
Genome-Wide Identification, Characterization, and Expression Analysis of VQ Gene Family in Salix suchowensis Under Abiotic Stresses and Hormone Treatments
Abstract
1. Introduction
2. Results
2.1. Identification of the SsVQ Genes and Analysis of Physicochemical Properties
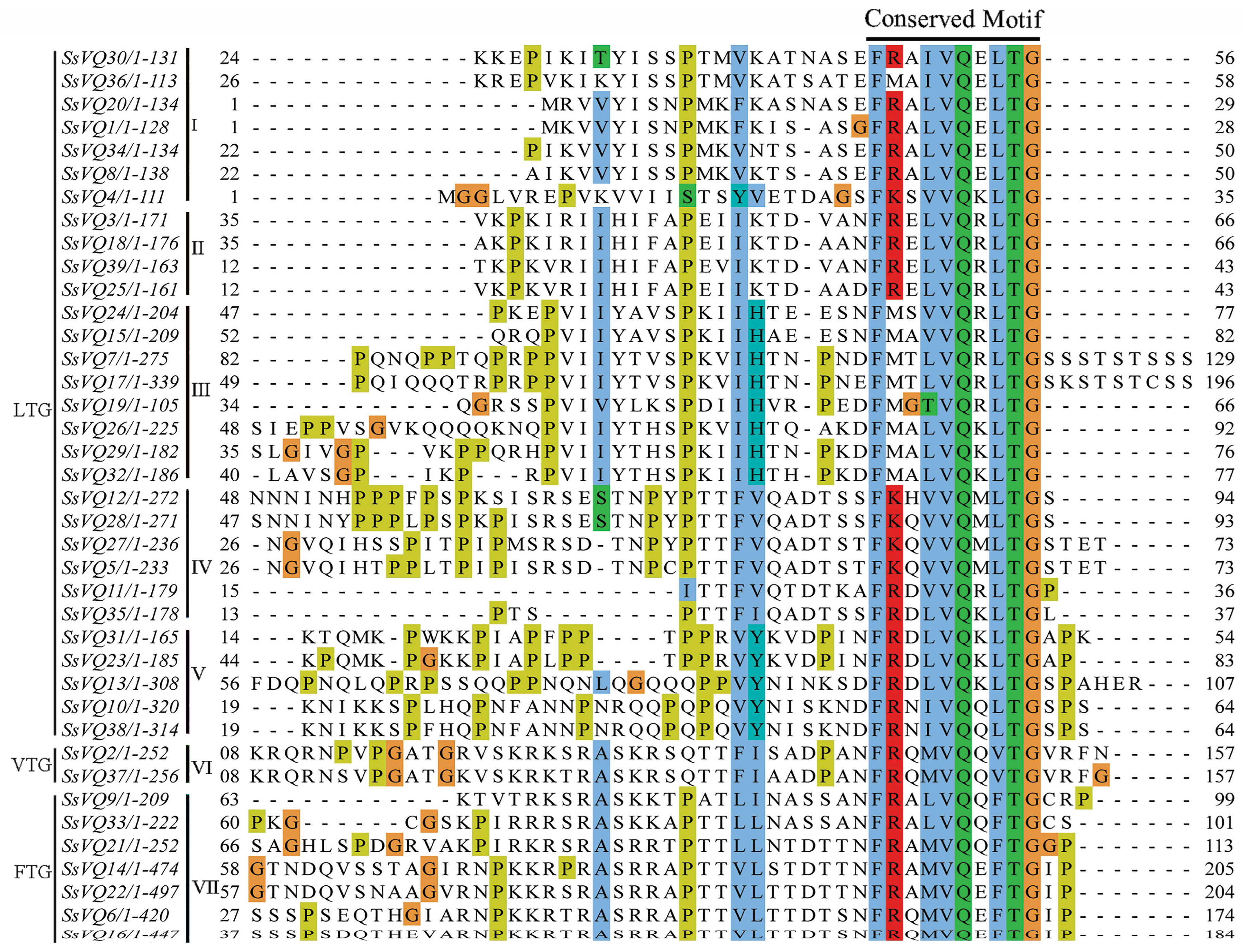
2.2. Phylogenetic Analysis of VQ Genes
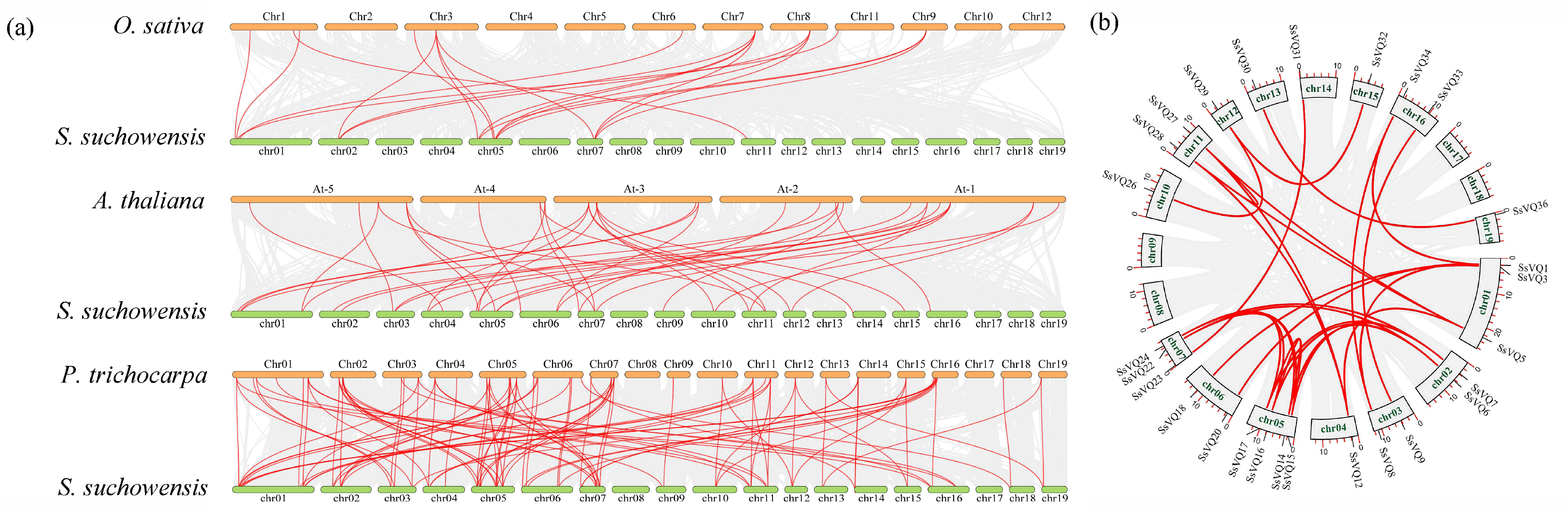
2.3. Duplication Events of VQ Genes
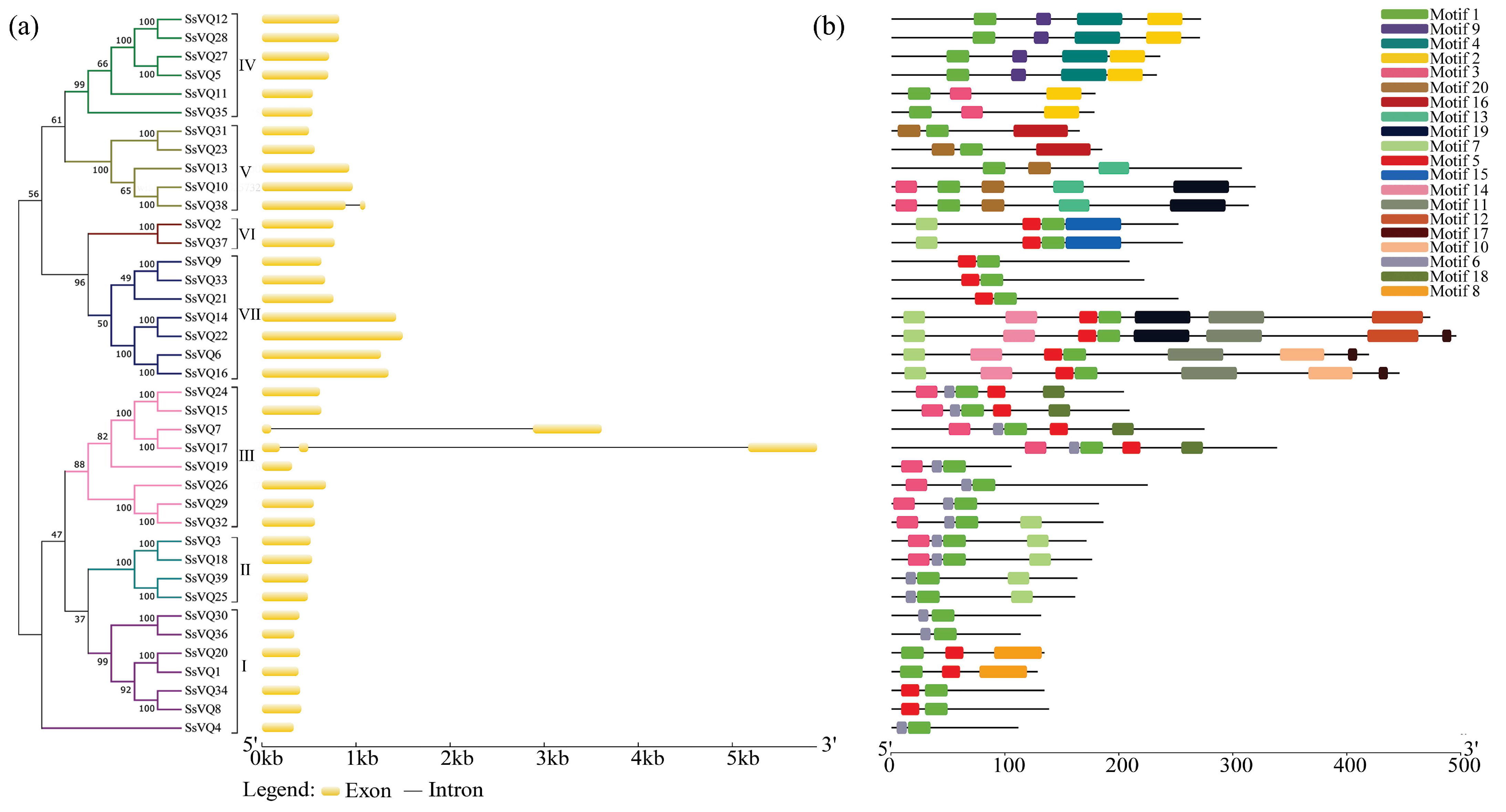
2.4. The Structure and Motif Analysis of the SsVQ Genes
2.5. Identification of Cis-Elements in the Promoters of SsVQ Genes
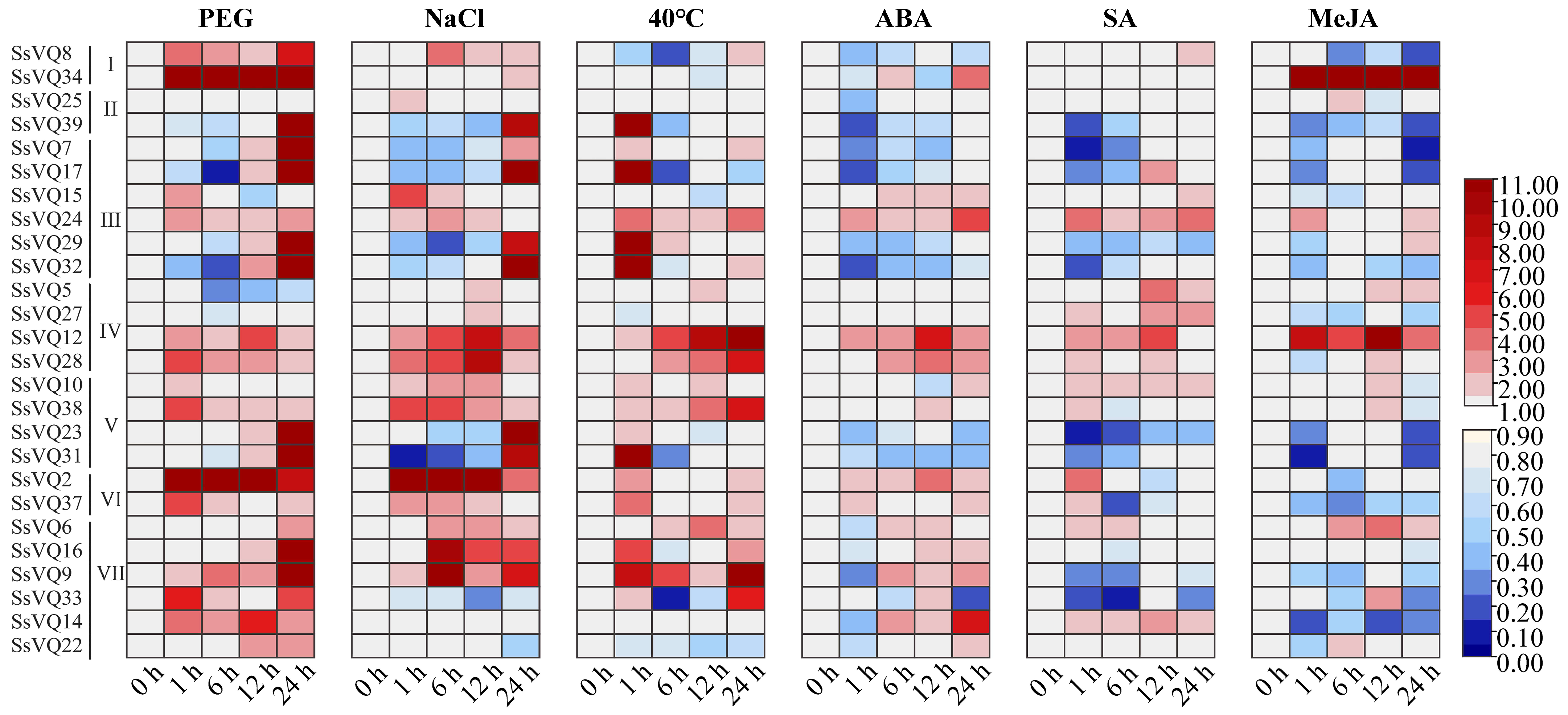
2.6. Expression Patterns of the SsVQ Genes in Response to Abiotic Stress and Exogenous Hormone
2.7. Co-Expression Networks of the SsVQ Genes
3. Discussion
4. Materials and Methods
4.1. Identification and Analysis of the SsVQ Genes
4.2. Multiple Alignment and Phylogenetic Tree Construction of the SsVQs
4.3. Collinearity Analysis of the SsVQ Genes
4.4. Conserved Motifs and Structure Analysis of the SsVQs
4.5. Identification of Cis-Elements in the Promoters of SsVQ Genes
4.6. Plant Materials, Growth Conditions, and Stress Treatments
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Co-Expression Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Li, Y.; Jia, H.X.; Li, J.B.; Huang, J.; Lu, M.Z.; Hu, J.J. The heat shock factor gene family in Salix suchowensis: A genome-wide survey and expression profiling during development and abiotic stresses. Front. Plant Sci. 2015, 6, 748. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Hu, Q.; Cai, Q.; Feng, K.; Ye, N.; Tuskan, G.A.; Milne, R.; Chen, Y.; Wan, Z.; Wang, Z.; et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 2014, 24, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Ye, N.; Yin, T. Detecting the Candidate Gender Determinants by Bioinformatic Prediction of miRNAs and Their Targets from Transcriptome Sequences of the Male and Female Flowers in Salix suchowensis. Biomed Res. Int. 2017, 2017, 9614596. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Chen, Y.; Hou, J.; Yang, Y.; Yin, T. Aux/IAA and ARF Gene Families in Salix suchowensis: Identification, Evolution, and Dynamic Transcriptome Profiling During the Plant Growth Process. Front. Plant Sci. 2021, 12, 666310. [Google Scholar] [CrossRef]
