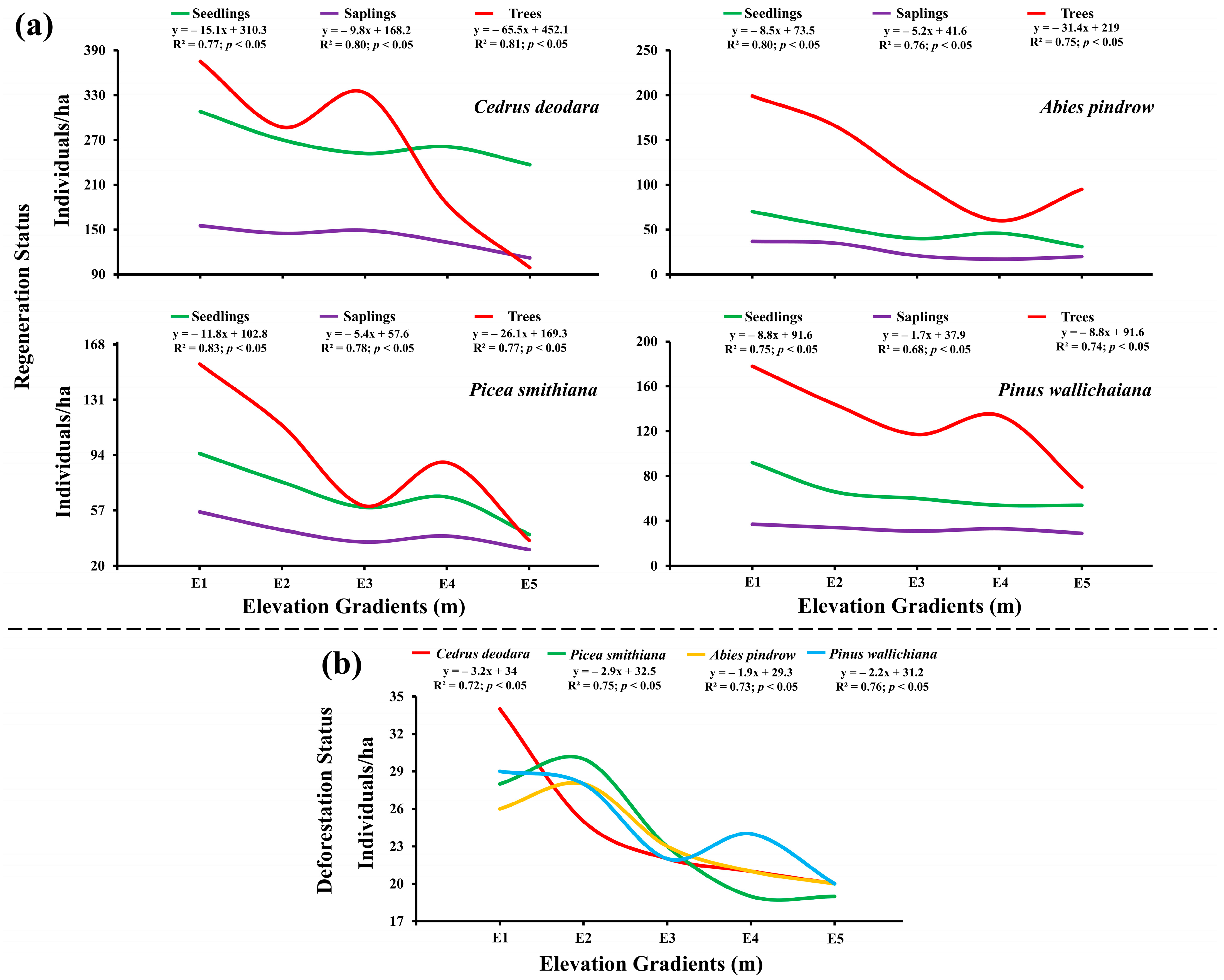
2.6. Regeneration Ability, Number of Trees per Ha and Deforestattion Status of the Conifer Forestat Various Elevation Gradients
2.7. PCA of Environmental and Vegetative Variables
3. Discussion
4. Materials and Methods
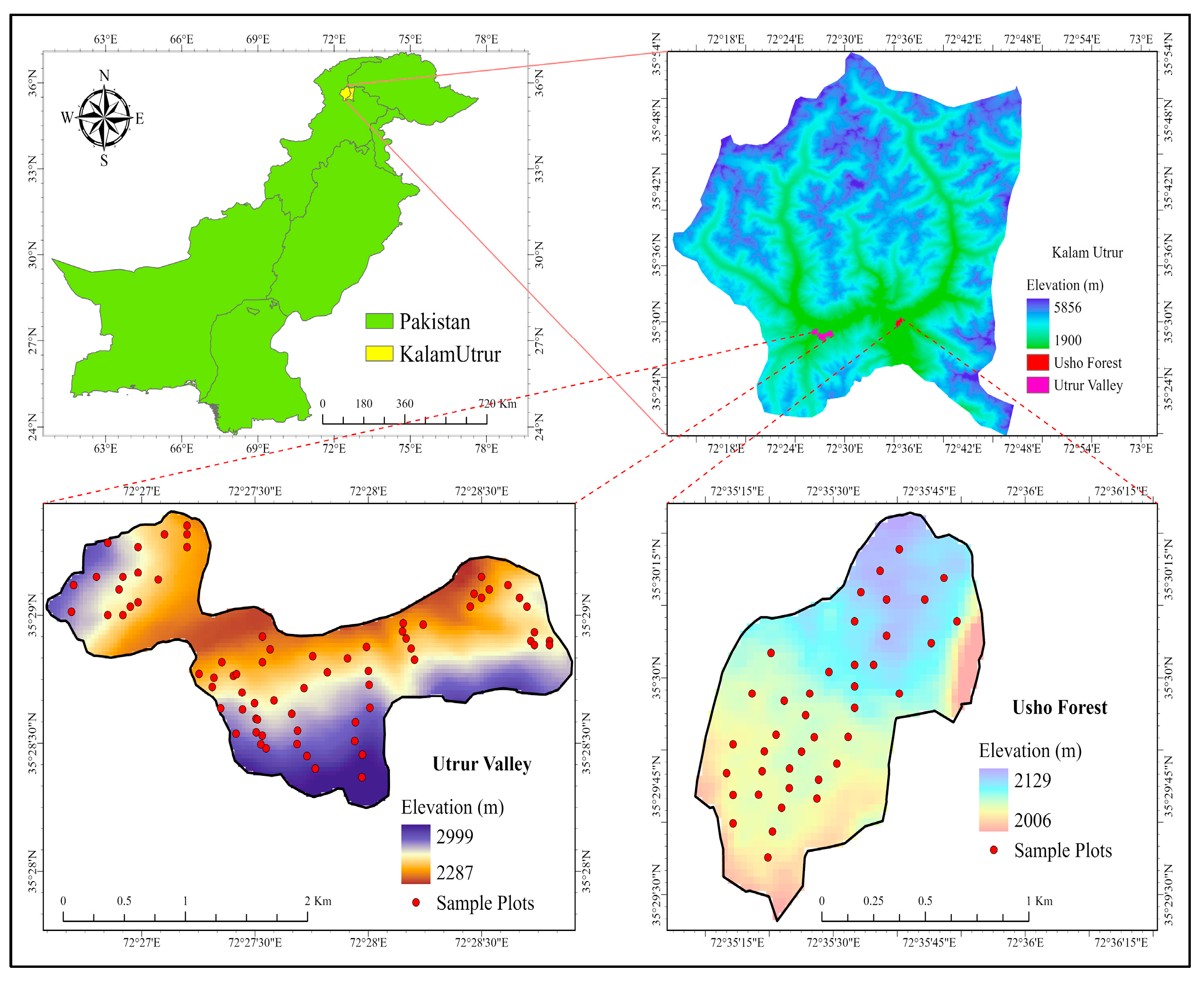
4.1. The Study Area
4.2. Data Collection
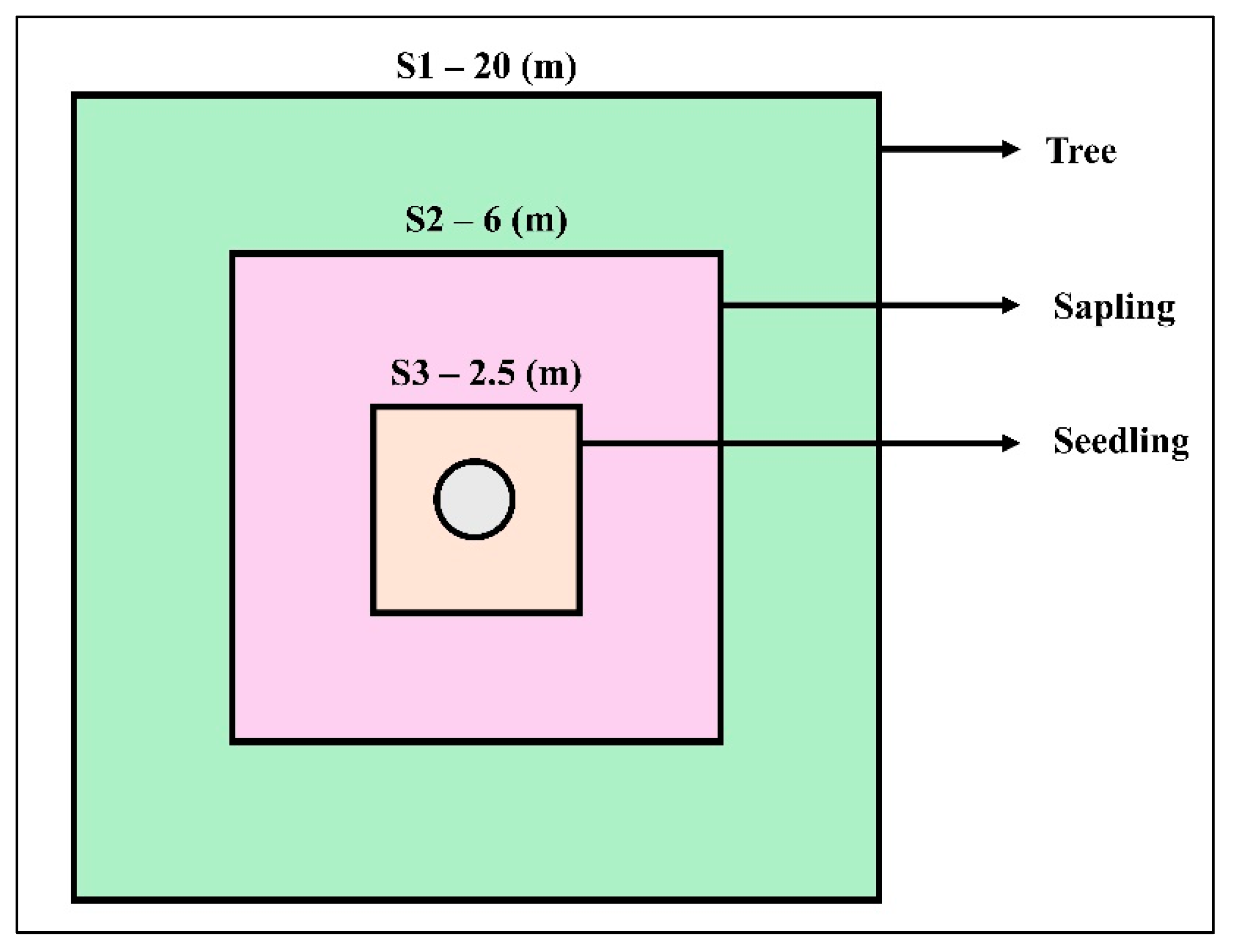
4.2.1. Field Inventory and Sampling
4.2.2. Litter Sampling
4.2.3. Deadwood Sampling
4.2.4. Sampling for Species Diversity
4.3. Calculation of Growing Stock and Biomass
4.4. Soil Measurement
4.5. Tree Species Composition, Dominance and Diversity
4.6. Total Carbon Stocks
4.7. Regeneration Ability
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Xu, K.; Miao, W.; Gao, L.; Ullah, S.; Lyu, Y.; Wang, X. Functional traits and stand factors as strong drivers of radial growth response to hotter drought in a temperate forest of North China. Eur. J. For. Res. 2022, 141, 927–938. [Google Scholar] [CrossRef]
- Dani, R.S.; Divakar, P.K.; Baniya, C.B. Diversity and composition of plants species along elevational gradient: Research trends. Biodivers. Conserv. 2023, 32, 2961–2980. [Google Scholar] [CrossRef]
- Wang, C.; Sheng, Q.; Zhao, R.; Zhu, Z. Differences in the suitable distribution area between northern and southern China landscape plants. Plants 2023, 12, 2710. [Google Scholar] [CrossRef]
- Yang, J.; El-Kassaby, Y.A.; Guan, W. The effect of slope aspect on vegetation attributes in a mountainous dry valley, Southwest China. Sci. Rep. 2020, 10, 16465. [Google Scholar] [CrossRef]
- Umair, M.; Hu, X.; Cheng, Q.; Ali, S.; Ni, J. Distribution patterns of gymnosperm species along elevations on the Qinghai–Tibet Plateau: Effects of climatic seasonality, energy–water, and physical tolerance variables. Plants 2023, 12, 4066. [Google Scholar] [CrossRef]
- Atalay, I.; Efe, R.; Öztürk, M. Ecology and classification of forests in Turkey. Procedia Soc. Behav. Sci. 2014, 120, 788–805. [Google Scholar] [CrossRef]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Naimi, B.; Rahbek, C.; Araújo, M.B. Improvements in reports of species redistribution under climate change are required. Sci. Adv. 2021, 7, eabe1110. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xi, M.; Liu, T.; Wu, X.; Ju, L.; Wang, D. The central role of transcription factors in bridging biotic and abiotic stress responses for plants’ resilience. New Crops 2024, 1, 100005. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Chang, J.G.; Shoshany, M.; Sternberg, M.; Karnieli, A. Assessment of plant species distribution and diversity along a climatic gradient from Mediterranean woodlands to semi-arid shrublands. GISci. Remote Sens. 2021, 58, 929–953. [Google Scholar] [CrossRef]
