
Assessing Vegetation Canopy Growth Variations in Northeast China


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (1)
- What is the spatiotemporal distribution of monthly canopy development, maturation, and senescence rate changes under the background of climate change?
- (2)
- What are the differences in canopy development, maturation, and senescence rate changes among different vegetation types?
- (3)
- What are the response characteristics of canopy development, maturation, and senescence rate changes in different vegetation types to climate change?
2. Results
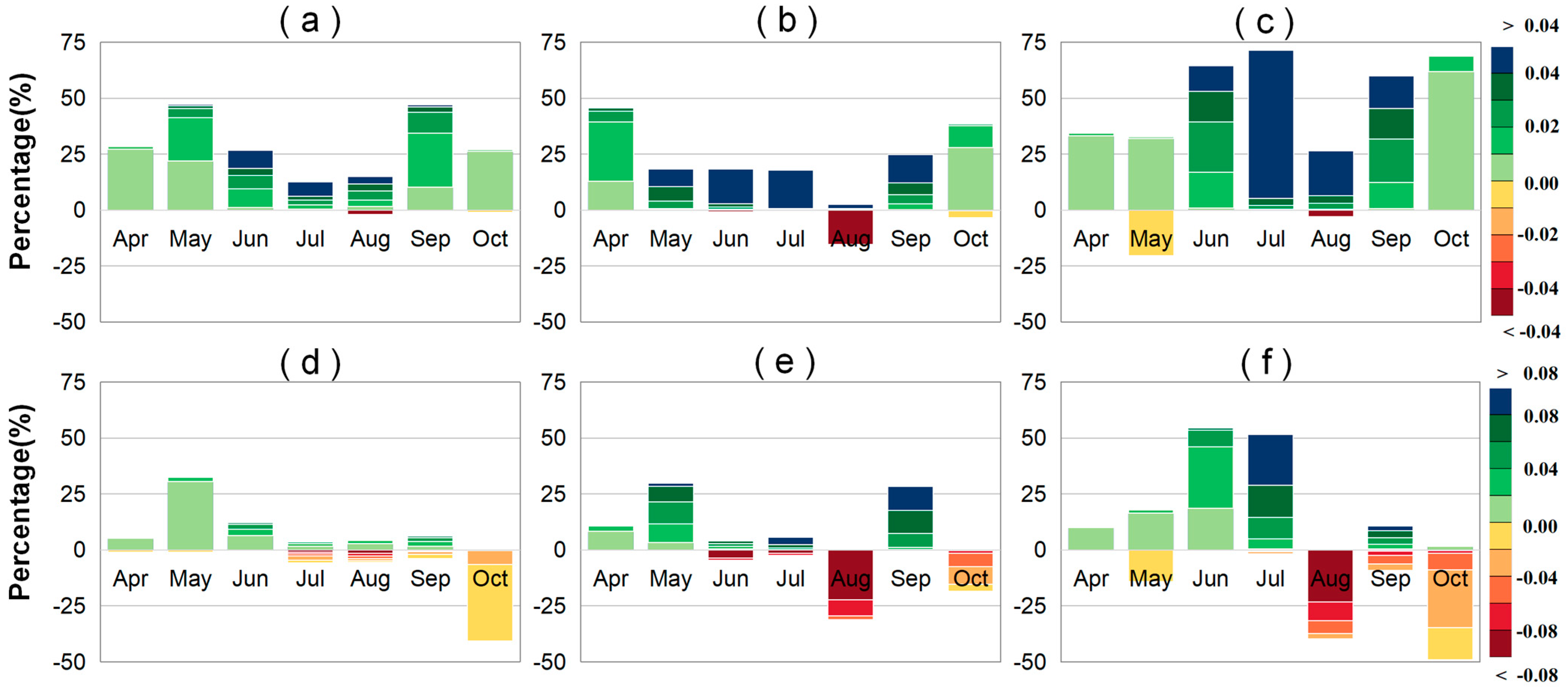
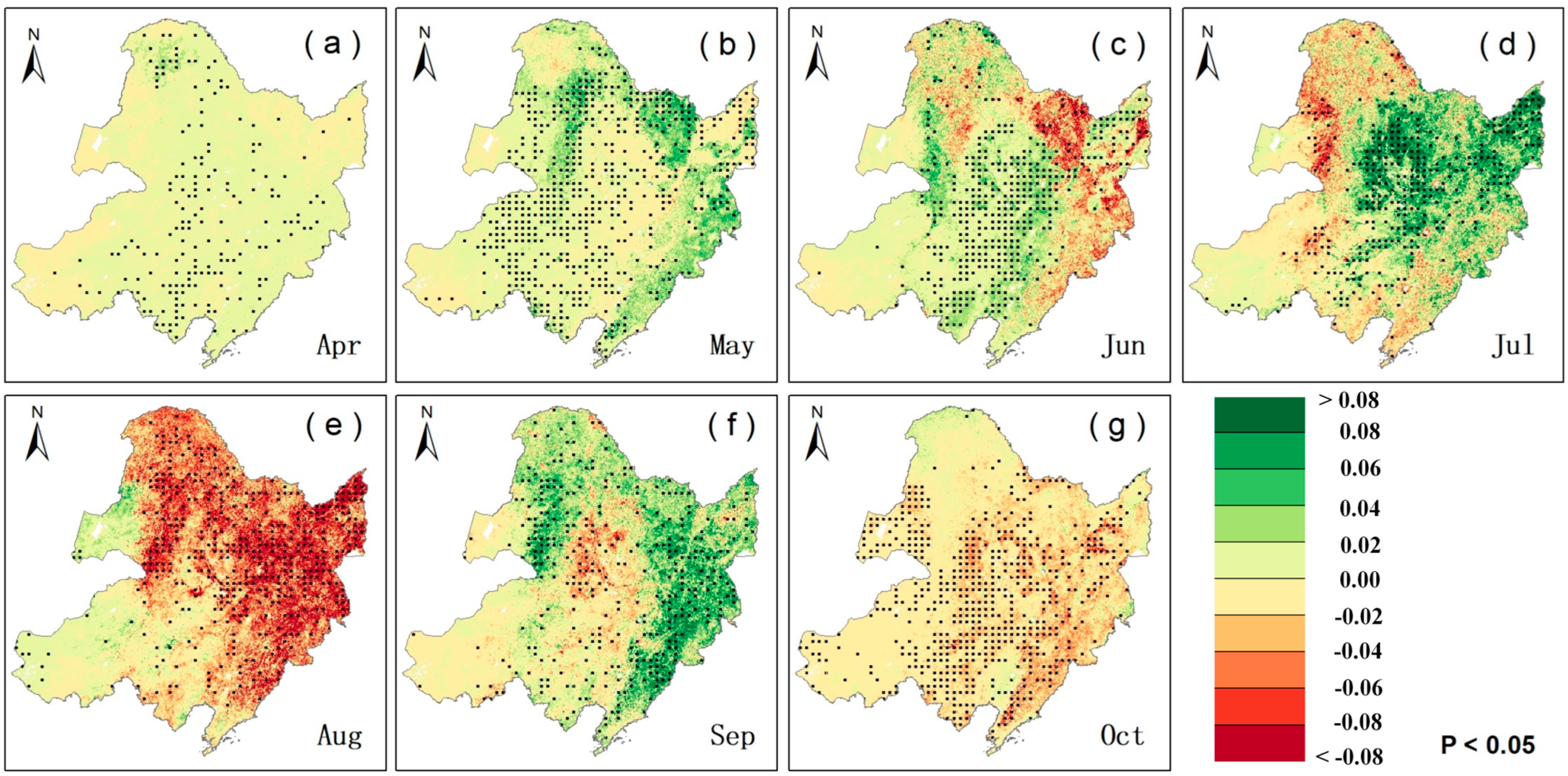
2.1. Trends in LAI and VLAI
2.2. Partial Correlation Analysis Between VLAI (or LAI) and Climatic Factors
2.2.1. Partial Correlation Analysis Between VLAI (or LAI) and Precipitation
2.2.2. Partial Correlation Analysis Between VLAI (or LAI) and Air Temperature
2.2.3. Partial Correlation Analysis Between VLAI (or LAI) and Solar Radiation
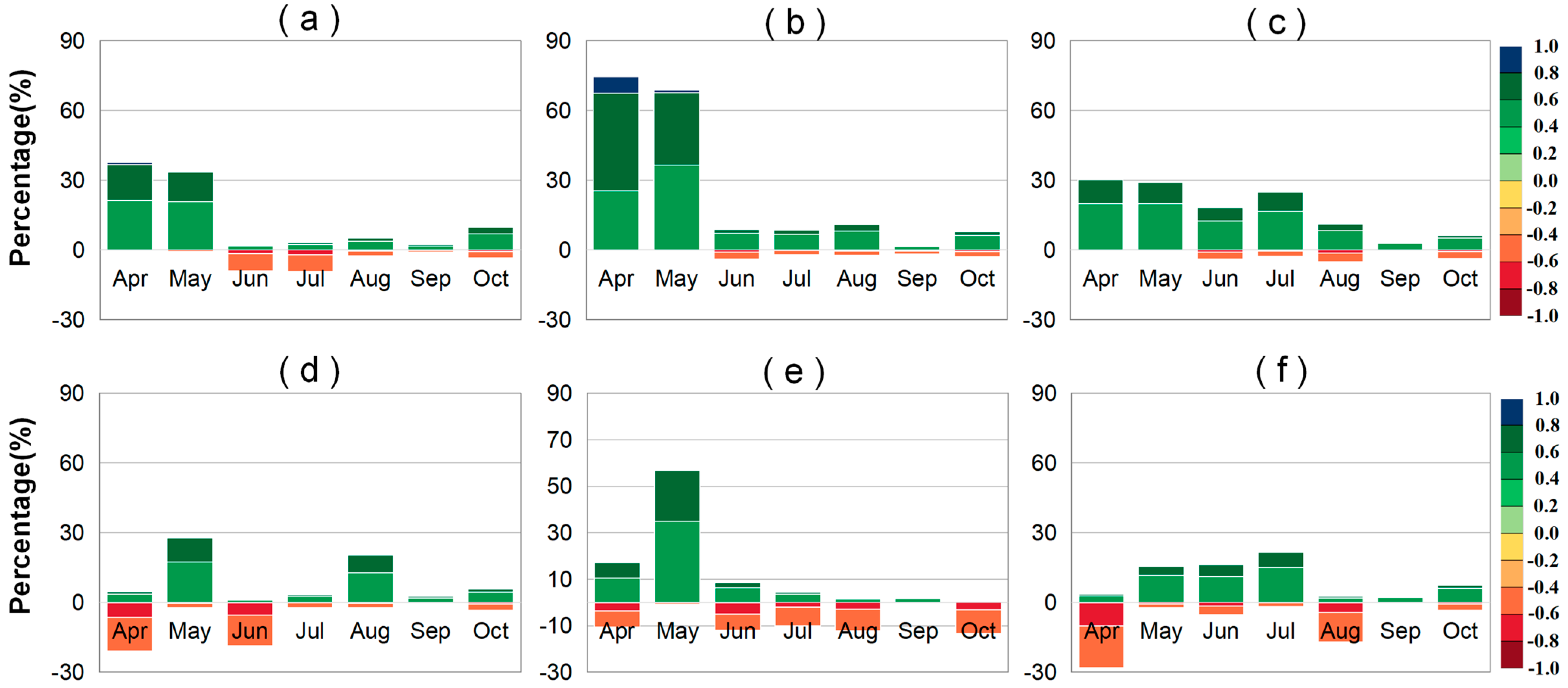
2.3. Lagged Effect of Climate Factors on LAI and VLAI
3. Discussion
3.1. Vegetation Canopy Development Changes at Finer Temporal Scale
3.2. Impact of Preseason Climate Factors on VLAI Changes
3.3. Uncertainties and Future Directions
4. Materials and Methods
4.1. Study Area
4.2. Datasets
4.3. Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Piao, S.L.; Wang, X.H.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.P.; Ciais, P.; Tommervik, H.; et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 2020, 1, 14–27. [Google Scholar] [CrossRef]
- Peng, S.S.; Chen, A.P.; Xu, L.; Cao, C.X.; Fang, J.Y.; Myneni, R.B.; Pinzon, J.E.; Tucker, C.J.; Piao, S.L. Recent change of vegetation growth trend in China. Environ. Res. Lett. 2011, 6, 044027. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Piao, S.L.; Myneni, R.B.; Huang, M.T.; Zeng, Z.Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Change 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Piao, S.L.; Ciais, P.; Huang, Y.; Shen, Z.H.; Peng, S.S.; Li, J.S.; Zhou, L.P.; Liu, H.Y.; Ma, Y.C.; Ding, Y.H.; et al. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43–51. [Google Scholar] [CrossRef]
- Yu, L.X.; Liu, Y.; Yang, J.C.; Liu, T.X.; Bu, K.; Li, G.S.; Jiao, Y.; Zhang, S.W. Asymmetric daytime and nighttime surface temperature feedback induced by crop greening across Northeast China. Agric. For. Meteorol. 2022, 325, 109136. [Google Scholar] [CrossRef]