- Ren, Y.; He, Z.; Liu, P.; Traw, B.; Sun, S.; Tian, D.; Yang, S.; Jia, Y.; Wang, L. Somatic Mutation Analysis in Salix suchowensis Reveals Early-Segregated Cell Lineages. Mol. Biol. Evol. 2021, 38, 5292–5308. [Google Scholar] [CrossRef]
- Wang, P.; Guo, J.; Zhou, J.; Wang, Y. Characterization of the complete chloroplast genome and development of molecular markers of Salix. Sci. Rep. 2024, 14, 28528. [Google Scholar] [CrossRef]
- He, L.; Liao, S.; Applequist, W.; Chen, S. The valid publication of Salix suchowensis (Salicaceae). PhytoKeys 2019, 131, 27–35. [Google Scholar] [CrossRef]
- Chi, Y.; Yang, Y.; Zhou, Y.; Zhou, J.; Fan, B.; Yu, J.Q.; Chen, Z. Protein-protein interactions in the regulation of WRKY transcription factors. Mol. Plant 2013, 6, 287–300. [Google Scholar] [CrossRef]
- Xu, K.; Wang, P. Genome-wide identification and expression analysis of the VQ gene family in Cucurbita pepo L. PeerJ 2022, 10, e12827. [Google Scholar] [CrossRef]
- Guo, J.; Chen, J.; Yang, J.; Yu, Y.; Yang, Y.; Wang, W. Identification, characterization and expression analysis of the VQ motif-containing gene family in tea plant (Camellia sinensis). BMC Genom. 2018, 19, 710. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, Y.; Yang, Y.; Chi, Y.J.; Zhou, J.; Chen, J.Y.; Wang, F.; Fan, B.; Shi, K.; Zhou, Y.H.; et al. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012, 159, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Kwon, S.I.; Choi, C.; Lee, H.; Ahn, I.; Park, S.R.; Bae, S.C.; Lee, S.C.; Hwang, D.J. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene 2013, 529, 208–214. [Google Scholar] [CrossRef]
- Chu, W.; Liu, B.; Wang, Y.; Pan, F.; Chen, Z.; Yan, H.; Xiang, Y. Genome-wide analysis of poplar VQ gene family and expression profiling under PEG, NaCl, and SA treatments. Tree Genet. Genomes 2016, 12, 1–17. [Google Scholar] [CrossRef]
- Jing, Y.; Lin, R. The VQ Motif-Containing Protein Family of Plant-Specific Transcriptional Regulators. Plant Physiol. 2015, 169, 371–378. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, X.; Fu, Y.; Wang, H.; Yu, C.; Chu, J.; Jiang, B.; Zhu, J. Genome-wide identification and expression analysis of VQ gene family under abiotic stress in Coix lacryma-jobi L. BMC Plant Biol. 2023, 23, 327. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Qian, Y.; Ren, Y.; Guan, Y.; Wu, X.; Ge, C.; Ding, H. The role of plant-specific VQ motif-containing proteins: An ever-thickening plot. Plant Physiol. Biochem. 2021, 159, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jing, Y.; Li, J.; Xu, G.; Lin, R. Arabidopsis VQ MOTIF-CONTAINING PROTEIN29 represses seedling deetiolation by interacting with PHYTOCHROME-INTERACTING FACTOR1. Plant Physiol. 2014, 164, 2068–2080. [Google Scholar] [CrossRef]
