
Unveiling the Genetic Tapestry: Exploring Rhizoctonia solani AG-3 Anastomosis Groups in Potato Crops across Borders

 ,
,  ,
,  , ,
, , 
 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
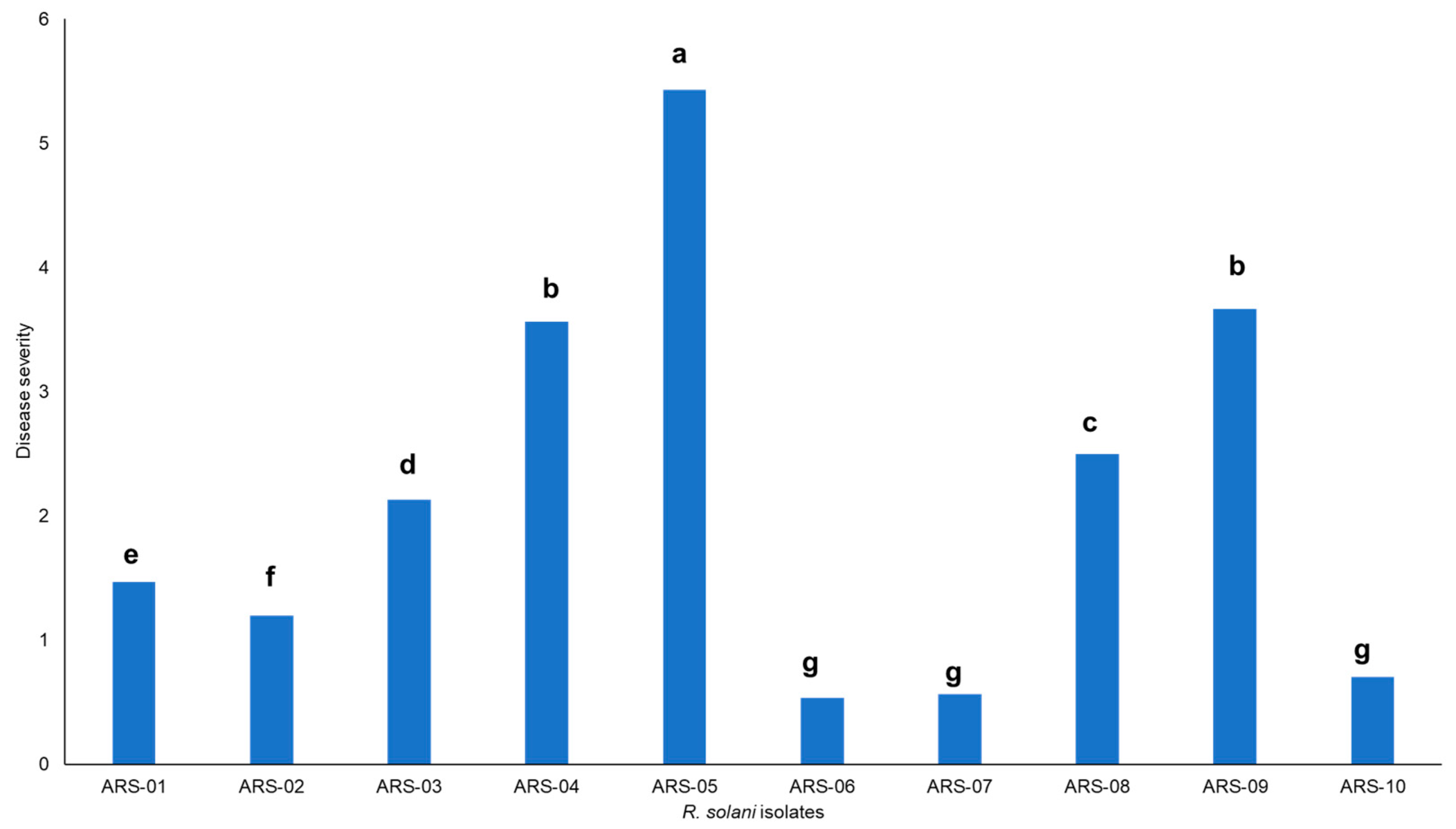
2.1. Isolation, Identification and Pathogenicity of Rhizoctonia Solani Isolates
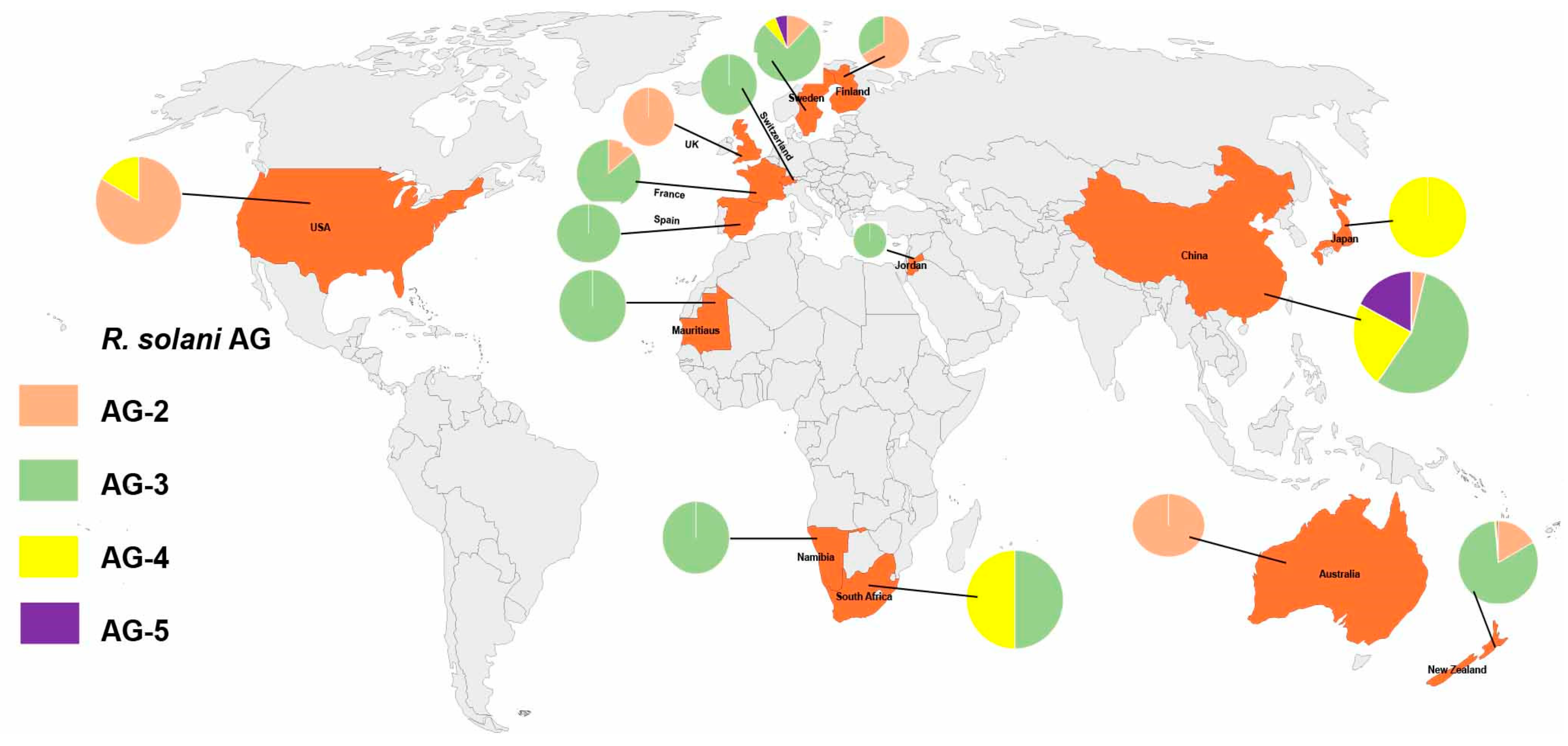
2.2. Distribution of Anastomosis Groups
2.3. Anastomosis Groups Based on Geographical Region: Number of Studies Reported
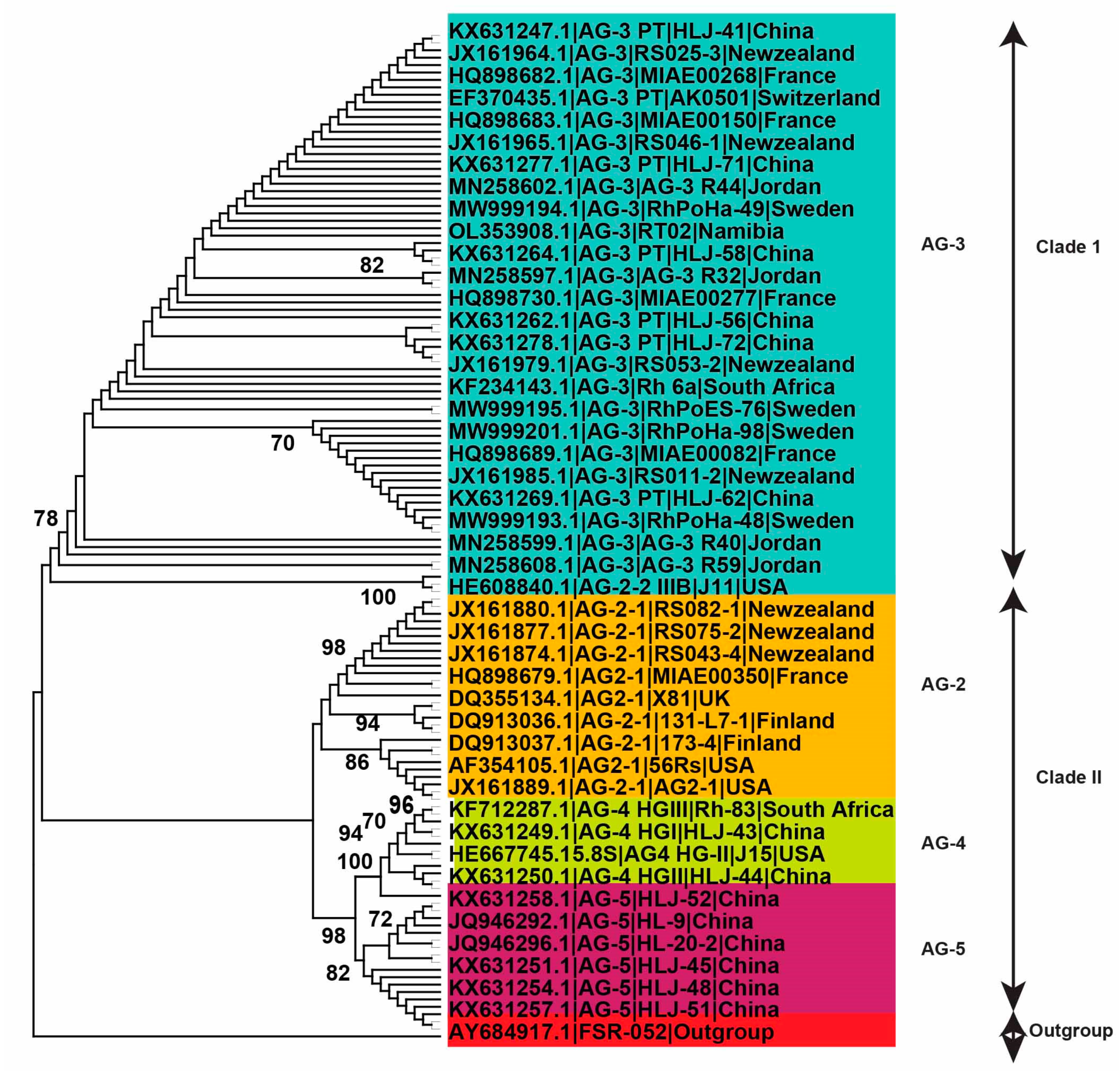
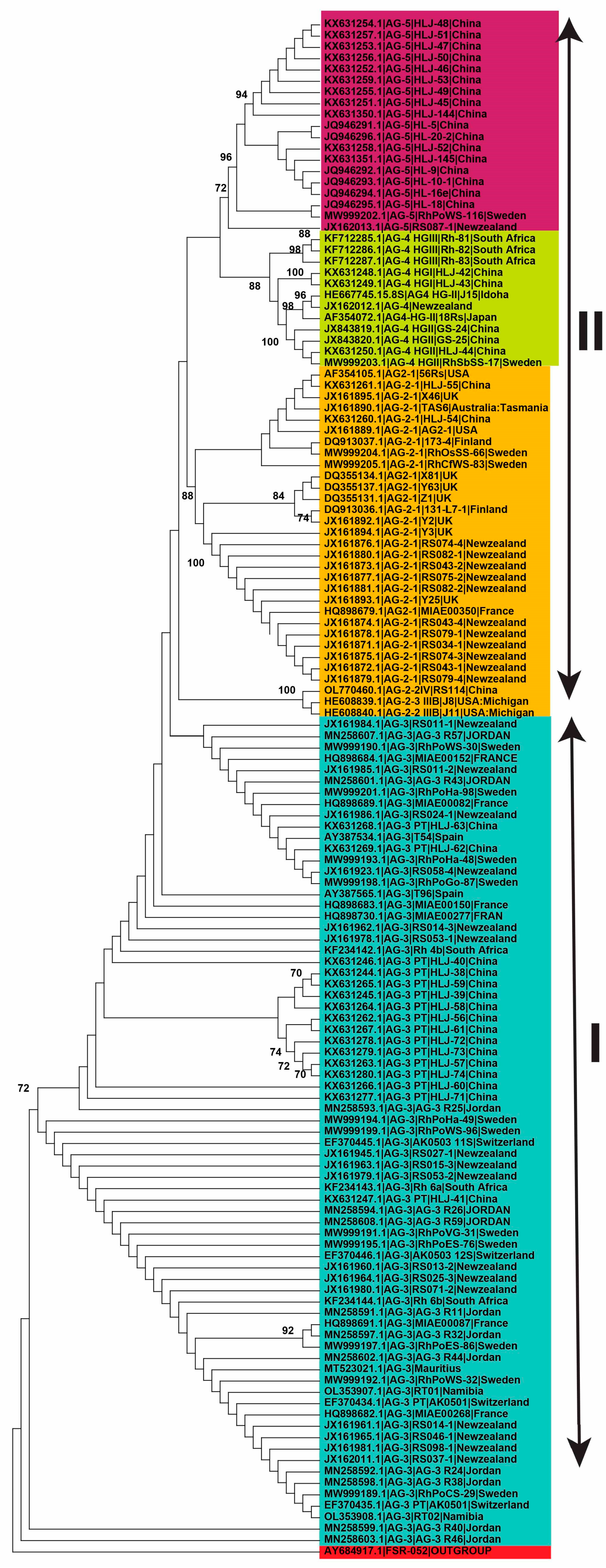
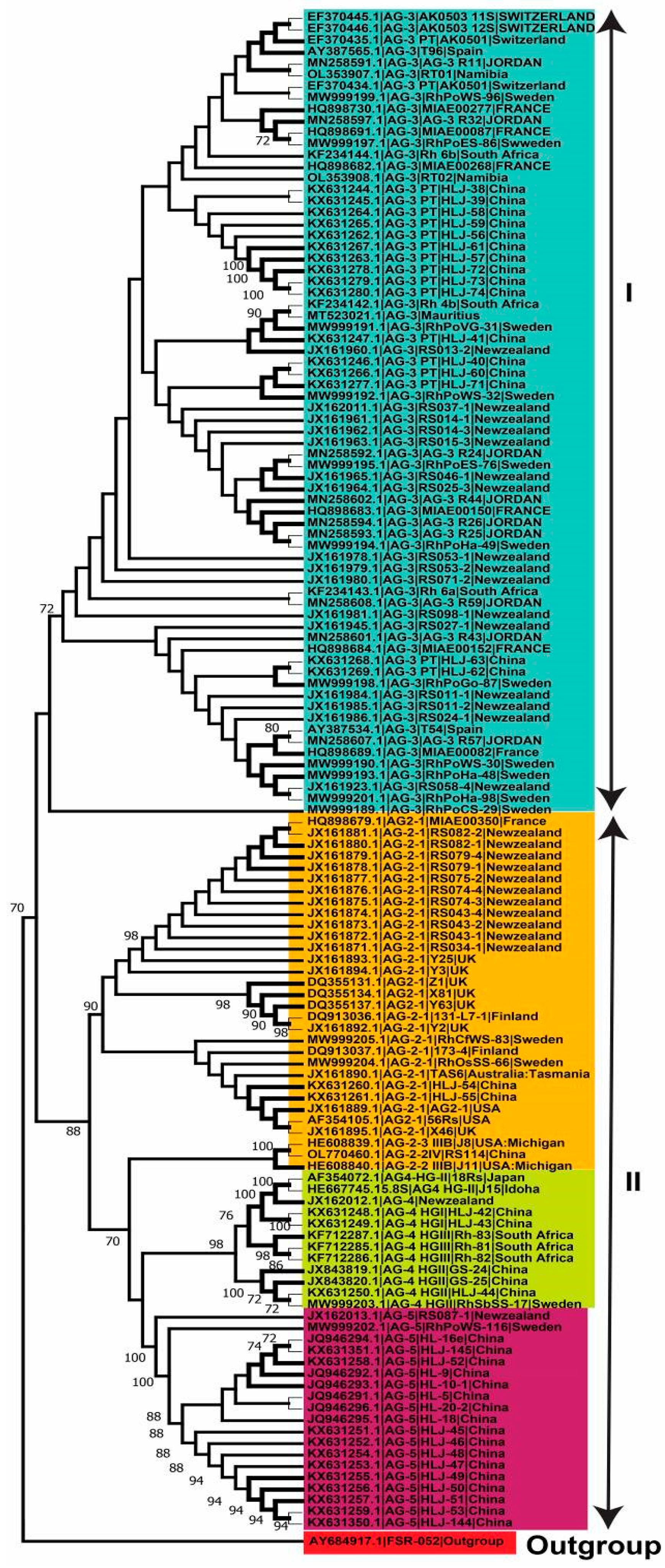
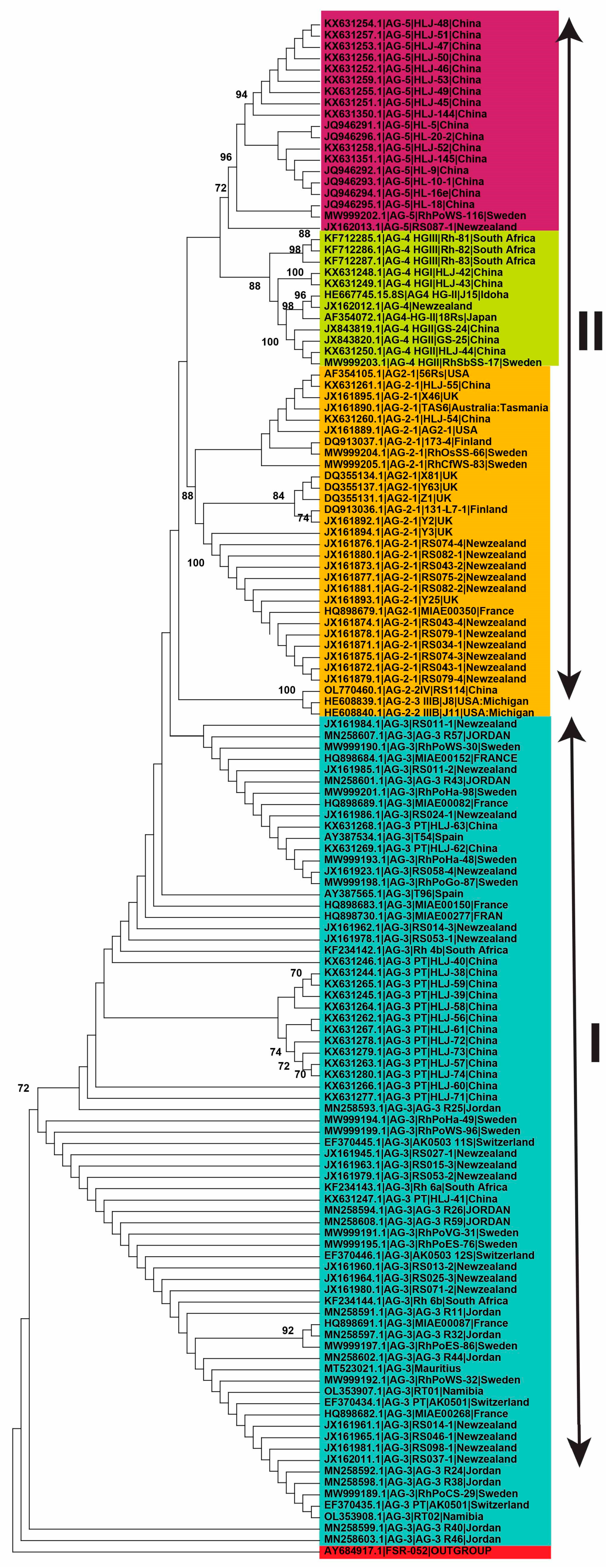
2.4. Genetic Diversity of Anastomosis Groups
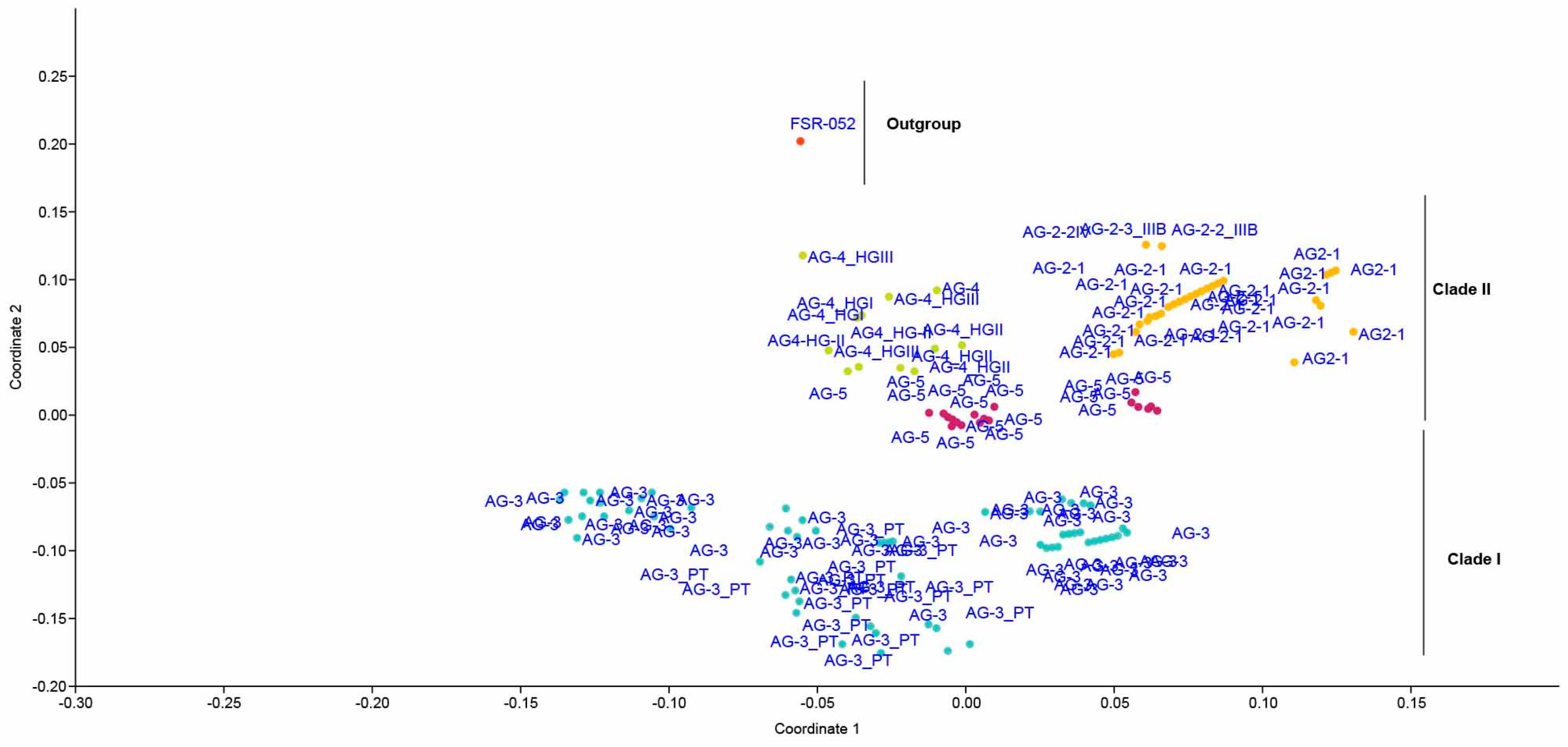
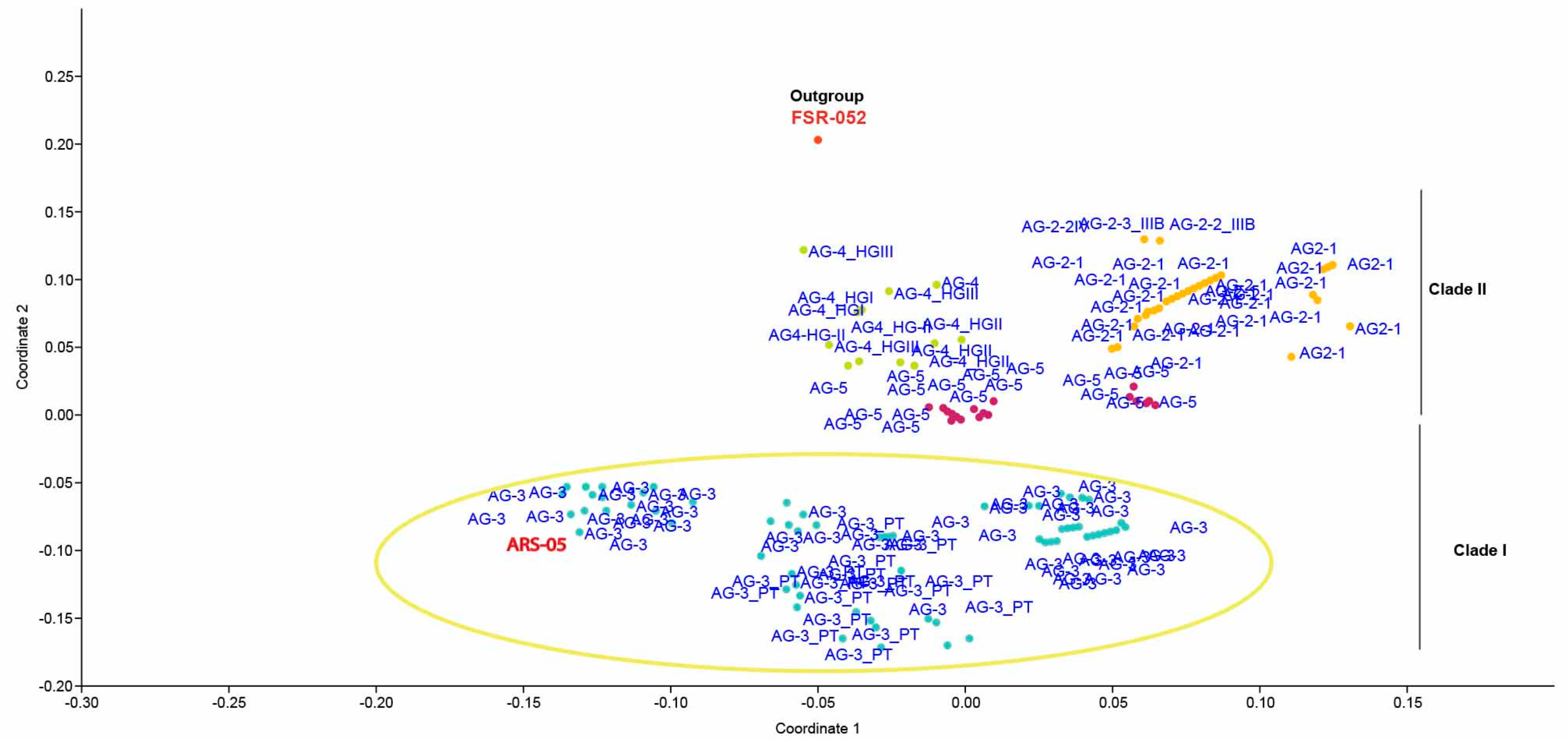
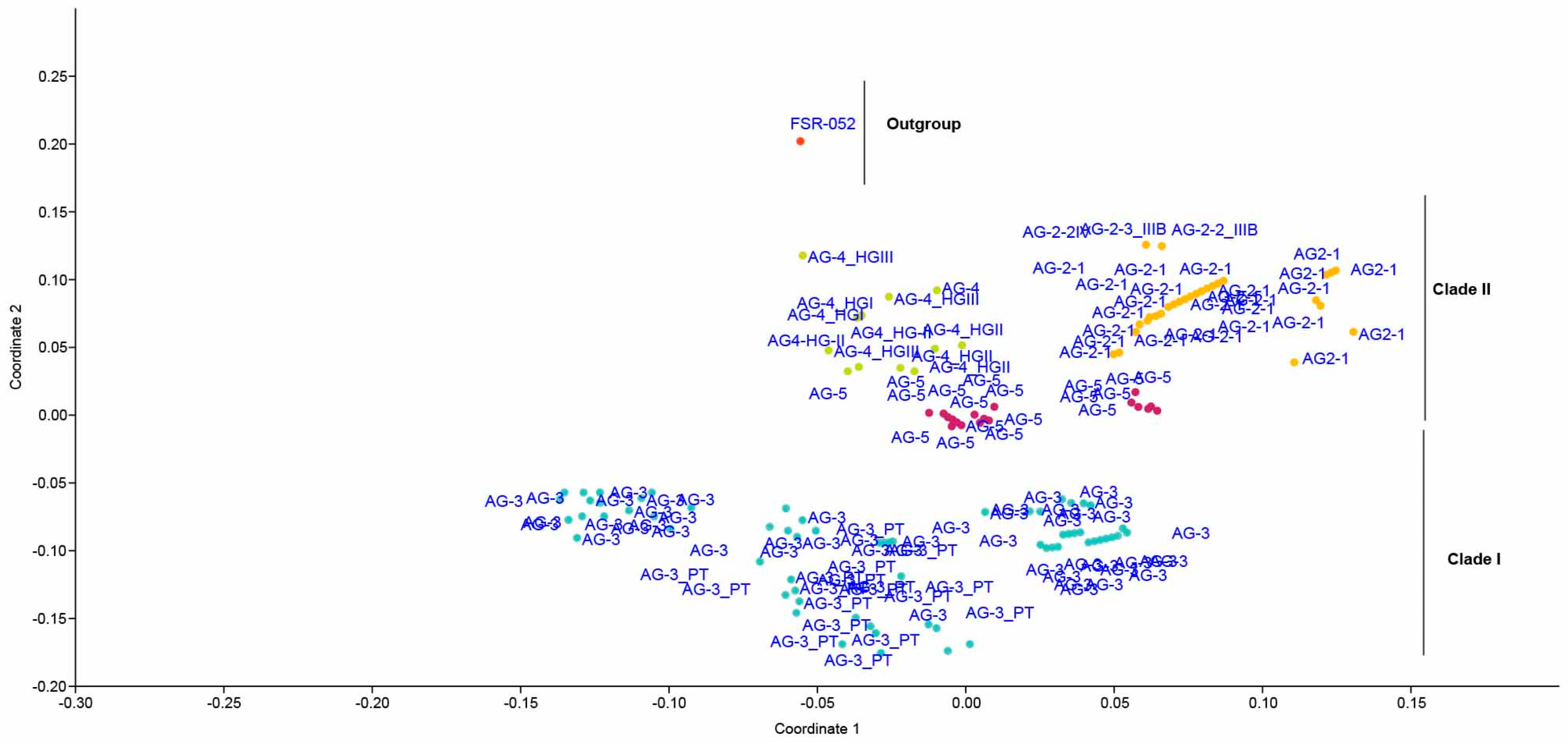
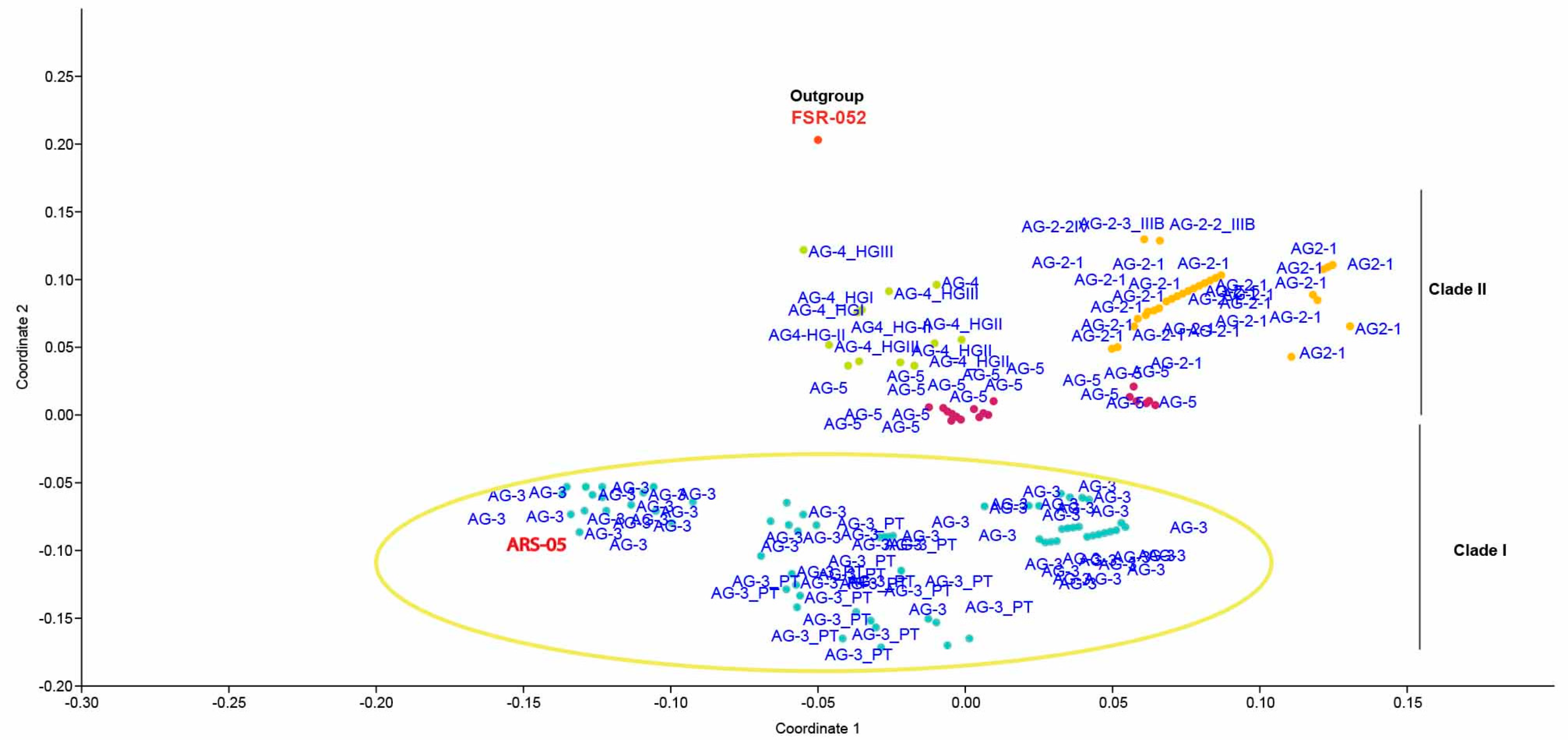
2.5. Genetic Relatedness among and within Clades and Subclades Representing Anastomosis Groups
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Fungal Isolation and Identification
4.2. Pathogenicity Test
4.3. Molecular Identification
4.4. Data Collection for Assessment of Anastomosis Groups (AGs) Associated with Potato
4.5. Phylogenetic Analysis
4.6. Principal Coordinate Analysis (PCoA) and Sequence Similarities
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brar, A.; Bhatia, A.K.; Pandey, V.; Kumari, P. Biochemical and phytochemical properties of potato: A Review. Chem. Sci. Rev. Lett. 2017, 6, 117–129. [Google Scholar]
- Dolničar, P. Importance of potato as a crop and practical approaches to potato breeding. Methods Protoc. 2021, 2354, 3–20. [Google Scholar]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database; FAO: Rome, Italy, 2023. [Google Scholar]
- Senapati, M.; Tiwari, A.; Sharma, N.; Chandra, P.; Bashyal, B.M.; Ellur, R.K.; Bhowmick, P.K.; Bollinedi, H.; Vinod, K.K.; Singh, A.K.; et al. Rhizoctonia solani Kühn Pathophysiology: Status and Prospects of Sheath Blight Disease Management in Rice. Front. Plant Sci. 2022, 13, 881116. [Google Scholar] [CrossRef]
- Jaradat, Z.; Aldakil, H.; Tadros, M.; Alboom, M.; Khataybeh, B. Rhizoctonia solani AG-3PT is the major pathogen associated with potato stem canker and black scurf in Jordan. AIMS Agric. Food 2023, 8, 119–136. [Google Scholar] [CrossRef]
- Li, Y.; Yang, N.; Mu, T.; Wu, X.; Zhao, C. Diversity of Mycoviruses Present in Strains of Binucleate Rhizoctonia and Multinucleate Rhizoctonia, Causal Agents for Potato Stem Canker or Black Scurf. J. Fungi 2023, 9, 214. [Google Scholar] [CrossRef]
