
The Identification and Analysis of the Self-Incompatibility Pollen Determinant Factor SLF in Lycium barbarum

Abstract
1. Introduction
2. Results
2.1. Identification and Cloning of the F-Box Gene Family in L. barbarum
2.2. Physicochemical Properties, Subcellular Localization Prediction, and Functional Prediction of S2-LbSLFs
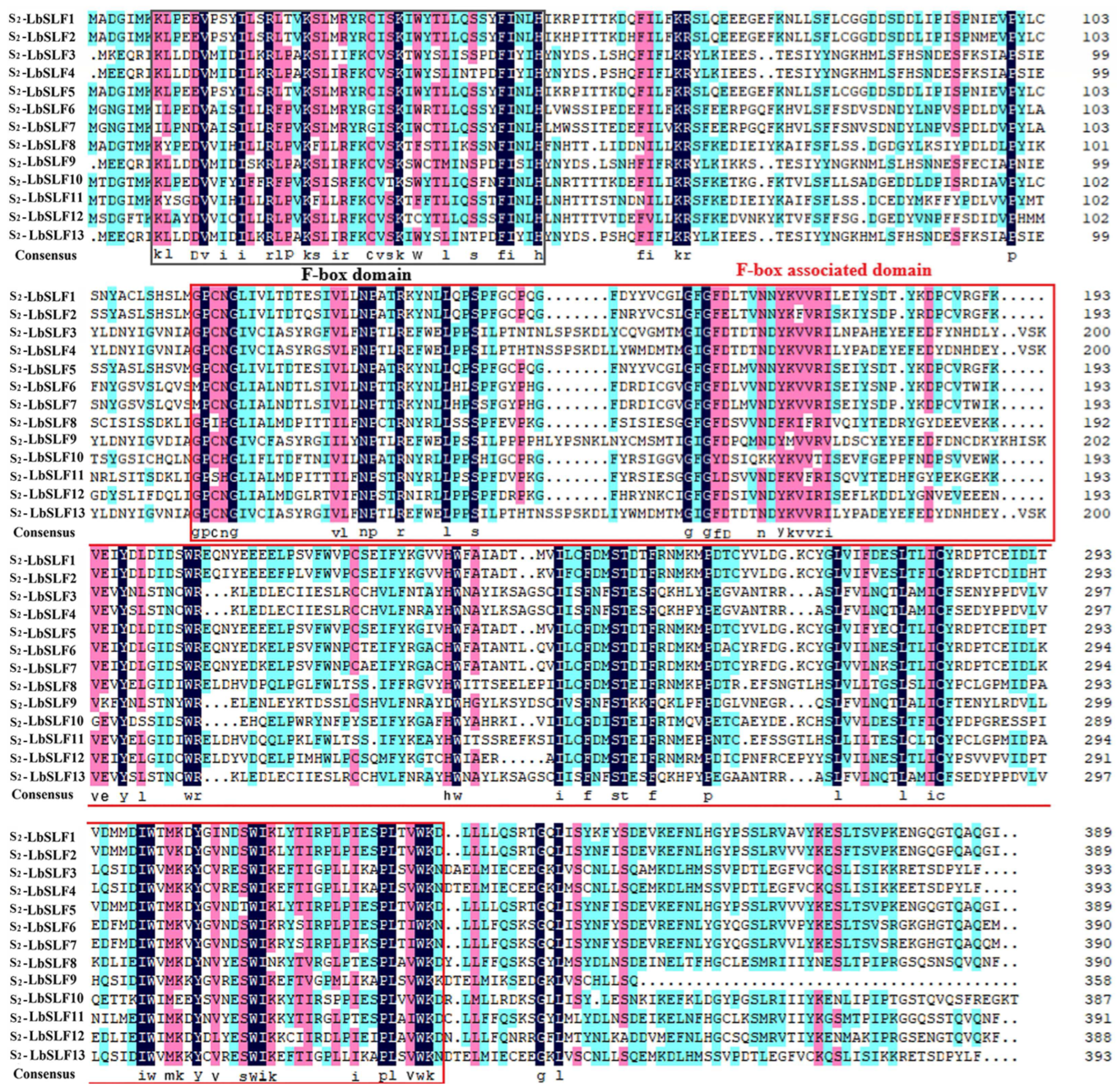
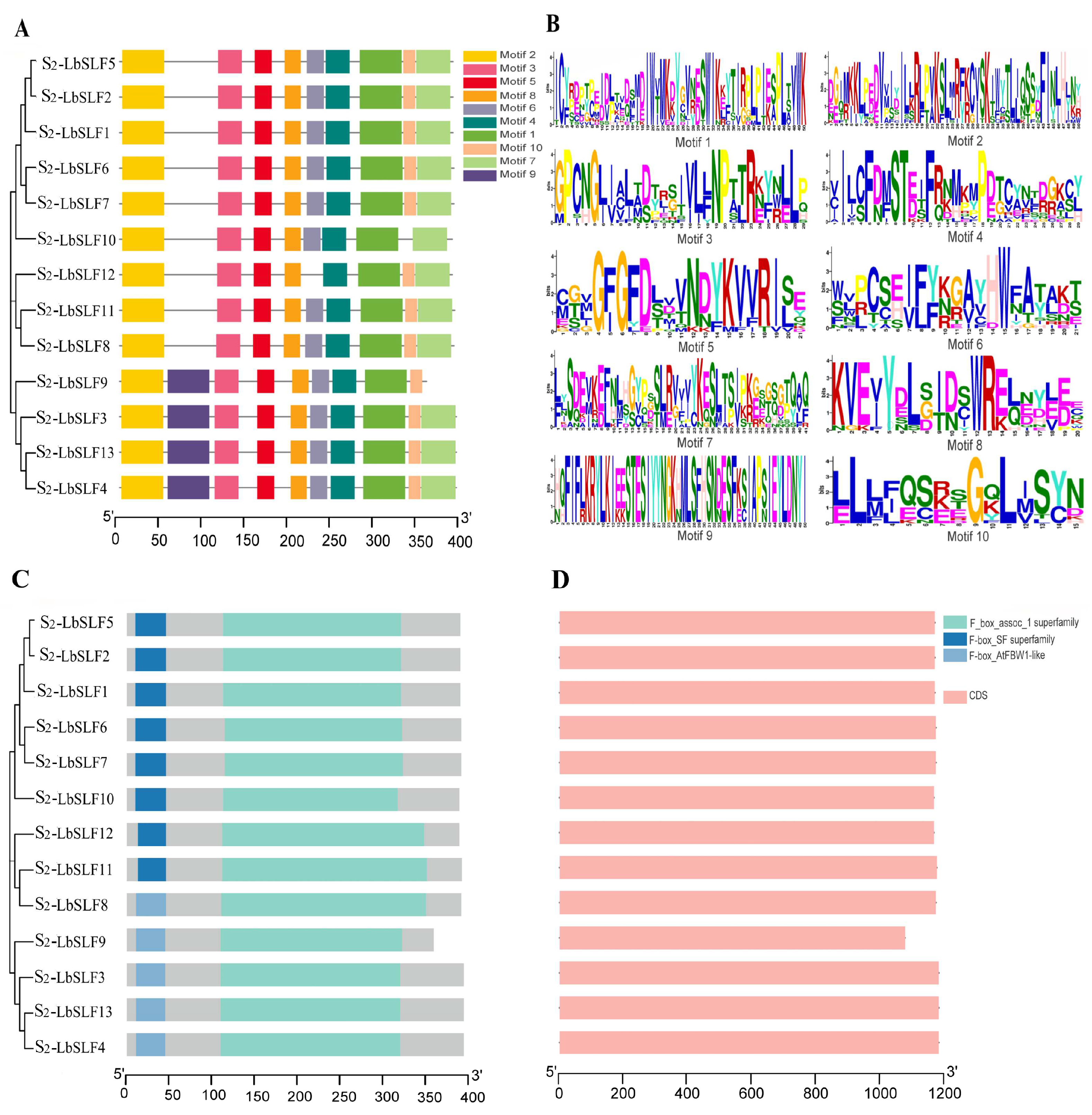
2.3. Conservation Motifs, Structural Domains, and Gene Structure of S2-LbSLF Gene Members
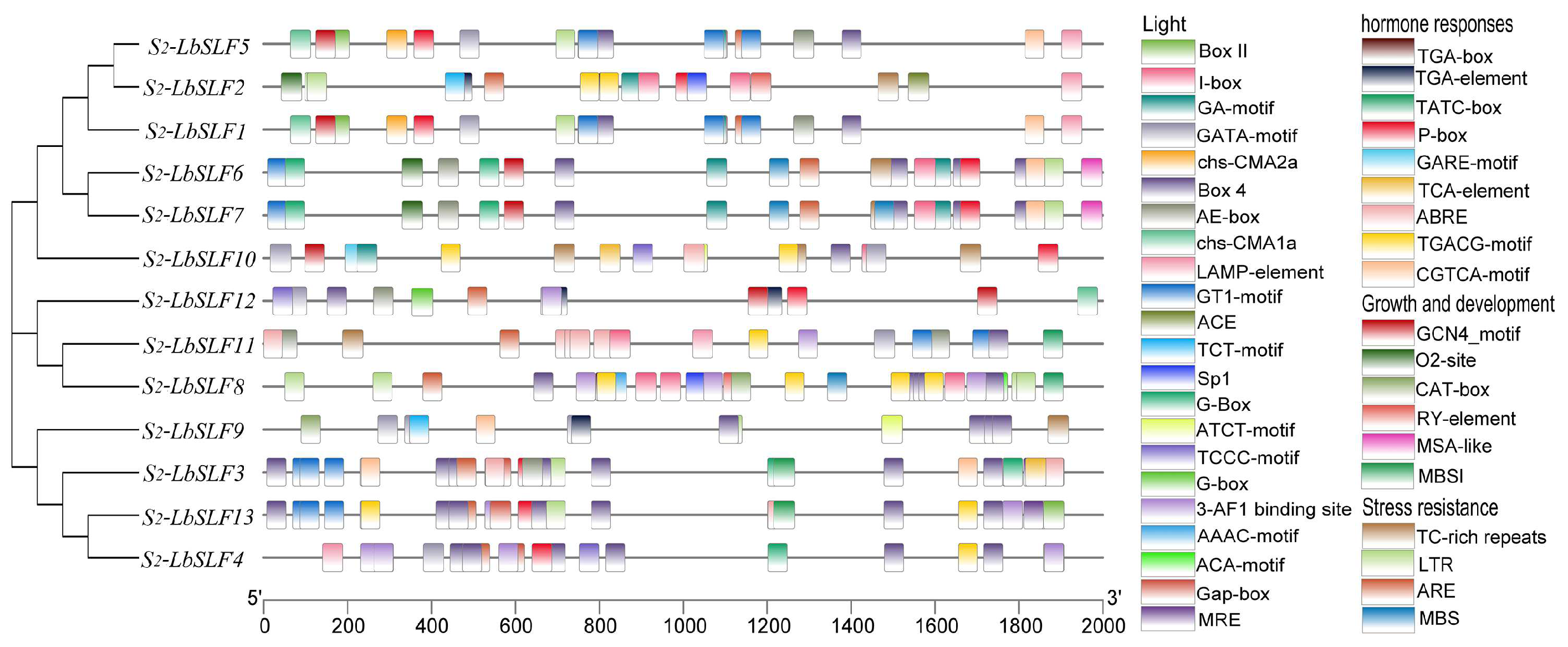
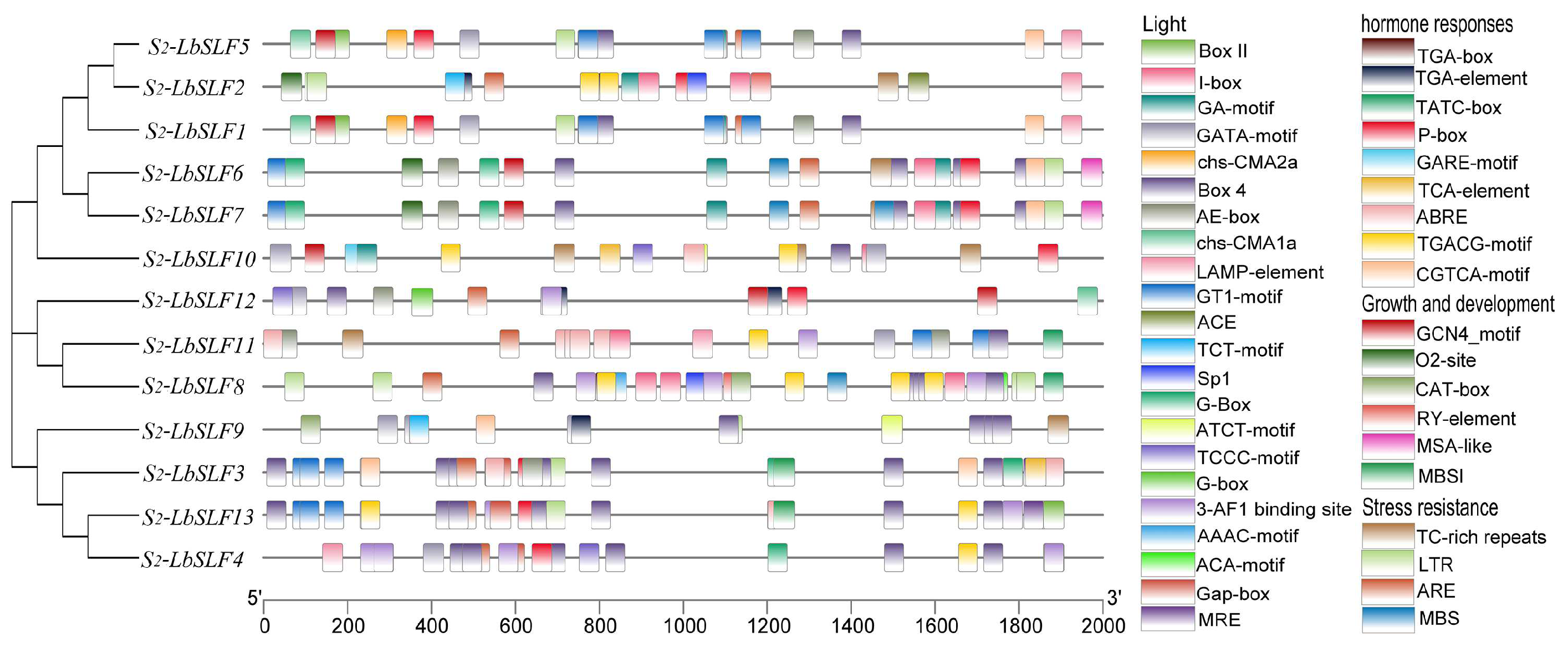
2.4. Analysis of Promoter Cis-Acting Elements
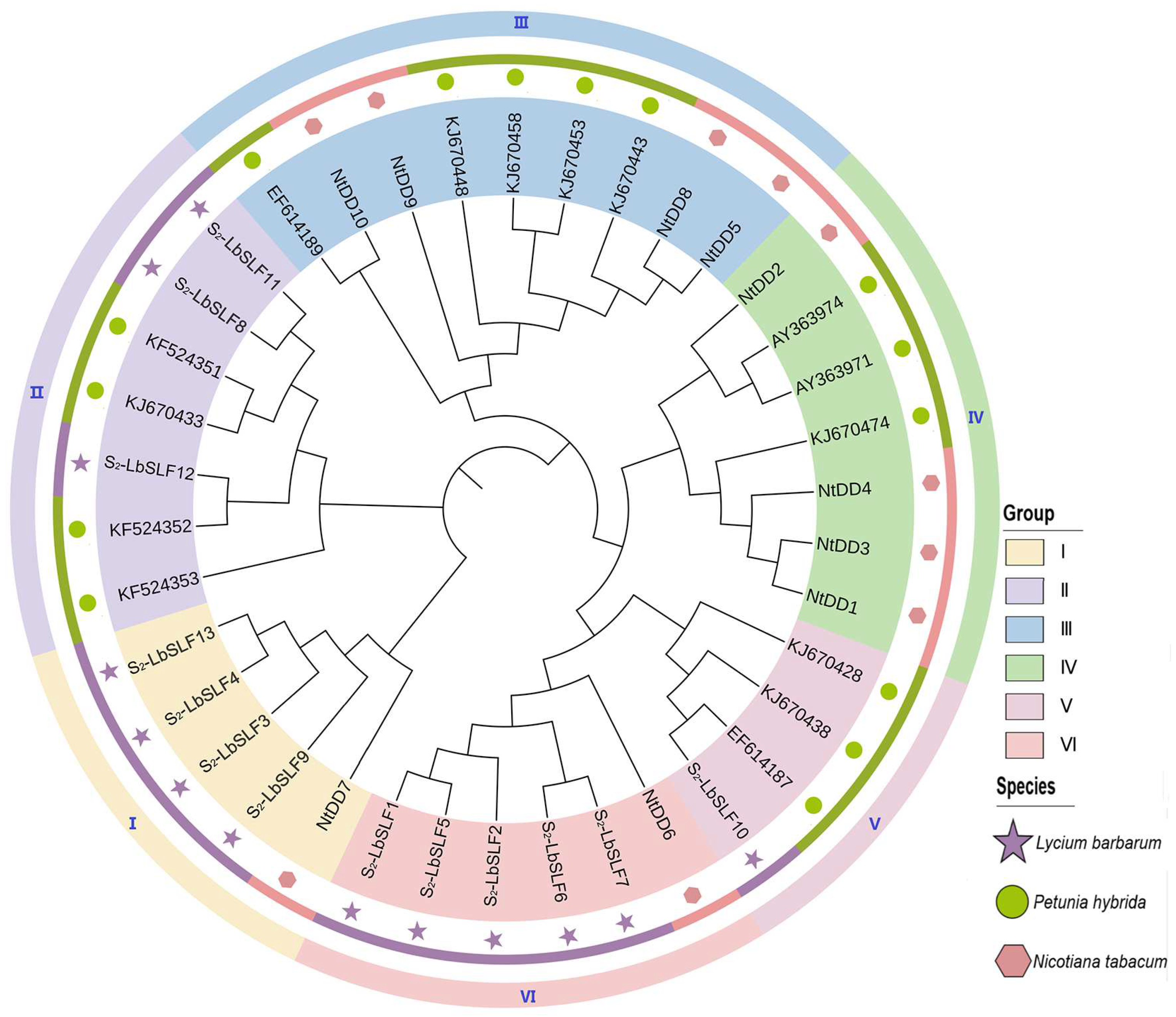
2.5. Phylogenetic Analysis and Classification of the L. barbarum S2-LbSLF Gene Family
2.6. Density and Collinearity Analysis of S2-LbSLF Genes
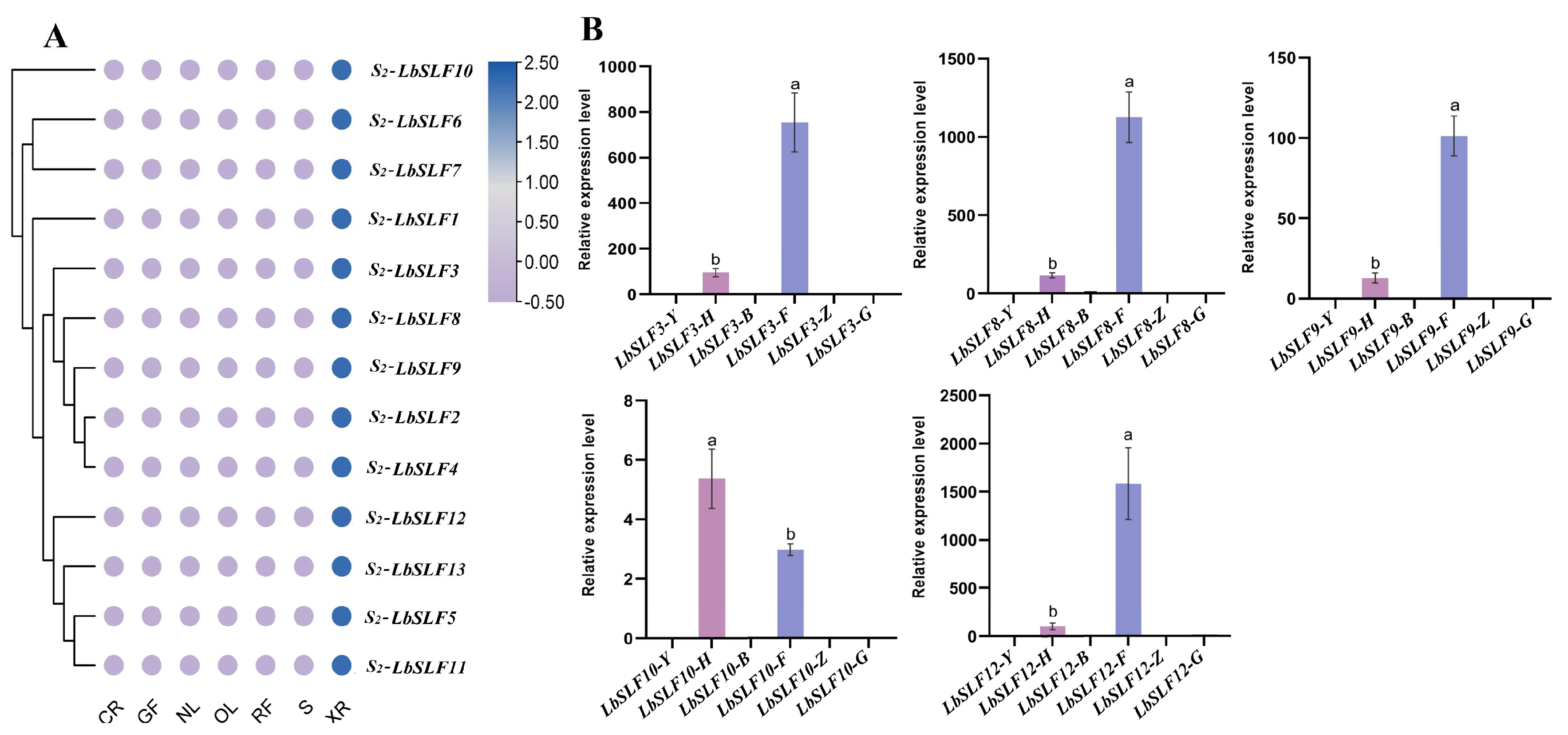
2.7. Analysis of Pollen-Specific Expression Patterns of S2-LbSLFs Genes
2.8. Interaction Analysis of SLF and S-RNase
3. Discussion
3.1. Cloning of Full-Length Sequences for 13 SLF Genes Located on Chromosome 2 in L. barbarum
3.2. Similarity in the Structure of F-Box Genes at the S Locus in Angiosperms
3.3. S2-LbSLF1–S2-LbSLF13 Are Good Candidates for the S-Determination Factor of L. barbarum Pollen
3.4. Physical Interactions between S2-LbSLFs and S-RNase
4. Materials and Methods
4.1. Identification of the L. barbarum F-Box Gene Family
4.2. Gene-Specific Amplification and Sequence Analysis
4.3. Physicochemical Property Analysis of S2-LbSLFs Genes
4.4. Phylogenetic Tree Development Analysis
4.5. Analysis of Conserved Protein Motifs, Protein Domains, and Gene Structure
4.6. Analysis of Cis Elements in SLF Gene Family Members
4.7. Analysis of S2-LbSLFs Gene Density and Collinearity
4.8. RNA Extraction and Expression Patterns of S2-LbSLFs
4.9. Yeast Two-Hybrid Analysis of Interaction between SLF and S-RNase
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Skenderidis, P.; Kerasioti, E.; Karkanta, E.; Stagos, D.; Kouretas, D.; Petrotos, K.; Hadjichristodoulou, C.; Tsakalof, A. Assessment of the antioxidant and antimutagenic activity of extracts from goji berry of Greek cultivation. Toxicol. Rep. 2018, 5, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yan, Y.M.; Zhang, L.T.; Mi, J.; Yu, L.M.; Zhang, F.F.; Lu, L.; Luo, Q.; Li, X.Y.; Zhou, X.; et al. A comprehensive review of goji berry processing and utilization. Food Sci. Nutr. 2023, 11, 7445–7457. [Google Scholar] [CrossRef]
- Jiao, E.; Li, X.Y.; He, J.; Dai, G. Pollination Compatibility of Six Lycium barbarum L. Varieties. Agric. Sci. Technol. 2013, 14, 1542–1544. [Google Scholar]