- Yang, M.; Liu, Z.; Yu, Y.; Yang, M.; Guo, L.; Han, X.; Ma, X.; Huang, Z.; Gao, Q. Genome-wide identification of the valine-glutamine motif containing gene family and the role of VQ25-1 in pollen germination in Brassica oleracea. Genes Genom. 2023, 45, 921–934. [Google Scholar] [CrossRef]
- Pan, J.; Wang, H.; Hu, Y.; Yu, D. Arabidopsis VQ18 and VQ26 proteins interact with ABI5 transcription factor to negatively modulate ABA response during seed germination. Plant J. 2018, 95, 529–544. [Google Scholar] [CrossRef]
- Wang, H.; Hu, Y.; Pan, J.; Yu, D. Arabidopsis VQ motif-containing proteins VQ12 and VQ29 negatively modulate basal defense against Botrytis cinerea. Sci. Rep. 2015, 5, 14185. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, Y.; Xiong, X.; Li, H.; Zhang, X.; Song, Y.; Zhang, X.; Min, D. The wheat VQ motif-containing protein TaVQ4-D positively regulates drought tolerance in transgenic plants. J. Exp. Bot. 2023, 74, 5591–5605. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, Y.; Xiong, R.; Gao, Y.; Yan, H.; Xiang, Y. A Moso bamboo gene VQ28 confers salt tolerance to transgenic Arabidopsis plants. Planta 2020, 251, 99. [Google Scholar] [CrossRef]
- Chen, J.; Wang, H.; Li, Y.; Pan, J.; Hu, Y.; Yu, D. Arabidopsis VQ10 interacts with WRKY8 to modulate basal defense against Botrytis cinerea. J. Integr. Plant Biol. 2018, 60, 956–969. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Li, X.; Ma, Z.; Lv, Y.; Hu, Y.; Yu, D. Arabidopsis WRKY2 and WRKY34 transcription factors interact with VQ20 protein to modulate pollen development and function. Plant J. 2017, 91, 962–976. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Ji, Y.; Jing, Y.; Li, L.; Chen, Y.; Wang, R.; Zhang, H.; Yu, D.; Chen, L. Arabidopsis SIGMA FACTOR BINDING PROTEIN1 (SIB1) and SIB2 inhibit WRKY75 function in abscisic acid-mediated leaf senescence and seed germination. J. Exp. Bot. 2022, 73, 182–196. [Google Scholar] [CrossRef]
