
Integrated Metabolomic and Transcriptomic Profiles Provide Insights into the Mechanisms of Anthocyanin and Carotenoid Biosynthesis in Petals of Medicago sativa ssp. sativa and Medicago sativa ssp. falcata

Abstract
1. Introduction
2. Results
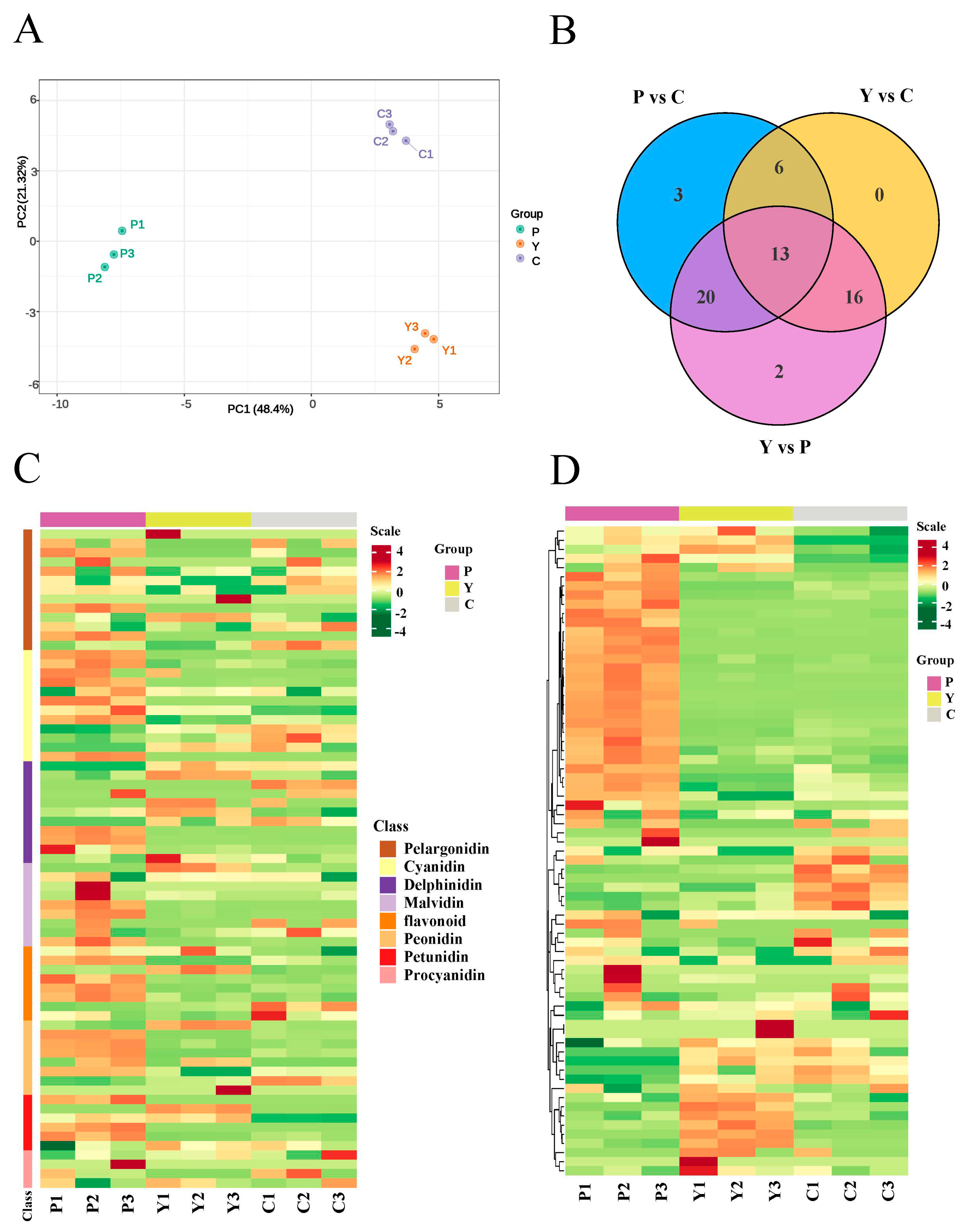
2.1. Targeted Anthocyanin Metabolite Assays of Flowers of Three Different Colors
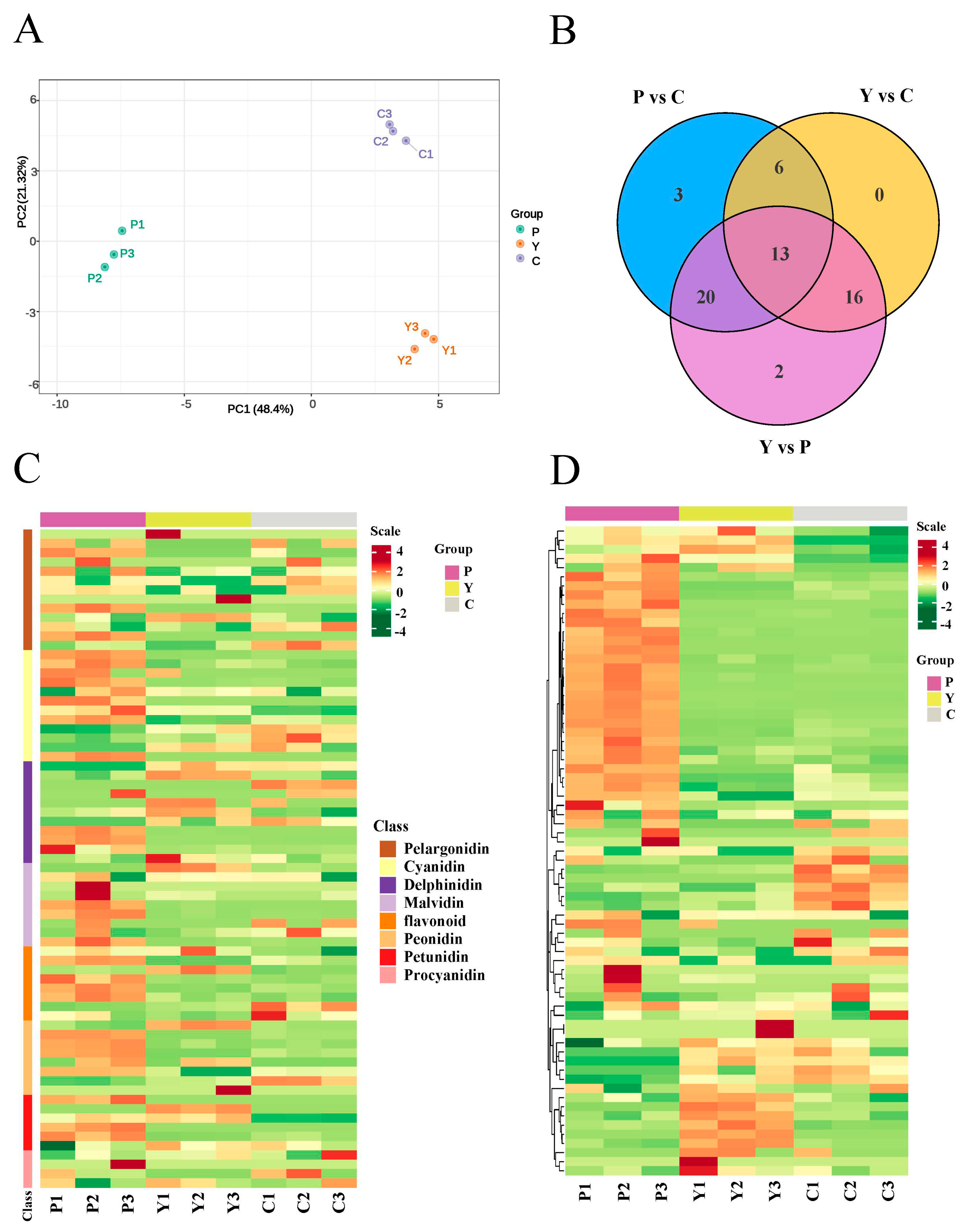
2.2. Anthocyanin Components Accumulated in Purple Flowers and Cream Flowers
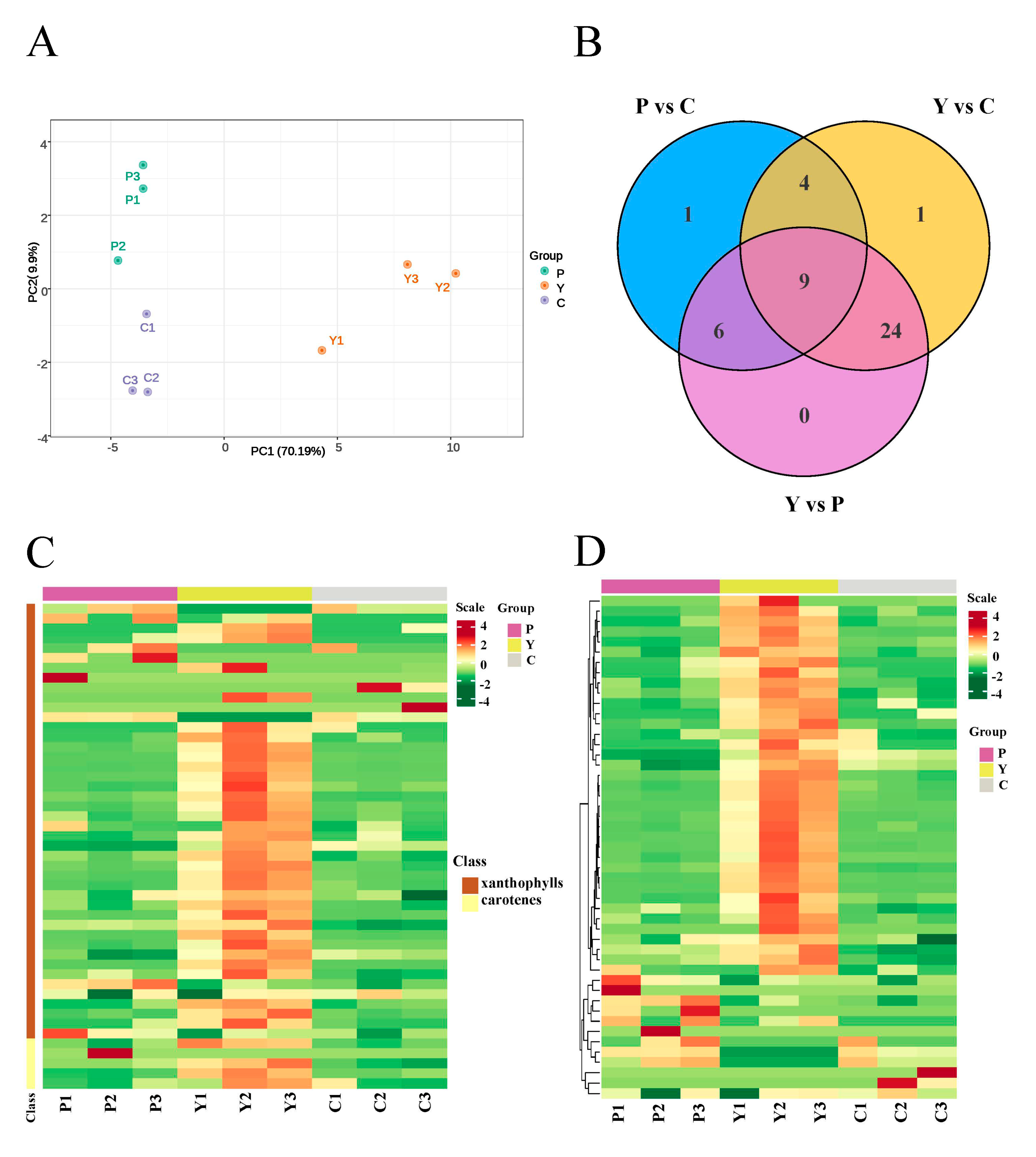
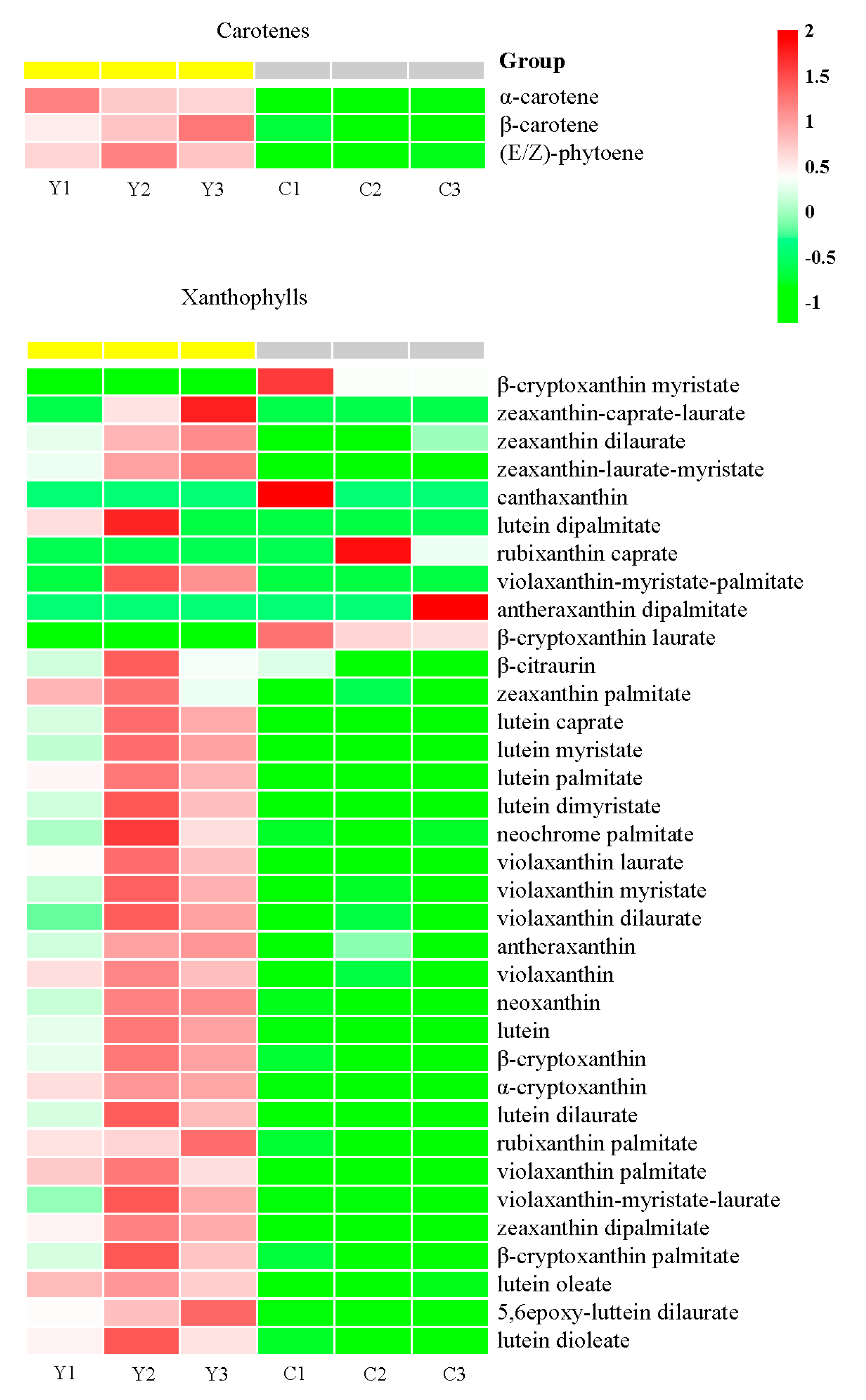
2.3. Targeted Carotenoid Metabolite Assays of Flowers of Three Different Colors
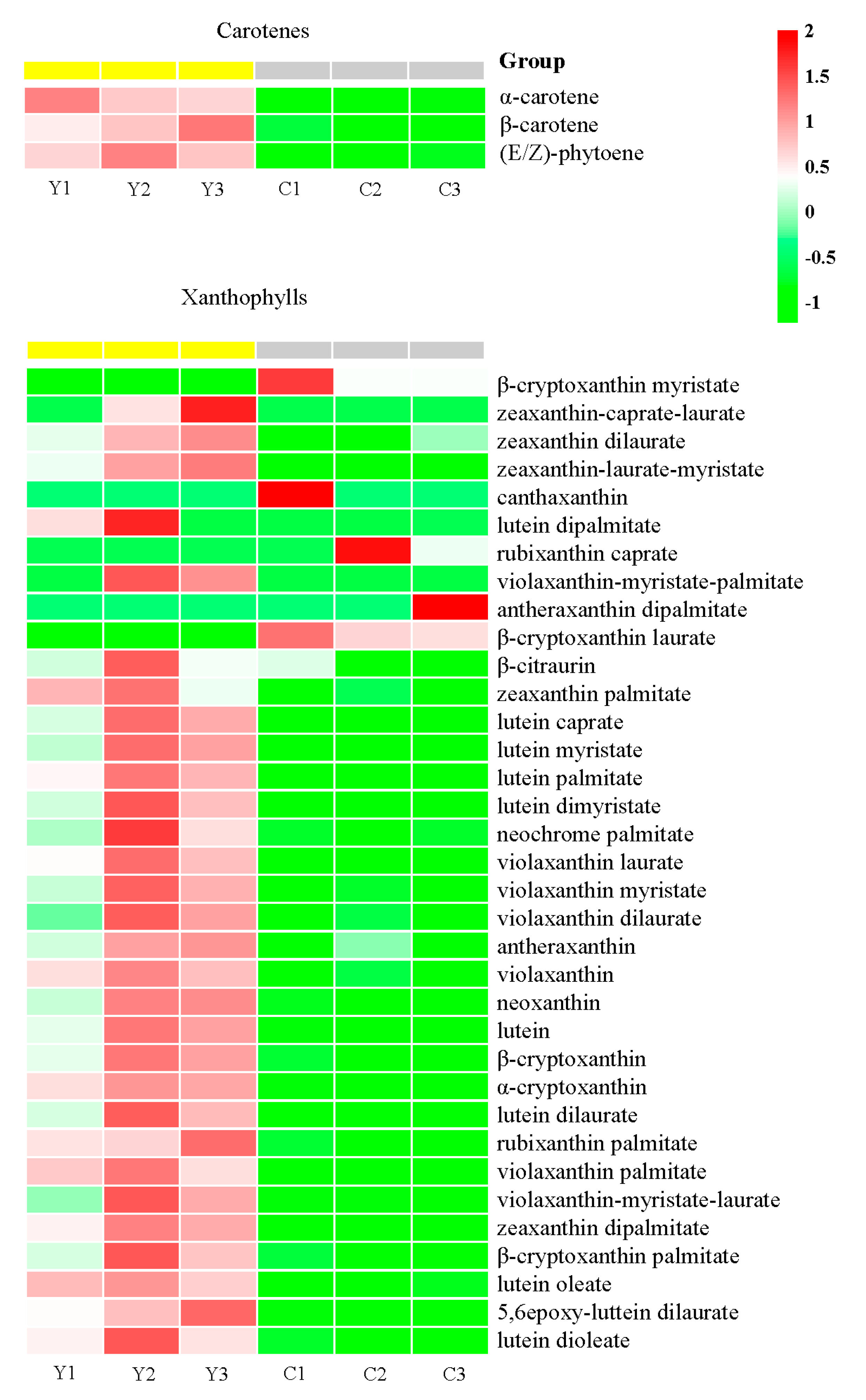