- Hu, J.B.; Liu, C.C.; Du, Z.Z.; Guo, F.R.; Song, D.; Wang, N.; Wei, Z.M.; Jiang, J.D.; Cao, Z.H.; Shi, C.M.; et al. Transposable elements cause the loss of self-incompatibility in citrus. Plant Biotech. J. 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, Y.; Zhang, H.; Song, Y.Z.; Zhao, F.; Zhang, Y.; Zhu, S.H.; Zhang, H.K.; Zhou, Z.D.; Guo, H.; et al. Origin, loss, and regain of self-incompatibility in angiosperms. Plant Cell 2022, 34, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Li, Y.Y.; Zhao, X.W.; Zhang, C.L.; Liu, D.K.; Lan, S.R.; Yin, W.L.; Liu, Z.J. Molecular insights into self-incompatibility systems: From evolution to breeding. Plant Commun. 2023, 100719, 2590–3462. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Carpenter, R.; Dickinson, H.G.; Coen, E.S. Origin of allelic diversity in antirrhinum S locus RNases. Plant Cell 1996, 8, 805–814. [Google Scholar] [PubMed]
- Tian, H.Y.; Zhang, H.K.; Huang, H.Q.; Zhang, Y.; Xue, Y.B. Phase separation of S-RNase promotes self-incompatibility in Petunia hybrida. J. Integr. Plant Biol. 2023. [Google Scholar] [CrossRef]
- Ren, Y.; Hua, Q.Z.; Pan, J.Y.; Zhang, Z.K.; Zhao, J.T.; He, X.H.; Qin, Y.H.; Hu, G.B. SKP1-like protein, CrSKP1-e, interacts with pollen-specific F-box proteins and assembles into SCF-type E3 complex in ‘Wuzishatangju’ (Citrus reticulata Blanco) pollen. PeerJ 2020, 8, e10578. [Google Scholar] [CrossRef]
- Liang, M.; Cao, Z.H.; Zhu, A.D.; Liu, Y.L.; Tao, M.Q.; Yang, H.Y.; Xu, Q.; Wang, S.H.; Liu, J.J.; Li, Y.P.; et al. Evolution of self-compatibility by a mutant Sm-RNase in citrus. Nat. Plants 2020, 6, 131–142. [Google Scholar] [CrossRef]
- Sun, L.H.; Cao, S.Y.; Zheng, N.; Kao, T.H. Analyses of Cullin1 homologs reveal functional redundancy in S-RNase-based self-incompatibility and evolutionary relationships in eudicots. Plant Cell 2023, 35, 673–699. [Google Scholar] [CrossRef]
- Kubo, K.I.; Tsukahara, M.; Fujii, S.; Murase, K.; Wada, Y.; Entani, T.; Iwano, M.; Takayama, S. Cullin1-P is an essential component of Non-self recognition system in self-incompatibility in Petunia. Plant Cell Physiol. 2016, 57, 2403–2416. [Google Scholar] [CrossRef] [PubMed]
- Sijacic, P.; Wang, X.; Skirpan, A.L.; Wang, Y.; Dowd, P.E.; McCubbin, A.G.; Huang, S.; Kao, T.H. Identification of the pollen determinant of S-RNase-mediated selfincompatibility. Nature 2004, 429, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Wang, F.; Zhao, L.; Zhou, J.L.; Lai, Z.; Zhang, Y.S.P.; Robbins, T.P.; Xue, Y.B. The F-box protein AhSLF-S2 controls the pollen function of S-RNase-based self-incompatibility. Plant Cell 2004, 16, 2307–2322. [Google Scholar] [CrossRef]
- Honsho, C. Self-incompatibility related to seedless fruit production in Citrus plants. Hort. J. 2023, 92, 1–12. [Google Scholar] [CrossRef]