- Hao, Z.; Tian, J.; Fang, H.; Fang, L.; Xu, X.; He, F.; Li, S.; Xie, W.; Du, Q.; You, X.; et al. A VQ-motif-containing protein fine-tunes rice immunity and growth by a hierarchical regulatory mechanism. Cell Rep. 2022, 40, 111235. [Google Scholar] [CrossRef]
- Meng, X.; Lu, M.; Xia, Z.; Li, H.; Liu, D.; Li, K.; Yin, P.; Wang, G.; Zhou, C. Wheat VQ Motif-Containing Protein VQ25-A Facilitates Leaf Senescence via the Abscisic Acid Pathway. Int. J. Mol. Sci. 2023, 24, 13839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Xu, R.R.; Liu, Y.; You, C.X.; An, J.P. MdVQ10 promotes wound-triggered leaf senescence in association with MdWRKY75 and undergoes antagonistic modulation of MdCML15 and MdJAZs in apple. Plant J. 2023, 115, 1599–1618. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, M.; Li, Y.; Zhang, J.; Su, H.; Cao, M.; Liu, Z.; Zhang, X.; Zhao, B.; Guo, Y.D.; et al. The transcription factor SlWRKY37 positively regulates jasmonic acid- and dark-induced leaf senescence in tomato. J. Exp. Bot. 2022, 73, 6207–6225. [Google Scholar] [CrossRef]
- Gayubas, B.; Castillo, M.C.; Ramos, S.; Leon, J. Enhanced meristem development, tolerance to oxidative stress and hyposensitivity to nitric oxide in the hypermorphic vq10-H mutant in AtVQ10 gene. Plant Cell Environ. 2023, 46, 3445–3463. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, J.; Li, J.; Sun, P.; Zhang, Y.; Xin, X.; Lu, M.; Hu, J. Genome-wide transcriptomic analysis of a desert willow, Salix psammophila, reveals the function of hub genes SpMDP1 and SpWRKY33 in drought tolerance. BMC Plant Biol. 2019, 19, 356. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, X.; Xiao, J.; Wang, S. Comprehensive analysis of VQ motif-containing gene expression in rice defense responses to three pathogens. Plant Cell Rep. 2014, 33, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Vannozzi, A.; Wang, G.; Zhong, Y.; Corso, M.; Cavallini, E.; Cheng, Z.M. A comprehensive survey of the grapevine VQ gene family and its transcriptional correlation with WRKY proteins. Front. Plant Sci. 2015, 6, 417. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-wide analysis of VQ motif-containing proteins in Moso bamboo (Phyllostachys edulis). Planta 2017, 246, 165–181. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Sevugan, M.; Ramachandran, S. Valine-glutamine (VQ) motif coding genes are ancient and non-plant-specific with comprehensive expression regulation by various biotic and abiotic stresses. BMC Genom. 2018, 19, 342. [Google Scholar] [CrossRef]