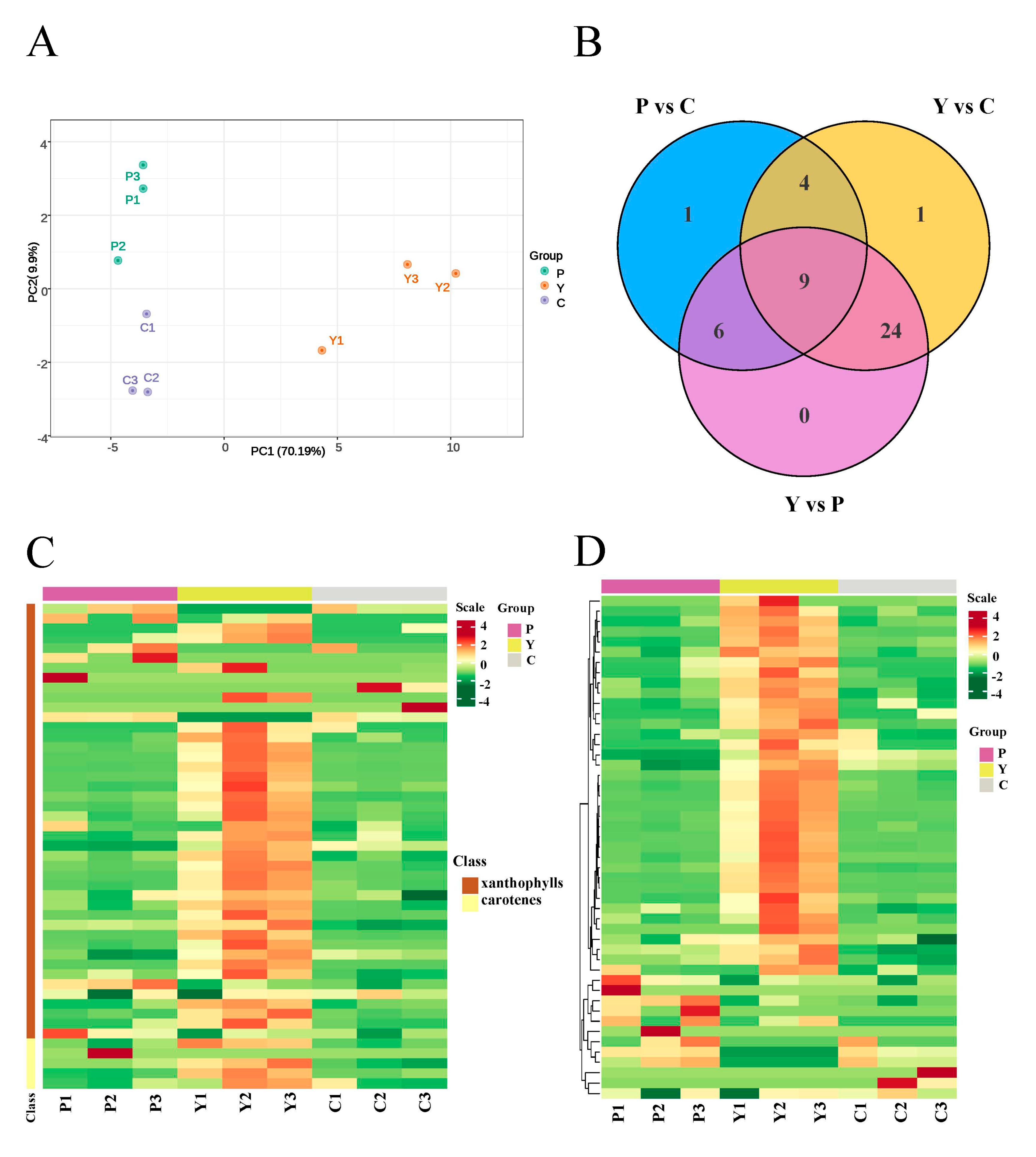
2.4. Identification of Carotenoid Components Involved in the Color Formation of Yellow Flowers
2.5. Transcriptome Analysis of Flower Petals of the Three Different Colors
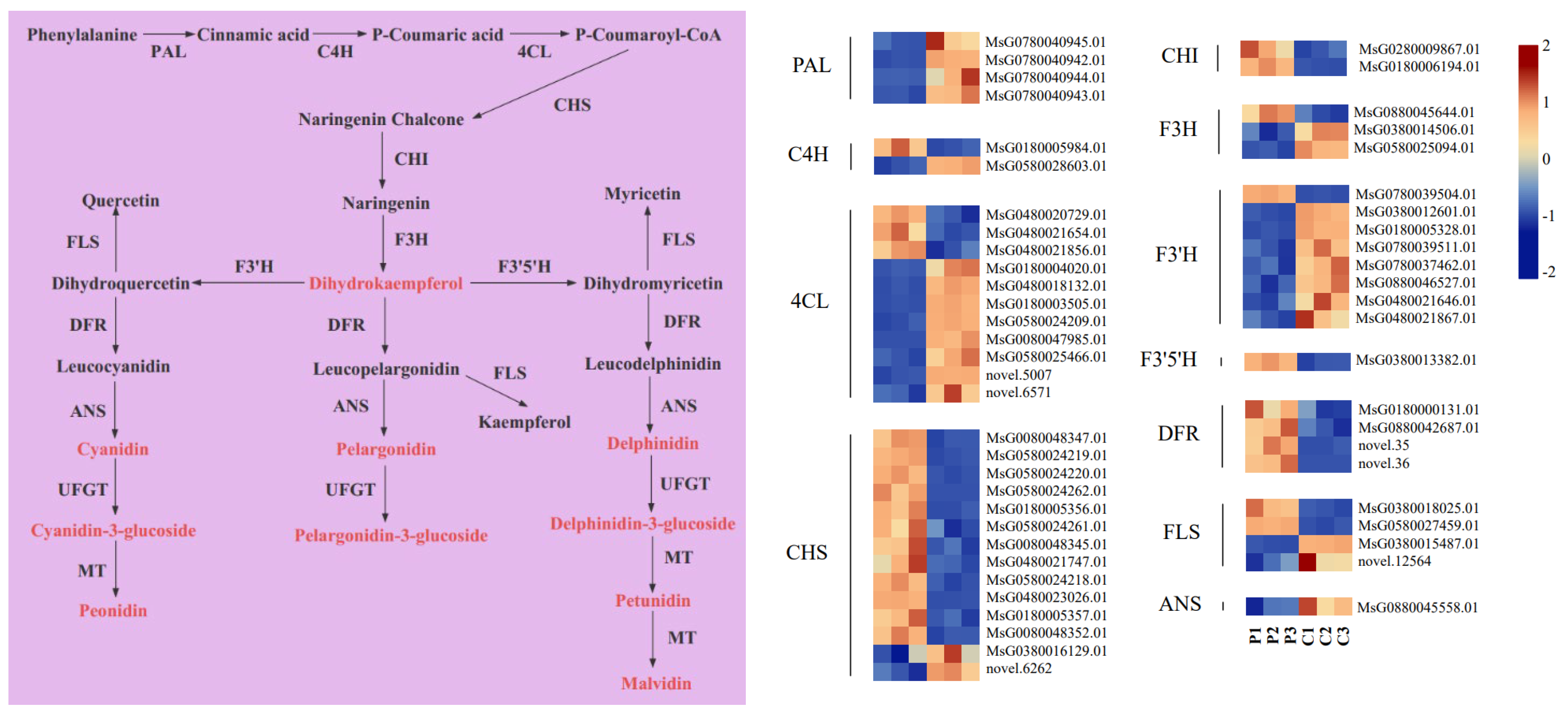
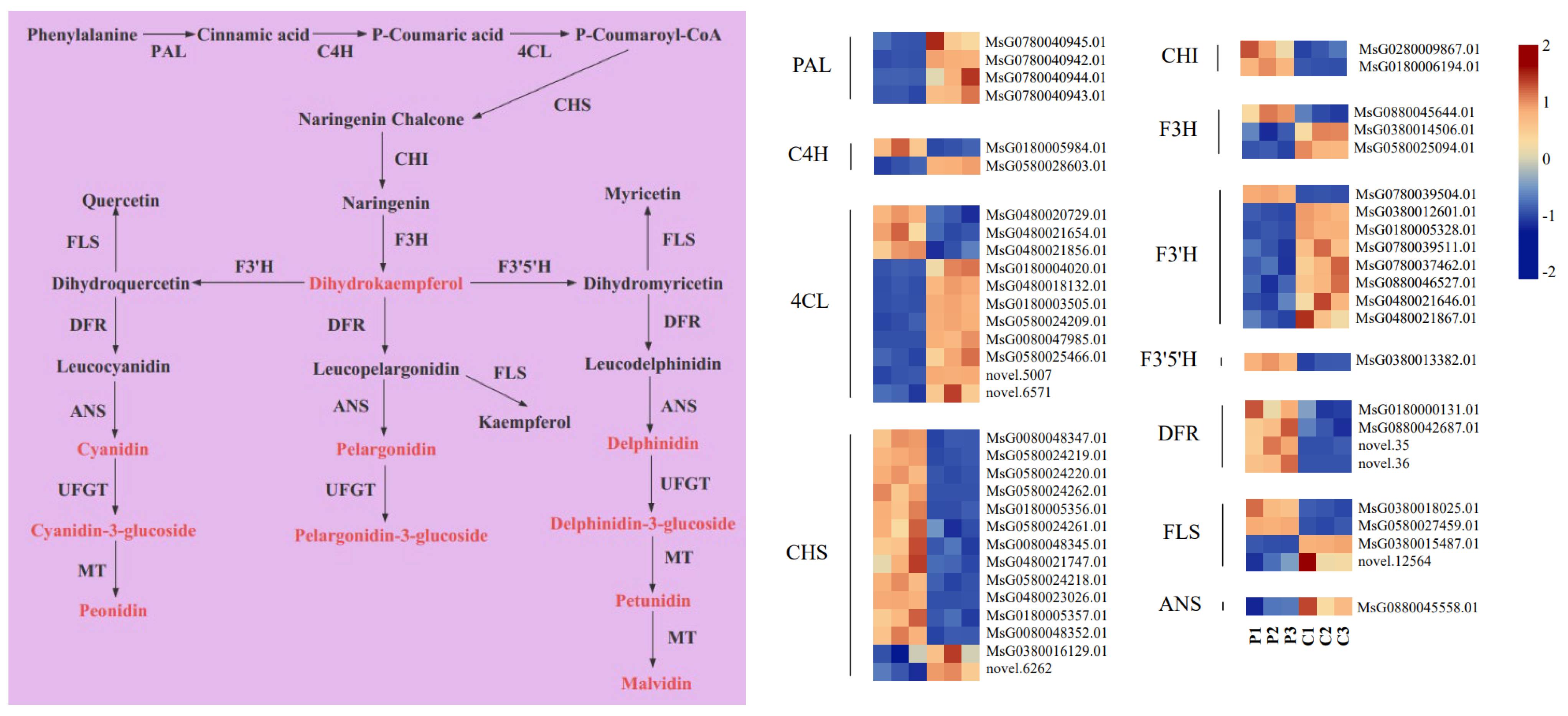
2.6. Expression Analysis of Anthocyanin Metabolic Pathway Genes
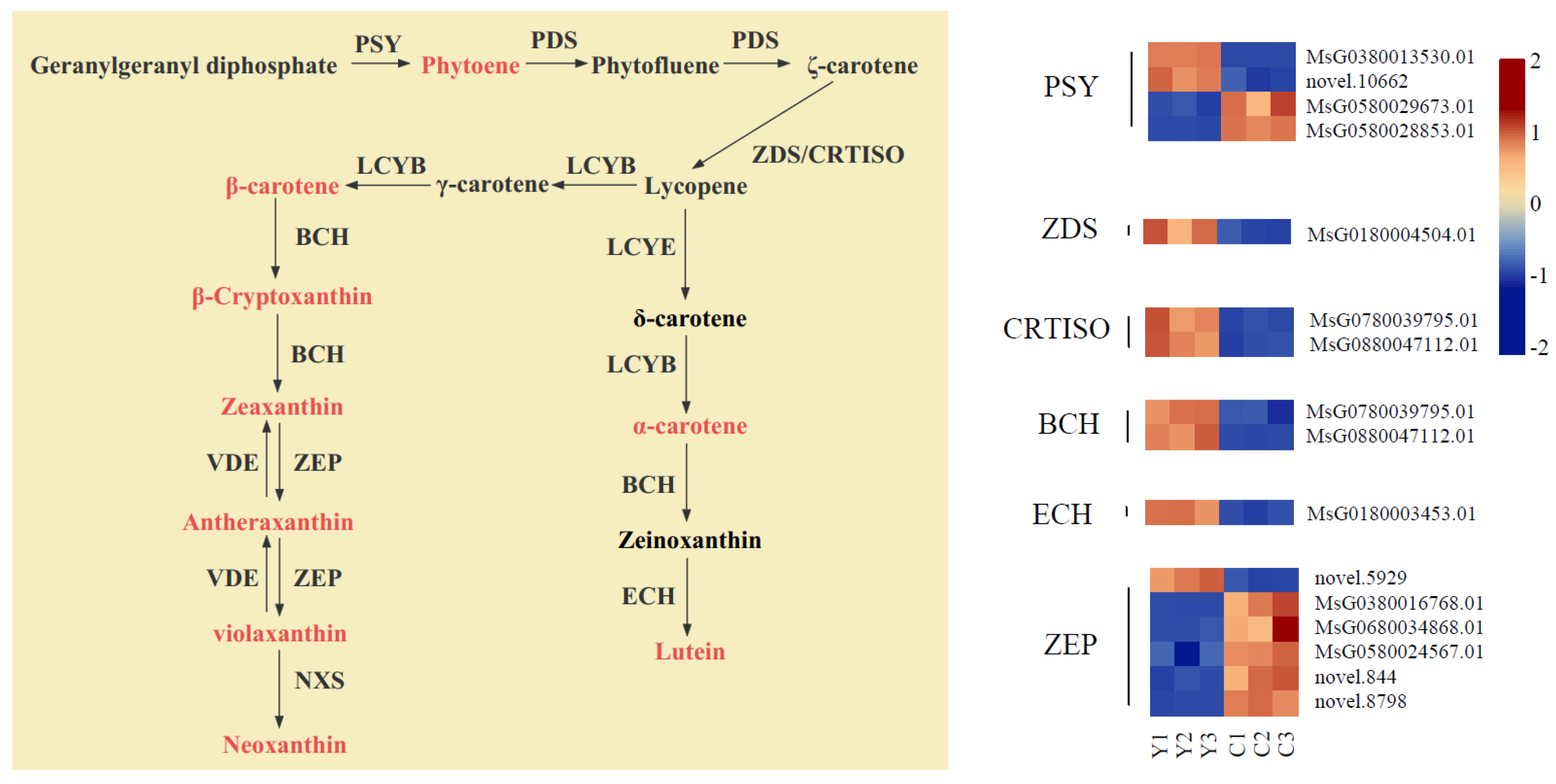
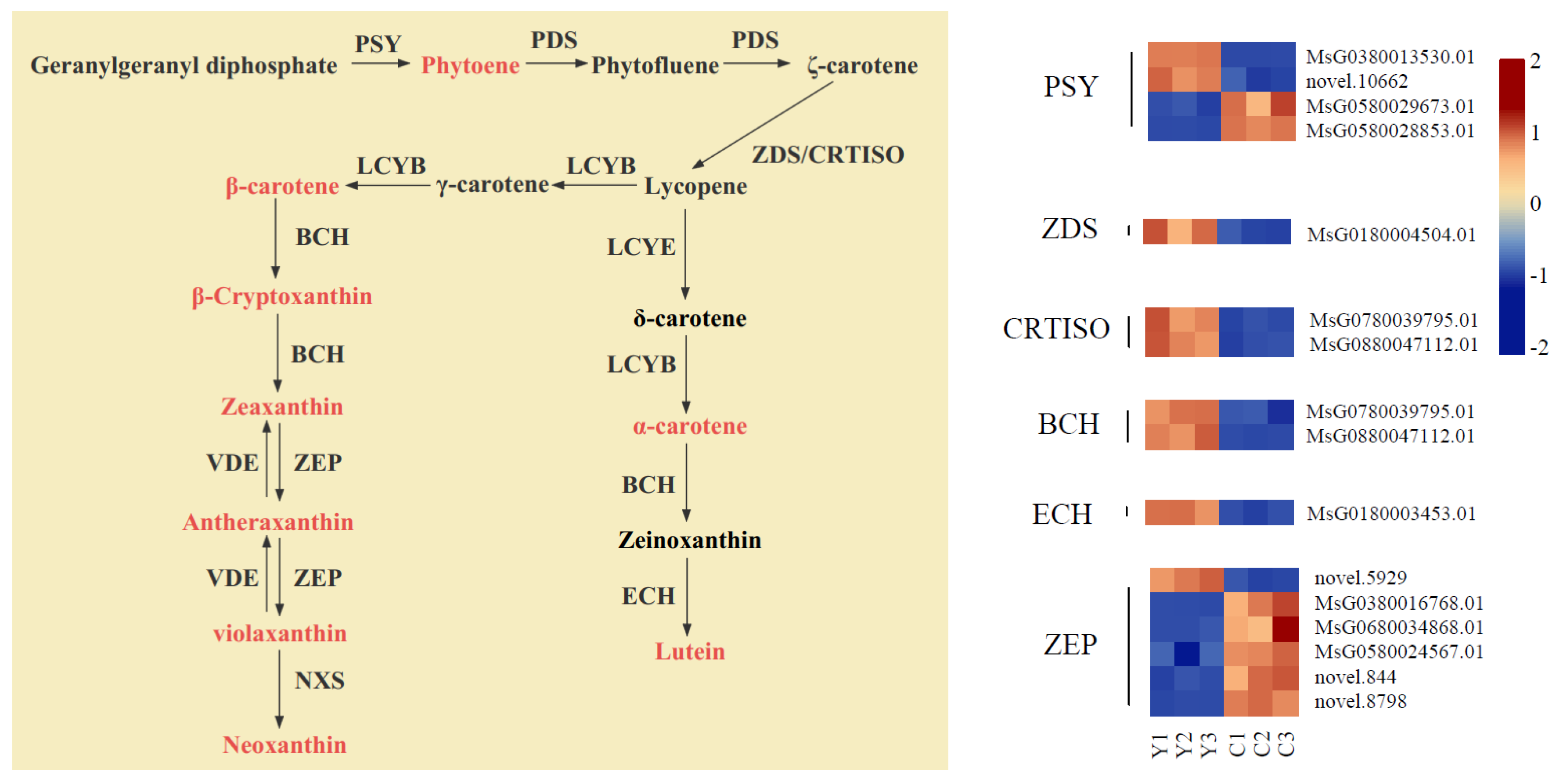
2.7. Expression Analysis of Carotenoid Metabolic Pathway Genes
2.8. Identification of Transcription Factors Related to Anthocyanin and Carotenoid Biosynthesis
3. Discussion
3.1. Component and Content of Anthocyanins and Carotenoids in Petals of the Three Different Colors