- Kubo, K.I.; Entani, T.; Takara, A.; Wang, N.; Fields, A.M.; Hua, Z.H.; Toyoda, M.; Kawashima, S.I.; Ando, T.; Isogai, A.; et al. Collaborative non-self recognition system in S-RNase-based self-incompatibility. Science 2010, 330, 796–799. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, D.; Tao, R. Recognition of S-RNases by an S locus F-box like protein and an S haplotype-specific F-box like protein in the Prunus-specific self-incompatibility system. Plant Mol. Biol. 2019, 100, 367–378. [Google Scholar] [CrossRef]
- Zhao, H.; Song, Y.Z.; Li, J.H.; Zhang, Y.; Huang, H.Q.; Li, Q.; Zhang, Y.; Xue, Y.B. Primary restriction of S-RNase cytotoxicity by a stepwise ubiquitination and degradation pathway in Petunia hybrida. New Phytol. 2021, 231, 1249–1264. [Google Scholar] [CrossRef]
- Liu, W.; Fan, J.B.; Li, J.H.; Song, Y.Z.; Li, Q.; Zhang, Y.; Xue, Y.B. SCF (SLF)-mediated cytosolic degradation of S-RNase is required for cross-pollen compatibility in S-RNase-based self-incompatibility in Petunia hybrida. Front. Genet. 2014, 5, 228. [Google Scholar] [CrossRef]
- Vieira, J.; Rocha, S.; Vázquez, N.; López-Fernández, H.; Fdez-Riverola, F.; Reboiro-Jato, M.; Vieira, C.P. Predicting specificities under the Non-self gametophytic Self-incompatibility recognition model. Front. Plant Sci. 2019, 10, 879. [Google Scholar] [CrossRef]
- Li, Y.; Duan, X.W.; Wu, C.B.; Yu, J.; Liu, C.S.; Wang, J.; Zhang, X.M.; Yan, G.H.; Jiang, F.; Li, T.Z.; et al. Ubiquitination of S4-RNase by S-locus F-box like 2 contributes to self-compatibility of Sweet Cherry ‘Lapins’. Plant Physiol. 2020, 184, 4. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.C.; Huang, J.; Zhang, Y.Q.; Li, P.X.; Zhang, G.Q.; Xu, Q.; Chen, L.J.; Wang, J.Y.; Luo, Y.B.; Liu, Z.J. Lack of S-RNase-based gametophytic Self-incompatibility in Orchids suggests that this system evolved after the monocot-eudicot split. Front. Plant Sci. 2017, 8, 1106. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Ma, W.S.; Han, B.; Liang, L.Z.; Zhang, Y.S.; Hong, G.F.; Xue, Y.B. An F-box gene linked to the self-incompatibility (S) locus of Antirrhinum is expressed specifically in pollen and tapetum. Plant Mol. Biol. 2002, 50, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Entani, T.; Iwano, M.; Shiba, H.; Che, F.S.; Isogai, A.; Takayama, S. Comparative analysis of the self-incompatibility (S-) locus region of Prunus mume: Identification of a pollen-expressed F-box gene with allelic diversity. Genes Cells 2003, 8, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, K.; Sassa, H.; Dandekar, A.M.; Gradziel, T.M.; Tao, R.; Hirano, H. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotypespecific polymorphism. Plant Cell 2003, 15, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Ikeda, K.; Ushijima, K.; Sassa, H.; Tao, R. A pollen-expressed gene for a novel protein with an F-box motif that is very tightly linked to a gene for S-RNase in two species of cherry, Prunus cerasus and P. avium. Plant Cell Physiol. 2003, 44, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; McCubbin, A.G.; Kao, T.H. Genetic mapping and molecular characterization of the self-incompatibility (S) locus in Petunia inflata. Plant Mol. Biol. 2003, 53, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tsukamoto, T.; Yi, K.W.; Wang, X.; Huang, S.; McCubbin, A.G.; Kao, T.H. Chromosome walking in the Petunia inflata self-incompatibility (S-) locus and gene identification in an 881-kb contig containing S2-RNase. Plant Mol. Biol. 2004, 54, 727–742. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Ando, T.; Watanabe, H.; Marchesi, E.; Kao, T.H. Duplication of the S-locus F-box gene is associated with breakdown of pollen function in an S-haplotype identified in a natural population of self-incompatible Petunia axillaris. Plant Mol. Biol. 2005, 57, 141–153. [Google Scholar] [CrossRef]
- Wheeler, D.; Newbigin, E. Expression of 10 S-class SLF-like genes in Nicotiana alata pollen and its implications for understanding the pollen factor of the S locus. Genetics 2007, 177, 2171–2180. [Google Scholar] [CrossRef]
- Matsumoto, D.; Yamane, H.; Abe, K.; Tao, R. Identification of a Skp1-like protein interacting with SFB, the pollen S determinant of the gametophytic self-incompatibility in prunus. Plant Physiol. 2012, 159, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda-Garcia, E.; Fulton, E.C.; Parlan, E.V.; O’Connor, L.E.; Fleming, A.A.; Replogle, A.J.; Rocha-Sosa, M.; Gendron, J.M.; Thines, B. Unique n-terminal interactions connect F-box stress induced (FBS) proteins to a WD40 repeat-like protein pathway in Arabidopsis. Plants 2021, 10, 2228. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Zhang, Y.; Song, Y.Z.; Zhang, H.; Fan, J.B.; Li, Q.; Zhang, D.F.; Xue, Y.B. Electrostatic potentials of the S-locus F-box proteins contribute to the pollen S specificity in self-incompatibility in Petunia hybrida. Plant J. 2017, 89, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.H.; Williams, J.S.; Wang, N.; Khatri, W.A.; Roman, D.S.; Kao, T.H. Use of domain-swapping to identify candidate amino acids involved in differential interactions between two allelic variants of type-1 S-locus F-box protein and S3-RNase in Petunia inflata. Plant Cell Physiol. 2018, 59, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.I.; Paape, T.; Hatakeyama, M.; Entani, T.; Takara, A.; Kajihara, K.; Tsukahara, M.; Shimizu-Inatsugi, R.; Shimizu, K.K.; Takayama, S. Gene duplication and genetic exchange drive the evolution of S-RNase-based self-incompatibility in Petunia. Nat. Plants 2015, 1, 14005. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.S.; Der, J.P.; DePamphilis, C.W.; Kao, T.H. Transcriptome analysis reveals the same 17 S-Locus F-box genes in two haplotypes of the self-incompatibility locus of Petunia inflata. Plant Cell 2014, 26, 2873–2888. [Google Scholar] [CrossRef]