- Song, W.; Zhao, H.; Zhang, X.; Lei, L.; Lai, J. Genome-Wide Identification of VQ Motif-Containing Proteins and their Expression Profiles Under Abiotic Stresses in Maize. Front. Plant Sci. 2015, 6, 1177. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Luo, R.; Liu, X.; Hou, Z.; Pei, W.; Zhu, W.; Cui, H. Characterization and the comprehensive expression analysis of tobacco valine-glutamine genes in response to trichomes development and stress tolerance. Bot. Stud. 2023, 64, 18. [Google Scholar] [CrossRef]
- Zheng, J.; Li, H.; Guo, Z.; Zhuang, X.; Huang, W.; Mao, C.; Feng, H.; Zhang, Y.; Wu, H.; Zhou, Y. Comprehensive Identification and Expression Profiling of the VQ Motif-Containing Gene Family in Brassica juncea. Biology 2022, 11, 1814. [Google Scholar] [CrossRef]
- Perruc, E.; Charpenteau, M.; Ramirez, B.C.; Jauneau, A.; Galaud, J.P.; Ranjeva, R.; Ranty, B. A novel calmodulin-binding protein functions as a negative regulator of osmotic stress tolerance in Arabidopsis thaliana seedlings. Plant J. 2004, 38, 410–420. [Google Scholar] [CrossRef]
- Zhai, M.; Ao, Z.; Qu, H.; Guo, D. Overexpression of the potato VQ31 enhances salt tolerance in Arabidopsis. Front. Plant Sci. 2024, 15, 1347861. [Google Scholar] [CrossRef]
- Weyhe, M.; Eschen-Lippold, L.; Pecher, P.; Scheel, D.; Lee, J. Menage a trois: The complex relationships between mitogen-activated protein kinases, WRKY transcription factors, and VQ-motif-containing proteins. Plant Signal. Behav. 2014, 9, e29519. [Google Scholar] [CrossRef] [PubMed]
- Pecher, P.; Eschen-Lippold, L.; Herklotz, S.; Kuhle, K.; Naumann, K.; Bethke, G.; Uhrig, J.; Weyhe, M.; Scheel, D.; Lee, J. The Arabidopsis thaliana mitogen-activated protein kinases MPK3 and MPK6 target a subclass of V‘Q-motif’-containing proteins to regulate immune responses. New Phytol. 2014, 203, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Li, Y.; Wang, F.; Cheng, Y.; Fan, B.; Yu, J.Q.; Chen, Z. Arabidopsis sigma factor binding proteins are activators of the WRKY33 transcription factor in plant defense. Plant Cell 2011, 23, 3824–3841. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, M.; Liu, Y.; He, Q.; Chai, M.; Liu, L.; Chen, F.; Huang, Y.; Yan, M.; Zhao, H.; et al. Genome-Wide Classification and Evolutionary and Functional Analyses of the VQ Family. Trop. Plant Biol. 2019, 12, 117–131. [Google Scholar] [CrossRef]