3.2. Key Genes That Participate in Anthocyanin Biosynthesis in Purple Flowers
3.3. Key Genes That Participate in Carotenoid Biosynthesis in Yellow Flowers
3.4. TFs Related to Anthocyanin and Carotenoid Biosynthesis
4. Materials and Methods
4.1. Plant Materials
4.2. Transcriptome Sequencing and Data Analysis
4.3. Extraction, Separation, Identification and Quantification of Anthocyanins
4.4. Extraction, Separation, Identification and Quantification of Carotenoids
4.5. qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagl, N.; Taski-Ajdukovic, K.; Barac, G.; Baburski, A.; Seccareccia, I.; Milic, D.; Katic, S. Estimation of the Genetic Diversity in Tetraploid Alfalfa Populations Based on RAPD Markers for Breeding Purposes. IJMS 2011, 12, 5449–5460. [Google Scholar] [CrossRef]
- Atumo, T.T.; Kauffman, R.; Gemiyo Talore, D.; Abera, M.; Tesfaye, T.; Tunkala, B.Z.; Zeleke, M.; Kebede Kalsa, G. Adaptability, Forage Yield and Nutritional Quality of Alfalfa (Medicago sativa) Genotypes. Sustain. Environ. 2021, 7, 1895475. [Google Scholar] [CrossRef]
- Miao, Z.; Xu, W.; Li, D.; Hu, X.; Liu, J.; Zhang, R.; Tong, Z.; Dong, J.; Su, Z.; Zhang, L.; et al. De Novo Transcriptome Analysis of Medicago Falcata Reveals Novel Insights about the Mechanisms Underlying Abiotic Stress-Responsive Pathway. BMC Genom. 2015, 16, 818. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lei, L.; Wengui, S.; Hongyan, L.; Fan, H.; Jun, L.; Meizhen, W. Study on the Taxonomy Status of Alfalfa with Cream Flowers. LR 2016, 39, 860–867. [Google Scholar] [CrossRef]
- Julier, B.; Huyghe, C.; Ecalle, C. Within- and Among-Cultivar Genetic Variation in Alfalfa: Forage Quality, Morphology, and Yield. Crop Sci. 2000, 40, 365–369. [Google Scholar] [CrossRef]
- Whitney, H.M.; Kolle, M.; Andrew, P.; Chittka, L.; Steiner, U.; Glover, B.J. Floral Iridescence, Produced by Diffractive Optics, Acts As a Cue for Animal Pollinators. Science 2009, 323, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, W.; Zhang, C.; Zhou, W.; Yan, P.; Xue, X.; Tian, Q.; Wang, D.; Niu, J.; Wang, S.; et al. Integrated Transcriptomic and Metabolomic Profiles Reveal Anthocyanin Accumulation in Scutellaria Baicalensis Petal Coloration. Ind. Crops Prod. 2023, 194, 116144. [Google Scholar] [CrossRef]
- Wang, F.; Ji, G.; Xu, Z.; Feng, B.; Zhou, Q.; Fan, X.; Wang, T. Metabolomics and Transcriptomics Provide Insights into Anthocyanin Biosynthesis in the Developing Grains of Purple Wheat (Triticum aestivum L.). J. Agric. Food Chem. 2021, 69, 11171–11184. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, L.; Sun, X.; Li, F.; Zhang, S.; Zhang, H.; Li, G.; Fang, Z.; Sun, R.; Hou, X.; et al. Transcriptome Analysis Reveals Anthocyanin Regulation in Chinese Cabbage (Brassica rapa L.) at Low Temperatures. Sci. Rep. 2022, 12, 6308. [Google Scholar] [CrossRef]