- Schluter, D.; Pennell, M. Speciation gradients and the distribution of biodiversity. Nature 2017, 546, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.M.; Mishra, A.K.; Tiwari, O.P.; Krishan, R.; Rana, Y.S. Effect of altitudinal gradients on forest structure and composition on ridge tops in Garhwal Himalaya. Energy Ecol. Environ. 2017, 2, 404–417. [Google Scholar] [CrossRef]
- Chauvier, Y.; Thuiller, W.; Brun, P.; Lavergne, S.; Descombes, P.; Karger, D.N.; Renaud, J.; Zimmermann, N.E. Influence of climate, soil, and land cover on plant species distribution in the European Alps. Ecol. Monogra. 2020, 91, e01433. [Google Scholar] [CrossRef]
- Tripathi, S.; Bhadouria, R.; Srivastava, P.; Devi, R.S.; Chaturvedi, R.; Raghubanshi, A.S. Effects of light availability on leaf attributes and seedling growth of four tree species in tropical dry forest. Ecol. Process 2020, 9, 2. [Google Scholar] [CrossRef]
- Kharkwal, G.; Mehrotra, P.; Rawat, Y.; Pangtey, Y. Phytodiversity and growth form in relation to altitudinal gradient in the central Himalayan (Kumaun) region of India. Curr. Sci. 2005, 89, 873–878. [Google Scholar]
- Kahyaoğlu, N.; Kara, Ö.; Güvendi, E. Effects of elevation on the aboveground biomass and carbon stock in the oriental Beech (Fagus orientalis Lipsky) forests of the Sinop region, Turkey. Appl. Ecol. Environ. Res. 2020, 18, 8049–8063. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.; Ohlemüller, R.; Roy, D.B.; Thomas, C. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Jurasinski, G.; Kreyling, J. Upward shift of alpine plants increases floristic similarity of Alpine summits. J. Veg. Sci. 2007, 18, 711–718. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- Sharma, C.; Mishra, A.; Krishan, R.; Prakash, T.O.; Singh, R.Y. Impact of climate on structure and composition of Ridge top forests in Garhwal Himalaya. Taiwania 2016, 61, 61–69. [Google Scholar]
- Sharma, C.M.; Tiwari, O.P.; Rana, Y.S.; Krishan, R.; Mishra, A.K. Elevational behaviour on dominance–diversity, regeneration, biomass and carbon storage in Ridge forests of Garhwal Himalaya, India. For. Ecol. Manag. 2018, 424, 105–120. [Google Scholar] [CrossRef]
- Sekar, K.C.; Thapliyal, N.; Pandey, A.; Joshi, B.; Mukherjee, S.; Bhojak, P.; Bisht, M.; Bhatt, D.; Singh, S.; Bahukhandi, A. Plant species diversity and density patterns along altitude gradient covering high-altitude alpine regions of west Himalaya, India. Geol. Ecol. Landsc. 2024, 8, 559–573. [Google Scholar] [CrossRef]
- Wani, Z.A.; Negi, V.S.; Bhat, J.A.; Satish, K.V.; Kumar, A.; Khan, S.; Dhyani, R.; Siddiqui, S.; Al-Qthanin, R.N.; Pant, S. Elevation, aspect, and habitat heterogeneity determine plant diversity and compositional patterns in the Kashmir Himalaya. Front. For. Glob. Change 2023, 6, 1019277. [Google Scholar]
- Ullah, S.; Liu, W.; Shah, J.A.; Shen, F.; Liao, Y.; Duan, H.; Huang, G.; Wu, J. Effects of canopy nitrogen addition and understory vegetation removal on nitrogen transformations in a subtropical Forest. Forests 2024, 15, 962. [Google Scholar] [CrossRef]