- Yu, L.X.; Xue, Y.K.; Diallo, I. Vegetation greening in China and its effect on summer regional climate. Sci. Bull. 2021, 66, 13–17. [Google Scholar] [CrossRef]
- Li, X.; Du, H.; Zhou, G.; Mao, F.; Zhu, D.E.; Zhang, M.; Xu, Y.; Zhou, L.; Huang, Z. Spatiotemporal patterns of remotely sensed phenology and their response to climate change and topography in subtropical bamboo forests during 2001–2017: A case study in Zhejiang Province, China. GISci. Remote Sens. 2023, 60, 2163575. [Google Scholar] [CrossRef]
- Yu, L.X.; Liu, T.X.; Bu, K.; Yan, F.Q.; Yang, J.C.; Chang, L.P.; Zhang, S.W. Monitoring the long term vegetation phenology change in Northeast China from 1982 to 2015. Sci. Rep. 2017, 7, 14770. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.Z.; Wu, Z.F.; Fu, Y.H. Review of vegetation phenology trends in China in a changing climate. Prog. Phys. Geogr.-Earth Environ. 2022, 46, 829–845. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Piao, S.L.; Sun, Y.; Rogers, B.M.; Li, X.Y.; Lian, X.; Liu, Z.H.; Chen, A.P.; Peñuelas, J. Future reversal of warming-enhanced vegetation productivity in the Northern Hemisphere. Nat. Clim. Change 2022, 12, 581–586. [Google Scholar] [CrossRef]
- Piao, S.L.; Liu, Q.; Chen, A.P.; Janssens, I.A.; Fu, Y.S.; Dai, J.H.; Liu, L.L.; Lian, X.; Shen, M.G.; Zhu, X.L. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X.; Liu, Y.; Liu, T.X.; Yan, F.Q. Impact of recent vegetation greening on temperature and precipitation over China. Agric. For. Meteorol. 2020, 295, 108197. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Y.; Yan, F.; Lu, L.; Li, X.; Zhang, S.; Yang, J. Phenological control of vegetation biophysical feedbacks to the regional climate. Geogr. Sustain. 2024, 6, 100202. [Google Scholar] [CrossRef]
- Chen, X.; An, S.; Inouye, D.W.; Schwartz, M.D. Temperature and snowfall trigger alpine vegetation green-up on the world’s roof. Glob. Change Biol. 2015, 21, 3635–3646. [Google Scholar] [CrossRef]
- Zheng, J.; Ge, Q.; Hao, Z. Impacts of climate warming on plants phenophases in China for the last 40 years. Chin. Sci. Bull. 2002, 47, 1826–1831. [Google Scholar]
- Tian, F.; Cai, Z.; Jin, H.; Hufkens, K.; Scheifinger, H.; Tagesson, T.; Smets, B.; Van Hoolst, R.; Bonte, K.; Ivits, E. Calibrating vegetation phenology from Sentinel-2 using eddy covariance, PhenoCam, and PEP725 networks across Europe. Remote Sens. Environ. 2021, 260, 112456. [Google Scholar] [CrossRef]