- Wang, C.P.; Wu, J.L.; Gao, Y.; Dai, G.L.; Shang, X.H.; Ma, H.J.; Zhang, X.; Xu, W.D.; Qin, K. Localization of S-locus-related self-incompatibility in Lycium barbarum Based on BSA Analysis. Horticulturae 2024, 10, 190. [Google Scholar] [CrossRef]
- McClure, B.A.; Haring, V.; Ebert, P.R.; Anderson, M.A.; Simpson, R.J.; Sakiyama, F.; Clarke, A.E. Style self-incompatibility gene products of Nicotlana alata are ribonucleases. Nature 1989, 342, 955–957. [Google Scholar] [CrossRef]
- Zhang, Z.; Qiu, W.M.; Liu, W.; Han, X.J.; Wu, L.H.; Yu, M.; Qiu, X.L.; He, Z.Q.; Li, H.Y.; Zhuo, R.Y. Genome-wide characterization of the hyperaccumulator Sedum alfredii F-box family under cadmium stress. Sci Rep. 2021, 11, 3023. [Google Scholar] [CrossRef]
- Lee, H.S.; Huang, S.; Kao, T.H. S proteins control rejection of incompatible pollen in Petunia inflata. Nature 1994, 367, 560–563. [Google Scholar] [CrossRef]
- Williams, J.S.; Natale, C.A.; Wang, N.; Li, S.; Brubaker, T.R.; Sun, P.; Kao, T.H. Four previously identified Petunia inflata S-Locus F-box genes are involved in pollen specificity in self-incompatibility. Mol. Plant 2014, 7, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Pimenta, J.; Gomes, A.; Laia, J.; Rocha, S.; Heitzler, P.; Vieira, C.P. The identification of the Rosa S-locus and implications on the evolution of the Rosaceae gametophytic self-incompatibility systems. Sci Rep. 2021, 11, 3710. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, C.; Ritter, E.; Barone, A.; Debener, T.; Walkemeier, B.; Schachtschabel, U.; Kaufmann, H.; Thompson, R.D.; Bonierbale, M.W.; Ganal, M.W.; et al. RFLP maps of potato and their alignment with the homoeologous tomato genome. Theor. Appl. Genet. 1991, 83, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.H.; Zhang, Y.; Copsy, L.; Han, Q.Q.; Zheng, D.F.; Coen, E.; Xue, Y.B. The snapdragon genomes reveal the evolutionary dynamics of the S-locus supergene. Mol. Biol. Evol. 2023, 40, msad080. [Google Scholar] [CrossRef] [PubMed]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.S.; Bliek, M.; Boersma, M.R.; Borghi, L.; Bruggmann, R.; et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2016, 2, 16074. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Moriya, S.; Haji, T.; Abe, K. Isolation and characterization of multiple F-box genes linked to the S9—And S10 -RNase in apple (Malus × domestica Borkh.). Plant Reprod. 