- Gazda, A.; Miścicki, S.; Chwistek, K. Tree species diversity and above-ground biomass of natural temperate forest: Montane versus lowland forest. Dendrobiology 2015, 73, 3–10. [Google Scholar] [CrossRef]
- Nagar, D.K.; Nagar, B.; Nagar, A.L.; Kashyap, S.D. Biomass and carbon density in relation to altitude in moist temperate western Himalayan ecosystem. Indian J. Ecol. 2017, 44, 746–754. [Google Scholar]
- Majumdar, T.; Selvan, T. Carbon storage in trees of urban and peri-urban forests of Agartala, Tripura. J. Adv. Res. Appl. Sci. 2018, 5, 715–731. [Google Scholar]
- Kobler, J.; Zehetgruber, B.; Dirnböck, T.; Jandl, R.; Mirtl, M.; Schindlbacher, A. Effects of aspect and altitude on carbon cycling processes in a temperate mountain forest catchment. Landsc. Ecol. 2019, 34, 325–340. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Snyder, P.K. Global consequences of land use. science 2005, 309, 570–574. [Google Scholar] [CrossRef]
- Negi, V.S.; Pandey, A.; Singh, A.; Bahukhandi, A.; Pharswan, D.S.; Gaira, K.S.; Wani, Z.A.; Bhat, J.A.; Siddiqui, S.; Yassin, H.M. Elevation gradients alter vegetation attributes in mountain ecosystems of eastern Himalaya, India. Front. For. Glob. Change 2024, 7, 1381488. [Google Scholar] [CrossRef]
- Badshah, M.T.; Ahmad, A.; Muneer, M.A.; Rehman, A.U.; Wang, J.; Khan, M.A.; Muhammad, B.; Amir, M.; Meng, J. Evaluation of the forest structure, diversity and biomass carbon potential in the southwest region of Guangxi, China. Appl. Ecol. Environ. Res. 2020, 18, 447–467. [Google Scholar] [CrossRef]
- Anjum, M.N.; Ghanim, A.A.J.; Farid, H.U.; Zaman, M.; Niazi, U.M.; Rahman, S.U.; Alsaiari, M.A.; Irfan, M. Assessment of climate change and its impacts on the flows of a subtropical river basin in the Hindu-Kush mountain, South Asia. Pure Appl. Geophys. 2022, 179, 3841–3857. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Ji, Y.; Yu, M.; Li, X.; Duan, J.; Wang, Y.; Gao, J.; Guo, X. Climate factors affect above–belowground biomass allocation in broad-leaved and coniferous forests by regulating soil nutrients. Plants 2023, 12, 3926. [Google Scholar] [CrossRef]
- Zhang, J.; Zuo, X.; Zhao, X.; Ma, J.; Roldán, E.M. Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland. Sci. Rep. 2020, 10, 9026. [Google Scholar]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Marks, C.O.; Muller Landau, H.C.; Tilman, D. Tree diversity, tree height and environmental harshness in eastern and western north America. Ecol Lett. 2016, 19, 743–751. [Google Scholar] [CrossRef]
- Bhandari, J.; Zhang, Y. Effect of altitude and soil properties on biomass and plant richness in the grasslands of Tibet, China, and Manang district, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Li, Y.Q.; Deng, X.W.; Huang, Z.H.; Xiang, W.H.; Yan, W.D.; Lei, P.F.; Zhou, X.L.; Peng, C.H. Development and evaluation of models for the relationship between tree height and diameter at breast height for Chinese fir plantations in subtropical China. PLoS ONE 2015, 10, 0125118. [Google Scholar] [CrossRef]