- Andreatta, D.; Bachofen, C.; Dalponte, M.; Klaus, V.H.; Buchmann, N. Extracting flowering phenology from grassland species mixtures using time-lapse cameras. Remote Sens. Environ. 2023, 298, 113835. [Google Scholar] [CrossRef]
- Dominguez, D.L.; Cirrincione, M.A.; Deis, L.; Martínez, L.E. Impacts of Climate Change-Induced Temperature Rise on Phenology, Physiology, and Yield in Three Red Grape Cultivars: Malbec, Bonarda, and Syrah. Plants 2024, 13, 3219. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, X.; Shen, Y.; Wang, J.; Crimmins, T.; Scheifinger, H. An optimal method for validating satellite-derived land surface phenology using in-situ observations from national phenology networks. ISPRS J. Photogramm. Remote Sens. 2022, 194, 74–90. [Google Scholar] [CrossRef]
- Gong, Z.; Ge, W.; Guo, J.; Liu, J. Satellite remote sensing of vegetation phenology: Progress, challenges, and opportunities. ISPRS J. Photogramm. Remote Sens. 2024, 217, 149–164. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, Z.; Chen, Y.; Li, Z.; Tao, F. ChinaCropPhen1km: A high-resolution crop phenological dataset for three staple crops in China during 2000–2015 based on leaf area index (LAI) products. Earth Syst. Sci. Data 2020, 12, 197–214. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.J.; Yin, G.F.; Xie, Q.Y.; Xu, B.D.; Verger, A.; Descals, A.; Filella, I.; Peñuelas, J. Divergence in Autumn Phenology Extracted From Different Satellite Proxies Reveals the Timetable of Leaf Senescence Over Deciduous Forests. Geophys. Res. Lett. 2024, 51, e2023GL107346. [Google Scholar] [CrossRef]
- Li, X.; Fu, Y.H.; Chen, S.; Xiao, J.; Yin, G.; Li, X.; Zhang, X.; Geng, X.; Wu, Z.; Zhou, X. Increasing importance of precipitation in spring phenology with decreasing latitudes in subtropical forest area in China. Agric. For. Meteorol. 2021, 304, 108427. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Change Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Liu, Q.; Fu, Y.H.; Zeng, Z.; Huang, M.; Li, X.; Piao, S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob. Change Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef]
- Zhang, J.R.; Tong, X.J.; Zhang, J.S.; Meng, P.; Li, J.; Liu, P.R. Dynamics of phenology and its response to climatic variables in a warm-temperate mixed plantation. For. Ecol. Manag. 2021, 483, 118785. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Chen, A.; Wang, K.; Li, X.; Buermann, W.; Huntingford, C.; Peñuelas, J.; Xu, H.; Myneni, R.B. Seasonal biological carryover dominates northern vegetation growth. Nat. Commun. 2021, 12, 983. [Google Scholar] [CrossRef]