- Qu, Y.; Bi, C.; He, B.; Ye, N.; Yin, T.; Xu, L.A. Genome-wide identification and characterization of the MADS-box gene family in Salix suchowensis. PeerJ 2019, 7, e8019. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Han, X.; Zhang, J.; Sun, P.; Lu, M.; Hu, J. Selection of Reliable Reference Genes for Gene Expression Analysis under Abiotic Stresses in the Desert Biomass Willow, Salix psammophila. Front. Plant Sci. 2016, 7, 1505. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Jia, H.; Yue, Z.; Lu, M.; Xin, X.; Hu, J. Genome-Wide Characterization of the sHsp Gene Family in Salix suchowensis Reveals Its Functions under Different Abiotic Stresses. Int. J. Mol. Sci. 2018, 19, 3246. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequenced ID | Protein Length (aa) | Molecular Weight (Da) | CDS Length (bp) | Exons | pI | Chromosome Location | Subcellular Location |
|---|---|---|---|---|---|---|---|---|
| SsVQ1 | willow_GLEAN_10003577 | 128 | 14,077.71 | 387 | 1 | 4.4 | chr1:1956562/1956948 | Chloroplast |
| SsVQ2 | willow_GLEAN_10006995 | 252 | 20,306.29 | 759 | 1 | 8.99 | chr1:1542044/1542802 | Nucleus |
| SsVQ3 | willow_GLEAN_10010342 | 171 | 19,237.06 | 516 | 1 | 8.37 | chr1:2484352/2484867 | Nucleus |
| SsVQ4 | willow_GLEAN_10017041 | 111 | 12,047.6 | 336 | 1 | 4.82 | chr1:21350745/21351080 | Nucleus |
| SsVQ5 | willow_GLEAN_10017545 | 233 | 25,421.43 | 702 | 1 | 9.61 | chr1:22790432/22791133 | Nucleus |
| SsVQ6 | willow_GLEAN_10020919 | 420 | 45,266.98 | 1263 | 1 | 7.06 | chr2:6303101/6304363 | Nucleus |
| SsVQ7 | willow_GLEAN_10022200 | 275 | 30,337.21 | 828 | 2 | 9.46 | chr2:3767449/3771058 | Nucleus |
| SsVQ8 | willow_GLEAN_10013497 | 138 | 15,010.84 | 417 | 1 | 5.76 | chr3:11005186/11005602 | Nucleus |
| SsVQ9 | willow_GLEAN_10022134 | 209 | 22,699.94 | 630 | 1 | 6.28 | chr3:4156214/4156843 | Plasma membrane |
| SsVQ10 | willow_GLEAN_10025732 | 320 | 34,874.73 | 963 | 1 | 10.43 | chr3:5293007/5293969 | Mitochondrion |
| SsVQ11 | willow_GLEAN_10002711 | 179 | 19,597.23 | 540 | 1 | 9.48 | chr4:7211068/7211607 | Chloroplast |
| SsVQ12 | willow_GLEAN_10017371 | 272 | 29,498.83 | 819 | 1 | 9.32 | chr4:1785239/1786057 | Nucleus |
| SsVQ13 | willow_GLEAN_10005030 | 308 | 33,697.03 | 927 | 1 | 10.4 | chr5:7911241/7912167 | Cytosol |
| SsVQ14 | willow_GLEAN_10006772 | 474 | 51,061.87 | 1425 | 1 | 7.33 | chr5:2808863/2810287 | Nucleus |