- Haq, S.M.; Rashid, I.; Calixto, E.S.; Ali, A.; Kumar, M.; Srivastava, G.; Bussmann, R.W.; Khuroo, A.A. Unravelling patterns of forest carbon stock along a wide elevational gradient in the Himalaya: Implications for climate change mitigation. For. Ecol. Manag. 2022, 521, 120442. [Google Scholar]
- Faria, D.; Mariano-Neto, E.; Martini, A.M.Z.; Ortiz, J.V.; Montingelli, R.; Rosso, S.; Paciencia, M.L.B.; Baumgarten, J. Forest structure in a mosaic of rainforest sites: The effect of fragmentation and recovery after clear cut. For. Ecol. Manag. 2009, 257, 2226–2234. [Google Scholar] [CrossRef]
- Rocha-Santos, L.; Pessoa, M.S.; Cassano, C.R.; Talora, D.C.; Orihuela, R.L.L.; Mariano-Neto, E.; Morante-Filho, J.C.; Faria, D.; Cazetta, E. The shrinkage of a forest: Landscape-scale deforestation leading to overall changes in local forest structure. Biol. Conserv. 2016, 196, 1–9. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Lu, W.; Deng, D. Gibberellin in plant height control: Old player, new story. Plant Cell Rep. 2017, 36, 391–398. [Google Scholar] [CrossRef]
- Wardle, P. An explanation for alpine timberline. N. Z. J. Bot. 1971, 9, 371–402. [Google Scholar] [CrossRef]
- Lippok, D.; Beck, S.G.; Renison, D.; Gallegos, S.C.; Saavedra, F.V.; Hensen, I.; Schleuning, M. Forest recovery of areas deforested by fire increases with elevation in the tropical Andes. For. Ecol. Manag. 2013, 295, 69–76. [Google Scholar] [CrossRef]
- Sheikh, M.A.; Kumar, M.; Todaria, N.P.; Bhat, J.A.; Kumar, A.; Pandey, R. Contribution of Cedrus Deodara forests for climate mitigation along altitudinal gradient in Garhwal Himalaya, India. Mitig. Adapt. Strateg. Glob. Change 2021, 26, 5. [Google Scholar] [CrossRef]
- Lu, D.; Wang, G.G.; Yan, Q.; Gao, T.; Zhu, J. Effects of gap size and within-gap position on seedling growth and biomass allocation: Is the gap partitioning hypothesis applicable to the temperate secondary forest ecosystems in Northeast China? For. Ecol. Manag. 2018, 429, 351–362. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Zu, K.; Lenoir, J.; Fang, J.; Tang, Z.; Shen, Z.; Ji, C.; Wang, Z. Elevational shift in seed plant distributions in China’s mountains over the last 70 years. Glob. Ecol. Biogeogr. 2023, 32, 1098–1112. [Google Scholar] [CrossRef]
- Margreiter, V.; Walde, J.; Erschbamer, B. Competition free gaps are essential for the germination and recruitment of alpine species along an elevation gradient in the European Alps. Alp. Bot. 2021, 131, 135–150. [Google Scholar] [CrossRef]
- He, D.; Peng, D.; Yang, H.; Zhang, X. The response of seedlings and saplings to canopy structure and light in different gaps in a spruce-fir mixed stand in Changbai mountains, China. For. Ecol. Manag. 2023, 546, 121365. [Google Scholar] [CrossRef]