- Jump, A.S.; Ruiz-Benito, P.; Greenwood, S.; Allen, C.D.; Kitzberger, T.; Fensham, R.; Martínez-Vilalta, J.; Lloret, F. Structural overshoot of tree growth with climate variability and the global spectrum of drought-induced forest dieback. Glob. Change Biol. 2017, 23, 3742–3757. [Google Scholar] [CrossRef]
- Lian, X.; Peñuelas, J.; Ryu, Y.; Piao, S.; Keenan, T.F.; Fang, J.; Yu, K.; Chen, A.; Zhang, Y.; Gentine, P. Diminishing carryover benefits of earlier spring vegetation growth. Nat. Ecol. Evol. 2024, 8, 218–228. [Google Scholar] [CrossRef]
- Buermann, W.; Bikash, P.R.; Jung, M.; Burn, D.H.; Reichstein, M. Earlier springs decrease peak summer productivity in North American boreal forests. Environ. Res. Lett. 2013, 8, 024027. [Google Scholar] [CrossRef]
- Cohen, J.L.; Furtado, J.C.; Barlow, M.; Alexeev, V.A.; Cherry, J.E. Asymmetric seasonal temperature trends. Geophys. Res. Lett. 2012, 39, 4. [Google Scholar] [CrossRef]
- Zhang, Y.; Parazoo, N.C.; Williams, A.P.; Zhou, S.; Gentine, P. Large and projected strengthening moisture limitation on end-of-season photosynthesis. Proc. Natl. Acad. Sci. USA 2020, 117, 9216–9222. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.M.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res.-Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Wang, L.H.; Tian, F.; Wang, Y.H.; Wu, Z.D.; Schurgers, G.; Fensholt, R. Acceleration of global vegetation greenup from combined effects of climate change and human land management. Glob. Change Biol. 2018, 24, 5484–5499. [Google Scholar] [CrossRef]
- Meng, F.; Liu, D.; Wang, Y.; Wang, S.; Wang, T. Negative relationship between photosynthesis and late-stage canopy development and senescence over Tibetan Plateau. Glob. Change Biol. 2023, 29, 3147–3158. [Google Scholar] [CrossRef]
- Piao, S.; Wang, J.; Li, X.; Xu, H.; Zhang, Y. Spatio-temporal changes in the speed of canopy development and senescence in temperate China. Glob. Change Biol. 2022, 28, 7366–7375. [Google Scholar] [CrossRef]
- Zeng, Y.L.; Hao, D.L.; Park, T.; Zhu, P.; Huete, A.; Myneni, R.; Knyazikhin, Y.; Qi, J.B.; Nemani, R.R.; Li, F.; et al. Structural complexity biases vegetation greenness measures. Nat. Ecol. Evol. 2023, 7, 1790–1798. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhu, Z.C.; Zeng, H.; Myneni, R.B.; Zhang, Y.; Penuelas, J.; Piao, S.L. Seasonal peak photosynthesis is hindered by late canopy development in northern ecosystems. Nat. Plants 2022, 8, 1484–1492. [Google Scholar] [CrossRef]