- López-Corrales, R.; Michereff, S.J.; Garcia-Estrada, R.S.; Leon-Felix, J.; Correia, K.C.; Rabago-Zavala, K.; Tovar-Pedraza, J.M. Confirmed Report of Rhizoctonia solani AG-7 Causing Potato Stem Canker in Mexico. Plant Dis. 2023, 107, 2260. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Shah, F.; Butler, R.; Falloon, R.; Stewart, A.; Raikar, S.; Pitman, A.R. Genetic variability and pathogenicity of Rhizoctonia solani associated with black scurf of potato in New Zealand. Plant Pathol. 2014, 63, 651–666. [Google Scholar] [CrossRef]
- Iradukunda, L.; Wang, Y.-P.; Nkurikiyimfura, O.; Wang, T.; Yang, L.-N.; Zhan, J. Establishment and Application of a Multiplex PCR Assay for the Rapid Detection of Rhizoctonia solani Anastomosis Group (AG)-3PT, the Pathogen Causing Potato Black Scurf and Stem Canker. Pathogens 2022, 11, 627. [Google Scholar] [CrossRef]
- Atkinson, D.; Thornton, M.K.; Miller, J.S. Development of Rhizoctonia solani on stems, stolons and tubers of potatoes I. Effect of inoculum source. Am. J. Potato Res. 2010, 87, 374–381. [Google Scholar] [CrossRef]
- Salamone, A.L.; Okubara, P.A. Real-time PCR quantification of Rhizoctonia solani AG-3 from soil samples. J. Microbiol. Methods 2020, 172, 105914. [Google Scholar] [CrossRef]
- Dixon, G.R.; Tilston, E.L. Soil-borne pathogens and their interactions with the soil environment. In Soil Microbiology and Sustainable Crop Production; Springer: Dordrecht, the Netherlands, 2010; pp. 197–271. [Google Scholar]
- Tariq, M.; Yasmin, S.; Hafeez, F.Y. Biological control of potato black scurf by rhizosphere associated bacteria. Braz. J. Microbiol. 2010, 41, 439–451. [Google Scholar] [CrossRef]
- Tsror, L. Biology, epidemiology and management of Rhizoctonia solani on potato. J. Phytopathol. 2010, 158, 649–658. [Google Scholar] [CrossRef]
- Hussain, T.; Khan, A.A. Bacillus subtilis HussainT-AMU and its Antifungal activity against Potato Black scurf caused by Rhizoctonia solani on seed tubers. Biocatal. Agric. Biotechnol. 2020, 23, 101443. [Google Scholar] [CrossRef]
- Ogoshi, A. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kuhn. Annual review of phytopathology 1987, 25, 125–143. [Google Scholar] [CrossRef]
- Tsror, L. Fungal, oomycete, and plasmodiophorid diseases of potato and their control. In Potato Production Worldwide; Academic Press: Cambridge, MA, USA, 2023; pp. 145–178. [Google Scholar]
- Zhang, X.; Li, D.; Huo, H.; Xing, X.; Lian, Y.; Yu, Z.; Hao, J. Improving evaluation of potato resistance to Rhizoctonia solani infection by optimizing inoculum-based method combined with toxin-based assay. Crop Prot. 2021, 144, 105544. [Google Scholar] [CrossRef]