- Tesfaye, G.; Teketay, D.; Fetene, M.; Beck, E. Regeneration of seven indigenous tree species in a dry Afromontane forest, southern Ethiopia. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 135–143. [Google Scholar] [CrossRef]
- Rezende, V.L.; de Miranda, P.L.S.; Meyer, L.; Moreira, C.V.; Linhares, M.F.M.; de Oliveira-Filho, A.T.; Eisenlohr, P.V. Tree species composition and richness along altitudinal gradients as a tool for conservation decisions: The case of Atlantic semideciduous forest. Biodivers. Conserv. 2015, 24, 2149–2163. [Google Scholar] [CrossRef]
- Rana, P.; Koirala, M.; Bhuju, D.R.; Boonchird, C. Population structure of Rhododendron Campanulatum D. Don. and associated tree species along the elevational gradient of Manaslu conservation area, Nepal. J. Inst. Sci. Technol. 2016, 21, 95–102. [Google Scholar] [CrossRef]
- Strong, W.L. Biased richness and evenness relationships within Shannon–Wiener index values. Ecol. Indic. 2016, 67, 703–713. [Google Scholar] [CrossRef]
- Malik, Z.A.; Nautiyal, M.C. Species richness and diversity along the altitudinal gradient in Tungnath, the Himalayan benchmark site of Himadri. Trop. Plant Res. 2016, 3, 396–407. [Google Scholar]
- Dar, J.A.; Sundarapandian, S. Variation of biomass and carbon pools with forest type in temperate forests of Kashmir Himalaya, India. Environ. Monit. Assess. 2015, 187, 55. [Google Scholar] [CrossRef]
- Sinha, S.; Badola, H.K.; Chhetri, B.; Gaira, K.S.; Lepcha, J.; Dhyani, P.P. Effect of altitude and climate in shaping the forest compositions of Singalila national park in Khangchendzonga landscape, eastern Himalaya, India. J. Asia-Pac. Biodivers. 2018, 11, 267–275. [Google Scholar] [CrossRef]
- Van der Wal, A.; de Boer, W.; Smant, W.; Van Veen, J.A. Initial decay of woody fragments in soil is influenced by size, vertical position, nitrogen availability and soil origin. Plant Soil 2007, 301, 189–201. [Google Scholar] [CrossRef]
- Dar, J.A.; Rather, M.Y.; Subashree, K.; Sundarapandian, S.; Khan, M.L. Distribution patterns of tree, understorey, and detritus biomass in coniferous and broad-leaved forests of western Himalaya, India. J. Sustain. For. 2017, 36, 787–805. [Google Scholar] [CrossRef]
- Fravolini, G.; Tognetti, R.; Lombardi, F.; Egli, M.; Ascher-Jenull, J.; Arfaioli, P.; Bardelli, T.; Cherubini, P.; Marchetti, M. Quantifying decay progression of deadwood in Mediterranean mountain forests. For. Ecol. Manag. 2018, 408, 228–237. [Google Scholar] [CrossRef]
- Ullah, S.; Muhammad, B.; Amin, R.; Abbas, H.; Muneer, M.A. Sensitivity of arbuscular mycorrhizal fungi in old-growth forests: Direct effect on growth and soil carbon storage. Appl. Ecol. Env. Res. 2019, 6, 13749–13758. [Google Scholar] [CrossRef]