- Chmura, H.E.; Kharouba, H.M.; Ashander, J.; Ehlman, S.M.; Rivest, E.B.; Yang, L.H. The mechanisms of phenology: The patterns and processes of phenological shifts. Ecol. Monogr. 2019, 89, e01337. [Google Scholar] [CrossRef]
- Ma, Q.; Hänninen, H.; Berninger, F.; Li, X.; Huang, J.G. Climate warming leads to advanced fruit development period of temperate woody species but divergent changes in its length. Glob. Change Biol. 2022, 28, 6021–6032. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Zhang, Y.; Yao, Y.; Meng, F.; Zhao, Q.; Zhang, Y. Contrasting temperature effects on the velocity of early-versus late-stage vegetation green-up in the Northern Hemisphere. Glob. Change Biol. 2022, 28, 6961–6972. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.E.; Tjoelker, M.G.; Aspinwall, M.J.; Reich, P.B.; Barton, C.V.; Medlyn, B.E.; Duursma, R.A. Does physiological acclimation to climate warming stabilize the ratio of canopy respiration to photosynthesis? New Phytol. 2016, 211, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants—An economic analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- McCarthy, M.; Enquist, B. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Agüera, E.; De la Haba, P. Leaf senescence in response to elevated atmospheric CO2 concentration and low nitrogen supply. Biol. Plant. 2018, 62, 401–408. [Google Scholar] [CrossRef]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, M.; Lin, T.; Zhang, J.; Jones, L.; Yao, X.; Geng, H.; Liu, Y.; Zhang, G.; Cao, X. Spatial heterogeneity of vegetation phenology caused by urbanization in China based on remote sensing. Ecol. Indic. 2023, 153, 110448. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y.; Li, Z.; Li, Y.; Zhang, Q.; Zan, M. Driving forces of the changes in vegetation phenology in the qinghai–tibet plateau. Remote Sens. 2021, 13, 4952. [Google Scholar] [CrossRef]
- Dow, C.; Kim, A.Y.; D’Orangeville, L.; Gonzalez-Akre, E.B.; Helcoski, R.; Herrmann, V.; Harley, G.L.; Maxwell, J.T.; McGregor, I.R.; McShea, W.J. Warm springs alter timing but not total growth of temperate deciduous trees. Nature 2022, 608, 552–557. [Google Scholar] [CrossRef]
- Tao, F.; Zhang, S.; Zhang, Z. Spatiotemporal changes of wheat phenology in China under the effects of temperature, day length and cultivar thermal characteristics. Eur. J. Agron. 2012, 43, 201–212. [Google Scholar] [CrossRef]