- Le Cointe, R.; Simon, T.E.; Delarue, P.; Hervé, M.; Leclerc, M.; Poggi, S. Reducing the Use of Pesticides with Site-Specific Application: The Chemical Control of Rhizoctonia solani as a Case of Study for the Management of Soil-Borne Diseases. PLoS ONE 2016, 11, e0163221. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, L.; Dong, Y.; Chen, W.; Li, C.; Gao, X.; Chen, R.; Li, L.; Xu, Z. The antagonistic mechanism of Bacillus velezensis ZW10 against rice blast disease: Evaluation of ZW10 as a potential biopesticide. PLoS ONE 2021, 16, e0256807. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Yu, X.X.; Yu, Z.; Xue, Y.F.; Qi, L.P. A simple method based on laboratory inoculum and field inoculum for evaluating potato resistance to black scurf caused by Rhizoctonia solani. Breed Sci. 2014, 64, 156–163. [Google Scholar] [CrossRef]
- Abbas, A.; Ali, A.; Hussain, A.; Ali, A.; Alrefaei, A.F.; Naqvi, S.A.H.; Rao, M.J.; Mubeen, I.; Farooq, T.; Ölmez, F.; et al. Assessment of Genetic Variability and Evolutionary Relationships of Rhizoctonia solani Inherent in Legume Crops. Plants 2023, 12, 2515. [Google Scholar] [CrossRef] [PubMed]
- Muzhinji, N.; Woodhall, J.W.; Truter, M.; van der Waals, J.E. Population genetic structure of Rhizoctonia solani AG 3-PT from potatoes in South Africa. Fungal Biol. 2016, 120, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Liu, L.M.; Hou, Y.X.; Li, L.; Huang, S.W. Pathotypic and genetic diversity in the population of Rhizoctonia solani AG 1-IA causing rice sheath blight in China. Plant Pathol. 2015, 64, 718–728. [Google Scholar] [CrossRef]
- Das, S.; Plyler-Harveson, T.; Santra, D.K.; Maharjan, B.; Nielson, K.A.; Harveson, R.M. A longitudinal study on morpho-genetic diversity of pathogenic Rhizoctonia solani from sugar beet and dry beans of western Nebraska. BMC Microbiol. 2020, 20, 354. [Google Scholar] [CrossRef] [PubMed]
- Erper, I.; Ozer, G.; Kalendar, R.; Avci, S.; Yildirim, E.; Alkan, M.; Turkkan, M. Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey). J. Fungi 2021, 7, 234. [Google Scholar] [CrossRef]
- Keijer, J.; Houterman, P.M.; Dullemans, A.M.; Korsman, M.G. Heterogeneity in electrophoretic karyotype within and between anastomosis groups of Rhizoctonia solani. Mycol. Res. 1996, 100, 789–797. [Google Scholar] [CrossRef]
- Yanar, Y.; Yılmaz, G.; Cesmeli, I.; Coskun, S. Characterization of Rhizoctonia solani isolates from potatoes in turkey and screening potato cultivars for resistance to AG-3 isolates. Phytoparasitica 2005, 33, 370–376. [Google Scholar] [CrossRef]