| SsVQ15 | willow_GLEAN_10008301 | 209 | 22,596.7 | 630 | 1 | 6.18 | chr5:1833356/1833985 | Nucleus |
| SsVQ16 | willow_GLEAN_10012082 | 447 | 47,594 | 1344 | 1 | 6.15 | chr5:8238824/8240167 | Nucleus |
| SsVQ17 | willow_GLEAN_10015916 | 339 | 37,735.18 | 1020 | 3 | 9.65 | chr5:10092888/10098786 | Nucleus |
| SsVQ18 | willow_GLEAN_10025922 | 176 | 19,198.05 | 531 | 1 | 9.05 | chr6:11654744/11655274 | Nucleus |
| SsVQ19 | willow_GLEAN_10025976 | 105 | 11,418.14 | 318 | 1 | 9.84 | chr6:12133882/12134199 | Nucleus |
| SsVQ20 | willow_GLEAN_10027065 | 134 | 14,522.89 | 405 | 1 | 4.08 | chr6:1548291/1548695 | Nucleus |
| SsVQ21 | willow_GLEAN_10027239 | 252 | 26,918.68 | 759 | 1 | 9.85 | chr6:293114/293872 | Nucleus |
| SsVQ22 | willow_GLEAN_10007056 | 497 | 53,367.44 | 1494 | 1 | 6.55 | chr7:5496219/5497712 | Nucleus |
| SsVQ23 | willow_GLEAN_10012598 | 185 | 20,306.29 | 558 | 1 | 8.99 | chr7:315837/316394 | Cytosol |
| SsVQ24 | willow_GLEAN_10013987 | 204 | 22,118.2 | 615 | 1 | 6.51 | chr7:6606481/6607095 | Cytosol |
| SsVQ25 | willow_GLEAN_10022840 | 161 | 18,067.6 | 486 | 1 | 7.71 | chr9:2426972/2427457 | Nucleus |
| SsVQ26 | willow_GLEAN_10021307 | 225 | 24,179.1 | 678 | 1 | 9.44 | chr10:7351516/7352193 | Nucleus |
| SsVQ27 | willow_GLEAN_10007472 | 236 | 25,617.57 | 711 | 1 | 9.77 | chr11:7876010/7876720 | Nucleus |
| SsVQ28 | willow_GLEAN_10010863 | 271 | 29,235.6 | 816 | 1 | 9.74 | chr11:2409757/2410572 | Nucleus |
| SsVQ29 | willow_GLEAN_10019827 | 182 | 20,184.95 | 549 | 1 | 9.44 | chr12:2379475/2380023 | Nucleus |
| SsVQ30 | willow_GLEAN_10012169 | 131 | 14,800.56 | 396 | 1 | 5.25 | chr13:2567846/2568241 | Chloroplast |
| SsVQ31 | willow_GLEAN_10008946 | 165 | 18,484.41 | 498 | 1 | 9.01 | chr14:171647/172144 | Nucleus |
| SsVQ32 | willow_GLEAN_10002847 | 186 | 20,439.05 | 561 | 1 | 7.68 | chr15:4289624/4290184 | Nucleus |
| SsVQ33 | willow_GLEAN_10017873 | 222 | 23,897.12 | 669 | 1 | 4.74 | chr16:9603289/9603957 | Cytosol |
| SsVQ34 | willow_GLEAN_10023265 | 134 | 14,840.49 | 405 | 1 | 5.94 | chr16:1384684/1385088 | Cytosol |
| SsVQ35 | willow_GLEAN_10006845 | 178 | 19,382.71 | 537 | 1 | 7.8 | chr18:380560/381096 | Nucleus |
| SsVQ36 | willow_GLEAN_10004237 | 113 | 12,580.16 | 342 | 1 | 5.14 | chr19:584761/585102 | Nucleus |
| SsVQ37 | willow_GLEAN_10001773 | 256 | 27,397.3 | 771 | 1 | 5.93 | scaffold01123:3188/3958 | Nucleus |