- Yang, Y.; Ren, W.; Tao, B.; Ji, L.; Liang, L.; Ruane, A.C.; Fisher, J.B.; Liu, J.; Sama, M.; Li, Z. Characterizing spatiotemporal patterns of crop phenology across North America during 2000–2016 using satellite imagery and agricultural survey data. ISPRS J. Photogramm. Remote Sens. 2020, 170, 156–173. [Google Scholar] [CrossRef]
- Kang, J.; Yang, Z.; Yu, B.; Ma, Q.; Jiang, S.; Shishov, V.V.; Zhou, P.; Huang, J.-G.; Ding, X. An earlier start of growing season can affect tree radial growth through regulating cumulative growth rate. Agric. For. Meteorol. 2023, 342, 109738. [Google Scholar] [CrossRef]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Change Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Buermann, W.; Forkel, M.; O’sullivan, M.; Sitch, S.; Friedlingstein, P.; Haverd, V.; Jain, A.K.; Kato, E.; Kautz, M.; Lienert, S. Widespread seasonal compensation effects of spring warming on northern plant productivity. Nature 2018, 562, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Piao, S.; Li, L.Z.; Li, Y.; Huntingford, C.; Ciais, P.; Cescatti, A.; Janssens, I.A.; Peñuelas, J.; Buermann, W. Summer soil drying exacerbated by earlier spring greening of northern vegetation. Sci. Adv. 2020, 6, eaax0255. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Joel, G.; Field, C.B.; Fung, I.Y. Toward an allocation scheme for global terrestrial carbon models. Glob. Change Biol. 1999, 5, 755–770. [Google Scholar] [CrossRef]
- Hartmann, H.; Bahn, M.; Carbone, M.; Richardson, A. Plant carbon allocation in a changing world—Challenges and progress. New Phytol. 2020, 227, 981–988. [Google Scholar] [CrossRef]
- Merganičová, K.; Merganič, J.; Lehtonen, A.; Vacchiano, G.; Sever MZ, O.; Augustynczik, A.L.; Grote, R.; Kyselová, I.; Mäkelä, A.; Yousefpour, R. Forest carbon allocation modelling under climate change. Tree Physiol. 2019, 39, 1937–1960. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, H.; Harrison, S.P.; Prentice, I.C.; Qiao, S.; Tan, S. Optimality principles explaining divergent responses of alpine vegetation to environmental change. Glob. Change Biol. 2023, 29, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Delucia, E.H.; Maherali, H.; Carey, E.V. Climate-driven changes in biomass allocation in pines. Glob. Change Biol. 2000, 6, 587–593. [Google Scholar] [CrossRef]