- Francis, A.; Ghosh, S.; Tyagi, K.; Prakasam, V.; Rani, M.; Singh, N.P.; Pradhan, A.; Sundaram, R.M.; Priyanka, C.; Laha, G.S.; et al. Evolution of pathogenicity-associated genes in Rhizoctonia solani AG1-IA by genome duplication and transposon-mediated gene function alterations. BMC Biol. 2023, 21, 15. [Google Scholar] [CrossRef]
- Das, S. Rhizoctonia solani on Potato in New Zealand: Pathogen Characterisation and Identification of Double-Stranded RNA Viruses that May Affect Their Virulence. Ph.D. Dissertation, Lincoln University, Oxford, PA, USA, 2013. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Ferrucho, R.L.; Cifuentes, J.M.; Ceresini, P.; García-Domínguez, C. Rhizoctonia solani AG-3PT is the major pathogen associated with potato stem canker and black scurf in Colombia. Agron. Colomb. 2012, 30, 204–213. [Google Scholar]
- Tjimune, R.; Mangwende, E.; Lekota, M.; Muzhinji, N. First Report of Rhizoctonia solani AG 3-PT causing black scurf on potato tubers in Namibia. New Dis. Rep. 2022, 45, e12066. [Google Scholar] [CrossRef]
- Johnk, J.S.; Jones, R.K.; Shew, H.D.; Carling, D.E. Characterization of populations of Rhizoctonia solani AG-3 from potato and tobacco. Phytopathology 1993, 83, 854–858. [Google Scholar] [CrossRef]
- Jing, Y.; Li, X.N.; Yu, J.F. Anastomosis groups of Rhizoctonia solani from cotton in northern China. Mycosystem 2012, 31, 540–547. [Google Scholar]
- Lehtonen, M. Rhizoctonia solani as a Potato Pathogen–Variation of Isolates in Finland and Host Response. Ph.D. Thesis, Viikki Graduate School in Molecular Biosciences, University of Helsinki, Helsinki, Finland, 2009. [Google Scholar]
- Zimudzi, J.; Coutinho, T.A.; Van der Waals, J.E. Pathogenicity of fungi isolated from atypical skin blemishes on potatoes in South Africa and Zimbabwe. Potato Res. 2017, 60, 119–144. [Google Scholar] [CrossRef]
- Djébali, N.; Elkahoui, S.; Taamalli, W.; Hessini, K.; Tarhouni, B.; Mrabet, M. Tunisian Rhizoctonia solani AG3 strains affect potato shoot macronutrients content, infect faba bean plants and show in vitro resistance to azoxystrobin. Australas. Plant Pathol. 2014, 43, 347–358. [Google Scholar] [CrossRef]
- Lehtonen, M.J.; Wilson, P.S.; Ahvenniemi, P.; Valkonen, J. Formation of canker lesions on stems and black scurf on tubers in experimentally inoculated potato plants by isolates of AG2-1, AG3 and AG5 of Rhizoctonia solani: A pilot study and literature review. Agric. Food Sci. 2009, 18, 223–233. [Google Scholar] [CrossRef]
- Woodhall, J.W. Characterisation of Rhizoctonia solani Anastomosis Groups and Their Pathogenicity to Potato. Ph.D. Thesis, The Open University, Milton Keynes, UK, 2005. [Google Scholar] [CrossRef]