| SsVQ38 | willow_GLEAN_10001601 | 314 | 34,011.01 | 945 | 2 | 10.39 | scaffold01654:1195/2291 | Nucleus |
| SsVQ39 | willow_GLEAN_10001403 | 163 | 17,683.21 | 492 | 1 | 9.17 | scaffold02338:2728/3219 | Nucleus |
| Treatments | The Co-Expressed Number of N a | The Co-Expressed Ratio of N a | The Co-Expressed Number of P b | The Co-Expressed Ratio of P b | Co-Expressed Paralogous Pairs |
|---|---|---|---|---|---|
| PEG | 82 | 26.28 | 7 | 53.85 | SsVQ2-SsVQ37; SsVQ6-SsVQ16; SsVQ7-SsVQ17; SsVQ8-SsVQ34; SsVQ10-SsVQ38; SsVQ23-SsVQ31; SsVQ29-SsVQ32; |
| NaCl | 55 | 17.63 | 5 | 38.46 | SsVQ2-SsVQ37; SsVQ7-SsVQ17; SsVQ12-SsVQ28; SsVQ23-SsVQ31; SsVQ29-SsVQ32; |
| 40 °C | 56 | 17.95 | 7 | 53.85 | SsVQ2-SsVQ37; SsVQ5-SsVQ27; SsVQ7-SsVQ17; SsVQ9-SsVQ33; SsVQ12-SsVQ28; SsVQ23-SsVQ31; SsVQ29-SsVQ32; |
| ABA | 35 | 11.22 | 4 | 30.77 | SsVQ7-SsVQ17; SsVQ12-SsVQ28; SsVQ25-SsVQ39; SsVQ29-SsVQ32; |
| SA | 41 | 13.14 | 4 | 30.77 | SsVQ2-SsVQ37; SsVQ7-SsVQ17; SsVQ9-SsVQ33; SsVQ12-SsVQ28; |
| MeJA | 32 | 10.26 | 4 | 30.77 | SsVQ7-SsVQ17; SsVQ9-SsVQ33; SsVQ10-SsVQ38; SsVQ23-SsVQ31; |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wang, Y.; Wang, Y.; Zhu, J.; Chen, L.; Yan, X.; Yu, C.; Jiang, B. Genome-Wide Identification, Characterization, and Expression Analysis of VQ Gene Family in Salix suchowensis Under Abiotic Stresses and Hormone Treatments. Plants 2025, 14, 1431. https://doi.org/10.3390/plants14101431
Wang H, Wang Y, Wang Y, Zhu J, Chen L, Yan X, Yu C, Jiang B. Genome-Wide Identification, Characterization, and Expression Analysis of VQ Gene Family in Salix suchowensis Under Abiotic Stresses and Hormone Treatments. Plants. 2025; 14(10):1431. https://doi.org/10.3390/plants14101431
Chicago/Turabian StyleWang, Hongjuan, Yujiao Wang, Yongle Wang, Jiabao Zhu, Lei Chen, Xiaoming Yan, Chun Yu, and Benli Jiang. 2025. "Genome-Wide Identification, Characterization, and Expression Analysis of VQ Gene Family in Salix suchowensis Under Abiotic Stresses and Hormone Treatments" Plants 14, no. 10: 1431. https://doi.org/10.3390/plants14101431
APA StyleWang, H., Wang, Y., Wang, Y., Zhu, J., Chen, L., Yan, X., Yu, C., & Jiang, B. (2025). Genome-Wide Identification, Characterization, and Expression Analysis of VQ Gene Family in Salix suchowensis Under Abiotic Stresses and Hormone Treatments. Plants, 14(10), 1431. https://doi.org/10.3390/plants14101431