- Chen, C.; He, B.; Guo, L.; Zhang, Y.; Xie, X.; Chen, Z. Identifying critical climate periods for vegetation growth in the Northern Hemisphere. J. Geophys. Res. Biogeosci. 2018, 123, 2541–2552. [Google Scholar] [CrossRef]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- Zawadzki, J.; Cieszewski, C.J.; Zasada, M.; Lowe, R.C. Applying geostatistics for investigations of forest ecosystems using remote sensing imagery. Silva Fenn. 2005, 39, 599. [Google Scholar] [CrossRef]
- Liu, T.; Li, P.; Zhao, F.; Liu, J.; Meng, R. Early-Stage Mapping of Winter Canola by Combining Sentinel-1 and Sentinel-2 Data in Jianghan Plain China. Remote Sens. 2024, 16, 3197. [Google Scholar] [CrossRef]
- Yan, L.; Guan, Y.; Wang, H.; Lin, Y.; Yang, Y.; Wang, B.; Jiang, J. EIRAD: An Evidence-based Dialogue System with Highly Interpretable Reasoning Path for Automatic Diagnosis. IEEE J. Biomed. Health Inform. 2024, 28, 6141–6154. [Google Scholar] [CrossRef]
- Li, D.; Sun, Y.; Peng, J.; Cheng, S.; Yin, Z.; Cheng, N.; Liu, J.; Li, Z.; Xu, C. Dual Network Computation Offloading Based on DRL for Satellite-Terrestrial Integrated Networks. IEEE Trans. Mob. Comput. 2024, PP, 1–14. [Google Scholar] [CrossRef]
- Lv, Z.; Xu, B.; Zhong, L.; Chen, G.; Huang, Z.; Sun, R.; Huang, W.; Zhao, F.; Meng, R. Improved monitoring of southern corn rust using UAV-based multi-view imagery and an attention-based deep learning method. Comput. Electron. Agric. 2024, 224, 109232. [Google Scholar] [CrossRef]
- Bao, J.; Cheng, S.; Liu, J. PAM-FOG Net: A Lightweight Weed Detection Model Deployed on Smart Weeding Robots. ACM Trans. Sens. Netw. 2024, 1–29. [Google Scholar] [CrossRef]
- McDonough MacKenzie, C.; Gallinat, A.S.; Zipf, L. Low-cost observations and experiments return a high value in plant phenology research. Appl. Plant Sci. 2020, 8, e11338. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lee, C.K.; Wang, Z.; Wang, J.; Gu, Y.; Xie, J.; Law, Y.K.; Song, G.; Bonebrake, T.C.; Yang, X. Evaluating fine-scale phenology from PlanetScope satellites with ground observations across temperate forests in eastern North America. Remote Sens. Environ. 2022, 283, 113310. [Google Scholar] [CrossRef]
- Rouault, P.; Courault, D.; Pouget, G.; Flamain, F.; Diop, P.-K.; Desfonds, V.; Doussan, C.; Chanzy, A.; Debolini, M.; McCabe, M. Phenological and Biophysical Mediterranean Orchard Assessment Using Ground-Based Methods and Sentinel 2 Data. Remote Sens. 2024, 16, 3393. [Google Scholar] [CrossRef]