- Ceresini, P.C.; Shew, H.D.; Vilgalys, R.J.; Rosewich, U.L.; Cubeta, M.A. Genetic structure of populations of Rhizoctonia solani AG-3 on potato in eastern North Carolina. Mycologia 2002, 94, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Fang, X.; Iqbal, S.; Naqvi, S.A.H.; Mehmood, Y.; Rao, M.J.; Hassan, Z.; Ortiz, R.M.; Baazeem, A.; Moustafa, M.; et al. Population Genetics and Anastomosis Group’s Geographical Distribution of Rhizoctonia solani Associated with Soybean. Genes 2022, 13, 2417. [Google Scholar] [CrossRef] [PubMed]
- Tziros, G.T.; Karaoglanidis, G.S. Molecular identification and pathogenicity of Rhizoctonia solani and Pythium spp. associated with damping-off disease on baby leafy vegetables in Greece. Plant Pathol. 2022, 71, 1381–1391. [Google Scholar] [CrossRef]
- Anderson, J.P.; Hane, J.K.; Stoll, T.; Pain, N.; Hastie, M.L.; Kaur, P.; Hoogland, C.; Gorman, J.J.; Singh, K.B. Proteomic Analysis of Rhizoctonia solani Identifies Infection-specific, Redox Associated Proteins and Insight into Adaptation to Different Plant Hosts. Mol. Cell. Proteom. 2016, 15, 1188–1203. [Google Scholar] [CrossRef]
- Hejazi, R.; Esfahani, M.N.; Maleki, M.; Sedaghatfar, E. Susceptibility assessment and genetic population structure associated with Rhizoctonia solani AG3-PT-Potato stem canker disease. Physiol. Mol. Plant Pathol. 2022, 119, 101835. [Google Scholar] [CrossRef]
- Monazzah, M.; Esfahani, M.N.; Enferadi, S.T. Genetic structure and proteomic analysis associated in potato to Rhizoctonia solani AG-3PT-stem canker and black scurf. Physiol. Mol. Plant Pathol. 2022, 122, 101905. [Google Scholar] [CrossRef]
- Frank, J.A.; Leach, S.S. Comparison of tuberborne and soilborne inoculum in the Rhizoctonia disease of potato. Phytopathology 1980, 70, 1–53. [Google Scholar] [CrossRef]
- Errampalli, D.; Johnston, H.W. Control of tuber-borne black scurf [Rhizoctonia solani] and common scab [Streptomyces scabies] of potatoes with a combination of sodium hypochlorite and thiophanate-methyl preplanting seed tuber treatment. Can. J. Plant Pathol. 2001, 23, 68–77. [Google Scholar] [CrossRef]
- Islam, M.R.; Hossain, M.M. Occurrence and bio-control of stem canker and black scurf (Rhizoctonia solani) of potato in Bangladesh. Indian J. Agric. Sci. 2019, 89, 329–332. Available online: https://epubs.icar.org.in/index.php/IJAgS/article/view/48517 (accessed on 15 August 2018).
- Islam, M.R.; Hossain, M.M. Integrated management of stem canker and black scurf of potato. J. Plant Prot. Res. 2018, 58, 361–369. [Google Scholar]