- Muhammad, B.; Ilahi, T.; Ullah, S.; Wu, X.; Siddique, M.A.; Khan, M.A.; Badshah, M.T.; Jia, Z. Litter decomposition and soil nutrients prince Rupprecht’s (Larix principis-rupprechtii) plantations area in Saihanba, northern China. Appl. Ecol. Env. Res. 2020, 18, 4417–4434. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Harmon, M.E.; Domingo, J.B. Changing temporal patterns of forest carbon stores and net ecosystem carbon balance: The stand to landscape transformation. Landsc. Ecol. 2007, 22, 77–94. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Girardin, C.A.J.; Malhi, Y.; Aragao, L.; Mamani, M.; Huaraca Huasco, W.; Durand, L.; Feeley, K.J.; Rapp, J.; Silva-Espejo, J.E.; Silman, M. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Glob. Change Biol. 2010, 16, 3176–3192. [Google Scholar] [CrossRef]
- Moser, G.; Leuschner, C.; Hertel, D.; Graefe, S.; Soethe, N.; Iost, S. Elevation effects on the carbon budget of tropical mountain forests (S Ecuador): The role of the belowground compartment. Glob. Change Biol. 2011, 17, 2211–2226. [Google Scholar] [CrossRef]
- Dar, A.A.; Parthasarathy, N. Tree species composition, stand structure and distribution patterns across three Kashmir Himalayan forests, India. Écoscience 2022, 29, 311–324. [Google Scholar] [CrossRef]
- Gao, J.F.; Zhang, Y.X. Distributional patterns of species diversity of main plant communities along altitudinal gradient in secondary forest region, Guandi Mountain, China. J. For. Res. 2006, 17, 111–115. [Google Scholar] [CrossRef]
- Dawood, M.; Rahman, A.U.; Rahman, G.; Nadeem, B.; Miandad, M. Geo-statistical analysis of climatic variability and trend detection in the Hindu Kush region, North Pakistan. Environ. Monit. Assess. 2024, 196, 4. [Google Scholar] [CrossRef]
- Sala, O.E.; Stuart Chapin, F.I.I.I.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Haq, S.M.; Calixto, E.S.; Rashid, I.; Srivastava, G.; Khuroo, A.A. Tree diversity, distribution and regeneration in major forest types along an extensive elevational gradient in Indian Himalaya: Implications for sustainable forest management. For. Ecol. Manag. 2022, 506, 119968. [Google Scholar] [CrossRef]
- Khan, I.; Hayat, U.; Lushuang, G.; Khan, F.; Xinyi, H.; Shufan, W. Association of Carbon Pool with Vegetation Composition along the Elevation Gradients in Subtropical Forests in Pakistan. Forests 2024, 15, 1395. [Google Scholar] [CrossRef]
- Sheikh, M.A.; Kumar, M.; Todaria, N.P.; Pandey, R. Biomass and soil carbon along altitudinal gradients in temperate Cedrus deodara forests in central Himalaya, India: Implications for climate change mitigation. Ecol. Indic. 2020, 111, 106025. [Google Scholar] [CrossRef]
- Ali, A. Biomass and Carbon Tables for Major Tree Species of Gilgit Baltistan, Pakistan, Gilgit Baltistan Forests, Wildlife and Environment Department, Gilgit-Baltistan and Pakistan Forest Institute, Peshawar. Biomass Tables of Coniferous Species, Pdf 1. 2015. Available online: https://www.fwegb.gov.pk (accessed on 1 March 2025).