- Song, G.; Wang, J.; Zhao, Y.; Yang, D.; Lee, C.K.; Guo, Z.; Detto, M.; Alberton, B.; Morellato, P.; Nelson, B. Scale matters: Spatial resolution impacts tropical leaf phenology characterized by multi-source satellite remote sensing with an ecological-constrained deep learning model. Remote Sens. Environ. 2024, 304, 114027. [Google Scholar] [CrossRef]
- Gu, H.; Qiao, Y.; Xi, Z.; Rossi, S.; Smith, N.G.; Liu, J.; Chen, L. Warming-induced increase in carbon uptake is linked to earlier spring phenology in temperate and boreal forests. Nat. Commun. 2022, 13, 3698. [Google Scholar] [CrossRef]
- Yu, L.X.; Liu, Y.; Liu, T.X.; Yu, E.T.; Bu, K.; Jia, Q.Y.; Shen, L.D.; Zheng, X.M.; Zhang, S.W. Coupling localized Noah-MP-Crop model with the WRF model improved dynamic crop growth simulation across Northeast China. Comput. Electron. Agric. 2022, 201, 107323. [Google Scholar] [CrossRef]
- Ma, H.; Liang, S. Development of the GLASS 250-m leaf area index product (version 6) from MODIS data using the bidirectional LSTM deep learning model. Remote Sens. Environ. 2022, 273, 112985. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- ESA Land Cover Climate Change Initiative. Product User Guide Version 2.0 [Dataset]. The European Space Agency (ESA) CCI (Climate Change Initiative). 2022. Available online: https://data.ceda.ac.uk/neodc/esacci/land_cover/data (accessed on 26 May 2024).
- Shen, M.; Zhao, W.; Jiang, N.; Liu, L.; Cao, R.; Yang, W.; Zhu, X.; Wang, C.; Chen, X.; Chen, J. Challenges in remote sensing of vegetation phenology. Innov. Geosci. 2024, 2, 100070. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Yu, L.; Li, X.; Gao, L.; Bao, L.; Chang, X.; Gao, X.; Cai, Z. Assessing Vegetation Canopy Growth Variations in Northeast China. Plants 2025, 14, 143. https://doi.org/10.3390/plants14010143
Lu L, Yu L, Li X, Gao L, Bao L, Chang X, Gao X, Cai Z. Assessing Vegetation Canopy Growth Variations in Northeast China. Plants. 2025; 14(1):143. https://doi.org/10.3390/plants14010143
Chicago/Turabian StyleLu, Lijie, Lingxue Yu, Xuan Li, Li Gao, Lun Bao, Xinyue Chang, Xiaohong Gao, and Zhongquan Cai. 2025. "Assessing Vegetation Canopy Growth Variations in Northeast China" Plants 14, no. 1: 143. https://doi.org/10.3390/plants14010143
APA StyleLu, L., Yu, L., Li, X., Gao, L., Bao, L., Chang, X., Gao, X., & Cai, Z. (2025). Assessing Vegetation Canopy Growth Variations in Northeast China. Plants, 14(1), 143. https://doi.org/10.3390/plants14010143