- Islam, M.R.; Hossain, M.M. Effect of organic amendment on stem canker and black scurf diseases of potato. Biosci. J. 2018, 34, 1415–1423. [Google Scholar]
- Sharma, B.P. Participatory black scurf disease management on potato in Nepal. Nepal Agric. Res. J. 2007, 8, 56–62. [Google Scholar] [CrossRef]
- Rabbani, M.; Moslem, M.A.; Hadi, S. Molecular Characterization of Anastomosis Groups of Rhizoctonia solani Associated with Potato Tubers in Saudi Arabia. J. Pure Appl. Microbiol. 2013, 7, 1789–1798. [Google Scholar]
- Torrance, L.; Talianksy, M.E. Potato Virus Y Emergence and Evolution from the Andes of South America to Become a Major Destructive Pathogen of Potato and Other Solanaceous Crops Worldwide. Viruses 2020, 12, 1430. [Google Scholar] [CrossRef] [PubMed]
- Kanetis, L.; Tsimouris, D.; Christoforou, M. Characterization of Rhizoctonia solani associated with black scurf in Cyprus. Plant Dis. 2016, 100, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.M.; Gerard, E. Suppressive soils for Rhizoctonia diseases of potato in New Zealand. New Zealand Plant Prot. 2022, 75, 31–49. [Google Scholar] [CrossRef]
- Ceresini, P.C.; Shew, H.D.; Vilgalys, R.J.; Gale, L.R.; Cubeta, M.A. Detecting migrants in populations of Rhizoctonia solani anastomosis group 3 from potato in North Carolina using multilocus genotype probabilities. Phytopathology 2003, 93, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Kuninaga, S.; Carling, D.E.; Takeuchi, T.; Yokosawa, R. Comparison of rDNA-ITS sequences between potato and tobacco strains in Rhizoctonia solani AG-3. J. Gen. Plant Pathol. 2000, 66, 2–11. [Google Scholar] [CrossRef]
- Gondal, A.S.; Rauf, A.; Naz, F. Anastomosis Groups of Rhizoctonia solani associated with tomato foot rot in Pothohar Region of Pakistan. Sci Rep. 2019, 9, 3910. [Google Scholar] [CrossRef] [PubMed]
- Salazar, O.; Julián, M.C.; Hyakumachi, M.; Rubio, V. Phylogenetic grouping of cultural types of Rhizoctonia solani AG 2–2 based on ribosomal ITS sequences. Mycologia 2000, 92, 505–509. [Google Scholar] [CrossRef]
- Simons, S.A.; Gilligan, C.A. Relationships between stem canker, stolon canker, black scurf (Rhizoctonia solani) and yield of potato (Solanum tuberosum) under different agronomic conditions. Plant Pathol. 1997, 46, 651–658. [Google Scholar] [CrossRef]
- Zrenner, R.; Genzel, F.; Verwaaijen, B.; Wibberg, D.; Grosch, R. Necrotrophic lifestyle of Rhizoctonia solani AG3-PT during interaction with its host plant potato as revealed by transcriptome analysis. Sci. Rep. 2020, 10, 12574. [Google Scholar] [CrossRef]