- Ahmad, A.; Nizami, S.M.; Marwat, K.B.; Muhammad, J. Annual accumulation of carbon in the coniferous forest of Dir Kohistan: An inventory based estimate. Pak. J. Bot. 2016, 47, 115–118. [Google Scholar]
- Ali, F.; Khan, N.; Ali, K.; Khan, I. Influence of environmental variables on the distribution of woody species in muslim graveyards of Malakand division, Hindukush range mountains of Pakistan. Pak. J. Bot. 2017, 49, 2357–2366. [Google Scholar]
- Hairiah, K.; Sitompul, S.M.; Van Noordwijk, M.; Palm, C. Methods for Sampling Carbon Stocks Above and Below Ground; ICRAF: Bogor, Indonesia, 2001; pp. 1–23. Available online: https://apps.worldagroforestry.org/downloads/Publications/PDFS/LN01114.PDF (accessed on 1 March 2025).
- Pimentel, D.; Harvey, C.; Resosudarmo, P.; Sinclair, K.; Kurz, D.; McNair, M.; Crist, S.; Shpritz, L.; Fitton, L.; Saffouri, R.; et al. Environmental and economic costs of soil erosion and conservation benefits. Science 1995, 267, 1117–1123. [Google Scholar] [CrossRef]
- Pearson, T.R.; Walker, S.; Brown, S. Sourcebook for Land-Use, Land-Use Change and Forestry Projects; Winrock International and the Bio-Carbon Fund of the World Bank: Arlington, TX, USA, 2005; pp. 19–35. Available online: https://winrock.org/wp-content/uploads/2016/03/Winrock-BioCarbon_Fund_Sourcebook-compressed.pdf (accessed on 1 March 2025).
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communities; University of Illinois Press: Urbana, IL, USA, 1963; p. 117. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Yohannes, H.; Soromessa, T.; Argaw, M. Carbon stock analysis along slope and slope aspect gradient in Gedo Forest: Implicationsfor climate change mitigation. J. Earth Sci. Clim. Change 2015, 6, 6–11. [Google Scholar] [CrossRef]
- Ahmad, A.; Liu, Q.; Nizami, S.M.; Mannan, A.; Saeed, S. Carbon emission from deforestation, forest degradation and wood harvest in the temperate region of Hindukush Himalaya, Pakistan between 1994 and 2016. Land Use Policy 2018, 78, 781–790. [Google Scholar] [CrossRef]
- Akbar, S.; Kharabsheh, B.; Poletti-Hughes, J.; Shah, S.Z.A. Board structure and corporate risk taking in the UK financial sector. Int. Rev. Financ. Anal. 2017, 50, 101–110. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Categories | B | Std. Error | Beta | t | Sig. | Tolerance | VIF |
|---|---|---|---|---|---|---|---|---|
| Location | Usho | −9.456 | 2.82 | −0.035 | −3.354 | <0.001 *** | 0.175 | 5.704 |
| Slope | Gentle | −2.176 | 2.345 | −0.008 | −0.928 | 0.354 NS | 0.235 | 4.251 |
| Moderate | −3.37 | 2.612 | −0.009 | −1.29 | 0.197 NS | 0.391 | 2.555 | |
| Steep | 1.978 | 2.653 | 0.008 | 0.746 | 0.456 NS | 0.168 | 5.948 | |
| Aspect | G2 | −3.387 | 1.313 | −0.013 | −2.579 | 0.01 ** | 0.819 | 1.22 |
| G3 | −4.484 | 1.595 | −0.014 | −2.812 | 0.005 ** | 0.831 | 1.204 | |
| Species | Fir | −65.847 | 1.531 | −0.212 | −43.021 | <0.001 *** | 0.805 | 1.242 |
| Sp | −43.018 | 1.699 | −0.124 | −25.313 | <0.001 *** | 0.815 | 1.227 | |
| BP | −41.91 | 1.454 | −0.142 | −28.829 | <0.001 *** | 0.803 | 1.245 | |
| Elevation | E2 | −6.943 | 2.484 | −0.014 | −2.796 | 0.005 ** | 0.776 | 1.288 |
| E3 | −7.447 | 2.786 | −0.029 | −2.673 | 0.008 ** | 0.167 | 5.995 | |
| E4 | −2.015 | 2.931 | −0.007 | −0.687 | 0.492 NS | 0.19 | 5.267 | |