2013, 26, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.M.; Yin, H.; Qiao, X.; Tan, X.; Gu, C.; Wang, B.H.; Cheng, R.; Wang, Y.Z.; Zhang, S.L. F-box genes: Genome-wide expansion, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri). Plant Sci. 2016, 253, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Williams, J.S.; Sun, L.H.; Kao, T.H. Sequence analysis of the Petunia inflata S-locus region containing 17 S-locus F-box genes and the S-RNase gene involved in self-incompatibility. Plant J. 2020, 104, 1348–1368. [Google Scholar] [CrossRef]
- Feng, C.H.; Niu, M.X.; Liu, X.; Bao, Y.; Liu, S.J.; Liu, M.Y.; He, F.; Han, S.; Liu, C.; Wang, H.L.; et al. Genome-wide analysis of the FBA subfamily of the poplar F-box gene family and its role under drought stress. Int. J. Mol. Sci. 2023, 24, 4823. [Google Scholar] [CrossRef]
- Akash; Parida, A.P.; Srivastava, A.; Mathur, S.; Sharma, A.K.; Kumar, R. Identification, evolutionary profiling, and expression analysis of F-box superfamily genes under phosphate deficiency in tomato. Plant Physiol. Biochem. 2021, 162, 349–362. [Google Scholar] [CrossRef]
- Wu, G.F.; Cao, A.H.; Wen, Y.H.; Bao, W.C.; She, F.W.; Wu, W.Z.; Zheng, S.; Yang, N. Characteristics and functions of MYB (v-Myb avivan myoblastsis virus oncogene homolog)-related genes in Arabidopsis thaliana. Genes 2023, 14, 2026. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.F.; Xia, X.H.; Yao, W.J.; Cheng, Z.H.; Zhang, X.M.; Jiang, J.H.; Zhou, B.R.; Jiang, T.B. Genome-wide identification and expression patterns of the F-box family in poplar under salt stress. Int. J. Mol. Sci. 2022, 23, 10934. [Google Scholar] [CrossRef] [PubMed]
- Chunthong, K.; Pitnjam, K.; Chakhonkaen, S.; Sangarwut, N.; Panyawut, N.; Wasinanon, T.; Ukoskit, K.; Muangprom, A. Differential. Drought responses in F-box gene expression and grain yield between two Rice groups with contrasting drought tolerance. J. Plant Growth. Regul. 2017, 36, 970–982. [Google Scholar] [CrossRef]
- Sun, L.H.; Williams, J.S.; Li, S.; Wu, L.; Khatri, W.A.; Stone, P.G.; Keebaugh, M.D.; Kao, T.H. S-locus F-box proteins are solely responsible for pollen function in S-RNase-based self-incompatibility of Petunia Pollen. Plant Cell 2018, 30, 2959–2972. [Google Scholar] [CrossRef]
- Xu, K.H.; Wu, N.; Yao, W.B.; Li, X.W.; Zhou, Y.G.; Li, H.Y. The biological function and roles in phytohormone signaling of the F-box protein in plants. Agronomy 2021, 11, 2360. [Google Scholar] [CrossRef]
- Saxena, H.; Negi, H.; Sharma, B. Role of F-box E3-ubiquitin ligases in plant development and stress responses. Plant Cell Rep. 2023, 42, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Guo, J.L.; Zhou, F.L.; Singh, S.; Xu, X.; Xie, Q.; Yang, Z.B.; Huang, C.F. F-box protein RAE1 regulates the stability of the aluminum-resistance transcription factor STOP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 319–327. [Google Scholar] [CrossRef]