- Abbas, A.; Khan, S.U.; Khan, W.U.; Saleh, T.A.; Khan, M.H.U.; Ullah, S.; Ali, A.; Ikram, M. Antagonist effects of strains of Bacillus spp. against Rhizoctonia solani for their protection against several plant diseases: Alternatives to chemical pesticides. C. R. Biol. 2019, 342, 124–135. [Google Scholar] [CrossRef]
- Afridi, M.S.; Ali, S.; Salam, A.; César Terra, W.; Hafeez, A.; Sumaira Ali, B.S.; Al-Tami, M.; Ameen, F.; Ercisli, S.; Marc, R.A.; et al. Plant Microbiome Engineering: Hopes or Hypes. Biology 2022, 11, 1782. [Google Scholar] [CrossRef]
- Chand, T.; Logan, C. Cultural and pathogenic variation in potato isolates of Rhizoctonia solani in Northern Ireland. Trans. Br. Mycol. Soc. 1983, 81, 585–589. [Google Scholar] [CrossRef]
- Anguiz, R.; Martin, C. Anastomosis groups, pathogenicity, and other characteristics of Rhizoctonia solani isolated from potatoes in Peru. Plant Dis. 1989, 73, 199–201. [Google Scholar] [CrossRef]
- Carling, D.E.; Leiner, R.H. Effect of temperature on virulence of Rhizoctonia solani and other Rhizoctonia on potato. Phytopathology 1990, 80, 930–934. [Google Scholar] [CrossRef]
- Virgen-Calleros, G.; Olalde-Portugal, V.; Carling, D.E. Anastomosis groups of Rhizoctonia solani on potato in Central Mexico and potential for biological and chemical control. Am. J. Potato Res. 2000, 77, 219–224. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anastomosis Groups | Subgroups | Sweden | Namibia | Mauritius | Switzerland | China | South Africa | USA | Japan | New Zealand | Finland | France | UK | Spain | Australia | Jordan | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AG-2 | AG-2-1 | 2 | ---- | ---- | ---- | 2 | ---- | 3 | ---- | 24 | 2 | 1 | 3 | ---- | 1 | ---- | 38 |
| ---- | AG-2-2IIIB | ---- | ---- | ---- | ---- | ---- | ---- | 2 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 2 |
| ---- | AG-2-2IV | ---- | ---- | ---- | ---- | 1 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 1 |
| AG-3 | 3 | 13 | 2 | 1 | 2 | ---- | 3 | ---- | ---- | 116 | 1 | 6 | ---- | 2 | ---- | 20 | 166 |
| ---- | AG-3 PT | ---- | ---- | ---- | 2 | 42 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 44 |
| AG-4 | 4 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 1 | ---- | ---- | ---- | ---- | ---- | ---- | 1 |
| ---- | AG-4 HGI | ---- | ---- | ---- | ---- | 12 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 12 |
| ---- | AG-4 HGII | 1 | ---- | ---- | ---- | 5 | ---- | 1 | 1 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 8 |
| ---- | AG-4 HGIII | ---- | ---- | ---- | ---- | ---- | 3 | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | ---- | 3 |