| E5 | −7.388 | 3.236 | −0.016 | −2.283 | 0.023 * | 0.383 | 2.609 | |
| Basel area | 59.148 | 2.116 | 0.409 | 27.947 | <0.001 *** | 0.091 | 10.946 | |
| Volume | 2.06 | 0.05 | 0.6 | 41.607 | <0.001 *** | 0.094 | 10.656 |
| Elevation | Shannon Index H | Simpson Index D | Species Evenness E | Species Richness S |
|---|---|---|---|---|
| E1 | 1.18 ± 0.01 a | 3.28 ± 0.04 b | 0.91 ± 0.01 a | 3.68 ± 0.02 a |
| E2 | 1.15 ± 0.02 a | 3.18 ± 0.03 a | 0.91 ± 0.01 a | 3.67 ± 0.02 a |
| E3 | 1.09 ± 0.01 b | 2.98 ± 0.02 b | 0.89 ± 0.01 a | 3.67 ± 0.02 a |
| E4 | 1.10 ± 0.02 a | 3.07 ± 0.03 a | 0.87 ± 0.01 b | 3.66 ± 0.02 a |
| E5 | 1.09 ± 0.01 b | 2.94 ± 0.02 a | 0.87 ± 0.01 b | 3.67 ± 0.02 a |
| Adj. R2 | 0.72 | 0.73 | 0.87 | 0.16 |
| F | 11.41 | 11.15 | 27.00 | 2.45 |
| p-value | 0.04 | 0.04 | 0.01 | 0.22 |
| Month | Average Temperature (°C) | Minimum Temperature (°C) | Maximum Temperature (°C) | Precipitation (mm) | Relative Humidity (%) | Rainy Days |
|---|---|---|---|---|---|---|
| January | −13.7 ± 4.32 | −17.2 ± 6.12 | −9.8 ± 2.12 | 111 ± 11.22 | 56 ± 6.32 | 9 ± 1.61 |
| February | −11.2 ± 3.92 | −15 ± 5.32 | −7.3 ± 1.31 | 166 ± 16.83 | 61 ± 6.46 | 10 ± 2.01 |
| March | −7.1 ± 3.11 | −11.5 ± 3.44 | −2.7 ± 0.44 | 196 ± 20.12 | 60 ± 6.16 | 11 ± 2.21 |
| April | −2.4 ± 1.13 | −7.3 ± 1.92 | 1.9 ± 0.24 | 168 ± 17.32 | 59 ± 5.91 | 10 ± 2.11 |
| May | 2.5 ± 1.23 | −3.1 ± 1.02 | 7.2 ± 1.04 | 102 ± 10.72 | 63 ± 6.56 | 8 ± 1.31 |
| June | 8.1 ± 2.12 | 1.3 ± 0.32 | 13.3 ± 3.12 | 88 ± 9.12 | 64 ± 6.22 | 7 ± 1.01 |
| July | 12 ± 3.78 | 6.1 ± 2.67 | 16.9 ± 5.02 | 127 ± 12.52 | 74 ± 6.13 | 17 ± 2.16 |
| August | 11.5 ± 3.32 | 6.1 ± 2.71 | 16.2 ± 4.98 | 110 ± 11.32 | 77 ± 7.01 | 12 ± 1.98 |
| September | 7.4 ± 3.32 | 1.4 ± 0.35 | 12.6 ± 3.12 | 78 ± 9.99 | 71 ± 7.12 | 7 ± 1.01 |
| October | −0.1 ± 0.12 | −6.3 ± 3.31 | 5.6 ± 2.92 | 68 ± 9.78 | 64 ± 6.16 | 8 ± 1.21 |
| November | −7.9 ± 1.22 | −12.5 ± 4.62 | −3.2 ± 1.12 | 70 ± 10.22 | 59 ± 5.96 | 6 ± 0.98 |
| December | −12.5 ± 35 | −16.1 ± 5.22 | −8.6 ± 3.02 | 77 ± 10.32 | 55 ± 5.11 | 6 ± 0.97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muhammad, B.; Hayat, U.; Gopakumar, L.; Xiong, S.; Ali, J.; Badshah, M.T.; Ullah, S.; Rehman, A.U.; Yin, Q.; Jia, Z. Altitudinal Variations in Coniferous Vegetation and Soil Carbon Storage in Kalam Temperate Forest, Pakistan. Plants 2025, 14, 1534. https://doi.org/10.3390/plants14101534
Muhammad B, Hayat U, Gopakumar L, Xiong S, Ali J, Badshah MT, Ullah S, Rehman AU, Yin Q, Jia Z. Altitudinal Variations in Coniferous Vegetation and Soil Carbon Storage in Kalam Temperate Forest, Pakistan. Plants. 2025; 14(10):1534. https://doi.org/10.3390/plants14101534
Chicago/Turabian StyleMuhammad, Bilal, Umer Hayat, Lakshmi Gopakumar, Shuangjiang Xiong, Jamshid Ali, Muhammad Tariq Badshah, Saif Ullah, Arif UR Rehman, Qun Yin, and Zhongkui Jia. 2025. "Altitudinal Variations in Coniferous Vegetation and Soil Carbon Storage in Kalam Temperate Forest, Pakistan" Plants 14, no. 10: 1534. https://doi.org/10.3390/plants14101534
APA StyleMuhammad, B., Hayat, U., Gopakumar, L., Xiong, S., Ali, J., Badshah, M. T., Ullah, S., Rehman, A. U., Yin, Q., & Jia, Z. (2025). Altitudinal Variations in Coniferous Vegetation and Soil Carbon Storage in Kalam Temperate Forest, Pakistan. Plants, 14(10), 1534. https://doi.org/10.3390/plants14101534