- Li, X.C.; Sun, Y.; Liu, N.; Wang, P.; Pei, Y.K.; Liu, D.; Ma, X.W.; Ge, X.Y.; Li, F.G.; Hou, Y.X. Enhanced resistance to Verticillium dahliae mediated by an F-box protein GhACIF1 from Gossypium hirsutum. Plant Sci. 2019, 284, 127–134. [Google Scholar] [CrossRef]
- Sun, P.L.; Li, S.; Lu, D.H.; Williams, J.S.; Kao, T.H. Pollen S-locus F-box proteins of Petunia involved in S-RNase-based self-incompatibility are themselves subject to ubiquitin-mediated degradation. Plant J. 2015, 83, 213–223. [Google Scholar] [CrossRef]
- Xu, C.; Li, M.F.; Wu, J.K.; Guo, H.; Li, Q.; Zhang, Y.; Chai, J.J.; Li, T.Z.; Xue, Y.B. Identification of a canonical SCF(SLF) complex involved in S-RNase-based self-incompatibility of Pyrus (Rosaceae). Plant Mol. Biol. 2013, 81, 245–257. [Google Scholar] [CrossRef]
- Sun, P.L.; Kao, T.H. Self-Incompatibility in Petunia inflata: The relationship between a Self-incompatibility locus F-box protein and its non-self S-RNases. Plant Cell 2013, 25, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.H.; Meng, X.Y.; Kao, T.H. Comparison of Petunia inflata S-locus F-box protein (Pi SLF) with Pi SLF like proteins reveals its unique function in S-RNase based self-incompatibility. Plant Cell 2007, 19, 3593–3609. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Meng, D.; Gu, Z.Y.; Li, W.; Wang, A.D.; Yang, Q.; Zhu, Y.D.; Li, T.Z. A novel gene, MdSSK1, as a component of the SCF complex rather than MdSBP1 can mediate the ubiquitination of S-RNase in apple. J. Exp. Bot. 2014, 65, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Wang, H.Y.; Zhao, L.; Zhou, J.L.; Huang, J.; Zhang, Y.S.; Xue, Y.B. The F-box protein AhSLF-S2 physically interacts with S-RNases that may be inhibited by the ubiquitin/26S proteasome pathway of protein degradation during compatible pollination in Antirrhinum. Plant Cell 2004, 16, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.H.; Jiang, N.; Xing, W.; Li, T.L.; Yuan, D.; Li, W.T.; Li, J.T.; Luo, L. Cloning and tissue-specific expression analysis of hepcidin gene in koi (Cyprinus carpio). Microbiol. china 2017, 44, 325–335. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Chen, J.G.; Zhang, Y.Q.; Wang, C.P.; Lü, W.T.; Jin, J.B.; Hua, X.J. Proline induces calcium-mediated oxidative burst and salicylic acid signaling. Amino Acids 2011, 40, 1473–1484. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein Length (aa) | Molecular Weight (Da) | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity |
|---|---|---|---|---|---|---|