| AG-5 | ---- | 1 | ---- | ---- | ---- | 13 | ---- | ---- | ---- | 1 | ---- | ---- | ---- | ---- | ---- | ---- | 15 |
| Total | ---- | 17 | 2 | 1 | 4 | 75 | 6 | 6 | 1 | 142 | 3 | 7 | 3 | 2 | ---- | 20 | 290 |
| Clade I | Clade II | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AG-3 | AG-2 | AG-4 | AG-5 | |||||||||
| Clades | 3 | 3PT | AG-2-1 | AG-2-2 | 4 | AG-4-I | AG-4-II | AG-4-III | 5 | Outgroup | ||
| Clade I | AG-3 | 3 | 96.5–97 | |||||||||
| 3PT | 84–93.3 | 89.1–100 | ||||||||||
| Clade II | AG-2 | AG-2-1 | 65–90 | 82.6–94 | 98.4–100 | |||||||
| AG-2-2 | 57.8–84 | 79.8–85.5 | 83.6–86 | 97–100 | ||||||||
| AG-4 | 4 | 62.6–86 | 74.8–88.6 | 87.9–88 | 81.3–82.8 | 97–100 | ||||||
| AG-4-I | 62.2–87.1 | 79.2–87.8 | 80–88 | 79.7–83.1 | 87–95.2 | 84–100 | ||||||
| AG-4-II | 74–85.6 | 81.1–86.6 | 80–88 | 79.8–81.9 | 87.2–94 | 96.7–97 | 100 | |||||
| AG-4-III | 62–85 | 68–82.2 | 82.8–88.5 | 75.3–83.6 | 86.1–91.3 | 75–92.7 | 72–90 | 93–100 | ||||
| AG-5 | 5 | 57–90.3 | 83.7–90.5 | 84.8–87.7 | 82.3–85.5 | 81.8–83.3 | 82.6–84.1 | 86.8–87.8 | 76.3–84.3 | 97.8–100 | ||
| Outgroup | 57.1–68.6 | 63.7–69 | 67–68 | 67.4–68.5 | 67.9–68 | 67.3–67.7 | 67.4–67.9 | 62–67 | 65.2–66.3 | 100 * | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naqvi, S.A.H.; Abbas, A.; Farhan, M.; Kiran, R.; Hassan, Z.; Mehmood, Y.; Ali, A.; Ahmed, N.; Hassan, M.Z.; Alrefaei, A.F.; et al. Unveiling the Genetic Tapestry: Exploring Rhizoctonia solani AG-3 Anastomosis Groups in Potato Crops across Borders. Plants 2024, 13, 715. https://doi.org/10.3390/plants13050715
Naqvi SAH, Abbas A, Farhan M, Kiran R, Hassan Z, Mehmood Y, Ali A, Ahmed N, Hassan MZ, Alrefaei AF, et al. Unveiling the Genetic Tapestry: Exploring Rhizoctonia solani AG-3 Anastomosis Groups in Potato Crops across Borders. Plants. 2024; 13(5):715. https://doi.org/10.3390/plants13050715
Chicago/Turabian StyleNaqvi, Syed Atif Hasan, Aqleem Abbas, Muhammad Farhan, Rafia Kiran, Zeshan Hassan, Yasir Mehmood, Amjad Ali, Niaz Ahmed, Muhammad Zeeshan Hassan, Abdulwahed Fahad Alrefaei, and et al. 2024. "Unveiling the Genetic Tapestry: Exploring Rhizoctonia solani AG-3 Anastomosis Groups in Potato Crops across Borders" Plants 13, no. 5: 715. https://doi.org/10.3390/plants13050715
APA StyleNaqvi, S. A. H., Abbas, A., Farhan, M., Kiran, R., Hassan, Z., Mehmood, Y., Ali, A., Ahmed, N., Hassan, M. Z., Alrefaei, A. F., Ölmez, F., Yang, S.-H., & Baloch, F. S. (2024). Unveiling the Genetic Tapestry: Exploring Rhizoctonia solani AG-3 Anastomosis Groups in Potato Crops across Borders. Plants, 13(5), 715. https://doi.org/10.3390/plants13050715