| S2-LbSLF1 | 389 | 44,852.75 | 4.81 | 49.36 | 93.88 | −0.063 |
| S2-LbSLF2 | 389 | 44,951.82 | 5.28 | 40.67 | 89.85 | −0.076 |
| S2-LbSLF3 | 393 | 45,512.54 | 5.74 | 41.11 | 94.22 | −0.059 |
| S2-LbSLF4 | 393 | 45,805.57 | 5.12 | 46.43 | 89.26 | −0.183 |
| S2-LbSLF5 | 389 | 44,812.70 | 4.84 | 46.66 | 92.11 | −0.060 |
| S2-LbSLF6 | 390 | 45,250.00 | 5.38 | 39.44 | 91.38 | −0.105 |
| S2-LbSLF7 | 390 | 45,187.04 | 5.92 | 34.02 | 90.38 | −0.111 |
| S2-LbSLF8 | 390 | 44,949.81 | 5.07 | 42.69 | 99.41 | −0.007 |
| S2-LbSLF9 | 358 | 42,018.48 | 7.07 | 43.74 | 87.35 | −0.248 |
| S2-LbSLF10 | 388 | 45,029.85 | 7.06 | 42.38 | 86.34 | −0.217 |
| S2-LbSLF11 | 391 | 45,278.55 | 6.32 | 47.75 | 91.69 | −0.044 |
| S2-LbSLF12 | 388 | 45,351.63 | 5.23 | 46.10 | 93.87 | −0.059 |
| S2-LbSLF13 | 388 | 45,351.63 | 5.23 | 46.10 | 93.87 | −0.059 |
| Gene Name | Subcellular Location | Alpha Helix/% | Extended Strand/% | Beta Turn/% | Random Coil/% |
|---|---|---|---|---|---|
| S2-LbSLF1 | Nucleus | 17.48 | 32.13 | 3.08 | 47.30 |
| S2-LbSLF2 | Nucleus | 18.25 | 30.59 | 4.11 | 47.04 |
| S2-LbSLF3 | Nucleus | 15.01 | 31.30 | 5.09 | 48.60 |
| S2-LbSLF4 | Nucleus | 19.08 | 30.03 | 3.82 | 47.07 |
| S2-LbSLF5 | Nucleus | 16.97 | 32.65 | 4.11 | 46.27 |
| S2-LbSLF6 | Nucleus, Cell membrane | 17.18 | 30.51 | 3.59 | 48.72 |
| S2-LbSLF7 | Nucleus | 19.23 | 29.74 | 4.10 | 46.92 |
| S2-LbSLF8 | Nucleus | 17.44 | 30.00 | 4.62 | 47.95 |
| S2-LbSLF9 | Nucleus | 16.20 | 30.73 | 4.47 | 48.60 |
| S2-LbSLF10 | Nucleus | 18.56 | 28.09 | 4.90 | 48.45 |
| S2-LbSLF11 | Nucleus, Cell membrane, Chloroplast | 18.16 | 30.18 | 4.09 | 47.57 |
| S2-LbSLF12 | Nucleus | 17.01 | 30.67 | 4.64 | 47.68 |
| S2-LbSLF13 | Nucleus | 15.27 | 30.28 | 4.07 | 50.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Nan, X.; Zhang, X.; Xu, W.; Ma, H.; Yang, Z.; Wang, C. The Identification and Analysis of the Self-Incompatibility Pollen Determinant Factor SLF in Lycium barbarum. Plants 2024, 13, 959. https://doi.org/10.3390/plants13070959
Wu J, Nan X, Zhang X, Xu W, Ma H, Yang Z, Wang C. The Identification and Analysis of the Self-Incompatibility Pollen Determinant Factor SLF in Lycium barbarum. Plants. 2024; 13(7):959. https://doi.org/10.3390/plants13070959
Chicago/Turabian StyleWu, Jiali, Xiongxiong Nan, Xin Zhang, Wendi Xu, Haijun Ma, Zijun Yang, and Cuiping Wang. 2024. "The Identification and Analysis of the Self-Incompatibility Pollen Determinant Factor SLF in Lycium barbarum" Plants 13, no. 7: 959. https://doi.org/10.3390/plants13070959
APA StyleWu, J., Nan, X., Zhang, X., Xu, W., Ma, H., Yang, Z., & Wang, C. (2024). The Identification and Analysis of the Self-Incompatibility Pollen Determinant Factor SLF in Lycium barbarum. Plants, 13(7), 959. https://doi.org/10.3390/plants13070959