- Yang, L.; Ling, W.; Du, Z.; Chen, Y.; Li, D.; Deng, S.; Liu, Z.; Yang, L. Effects of Anthocyanins on Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2017, 8, 684–693. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Shao, S.; Sun, S.; Yang, D.; Lv, S. Anthocyanin: A Review of Plant Sources, Extraction, Stability, Content Determination and Modifications. Int. J. Food Sci. Tech. 2022, 57, 7573–7591. [Google Scholar] [CrossRef]
- Ferrer, J.-L.; Austin, M.B.; Stewart, C.; Noel, J.P. Structure and Function of Enzymes Involved in the Biosynthesis of Phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef]
- Qiu, W.; Su, W.; Cai, Z.; Dong, L.; Li, C.; Xin, M.; Fang, W.; Liu, Y.; Wang, X.; Huang, Z.; et al. Combined Analysis of Transcriptome and Metabolome Reveals the Potential Mechanism of Coloration and Fruit Quality in Yellow and Purple Passiflora edulis Sims. J. Agric. Food Chem. 2020, 68, 12096–12106. [Google Scholar] [CrossRef]
- He, X.; Zhao, X.; Gao, L.; Shi, X.; Dai, X.; Liu, Y.; Xia, T.; Wang, Y. Isolation and Characterization of Key Genes That Promote Flavonoid Accumulation in Purple-Leaf Tea (Camellia sinensis L.). Sci. Rep. 2018, 8, 130. [Google Scholar] [CrossRef]
- Chen, X.; Li, M.; Ni, J.; Hou, J.; Shu, X.; Zhao, W.; Su, P.; Wang, D.; Shah, F.A.; Huang, S.; et al. The R2R3-MYB Transcription Factor SsMYB1 Positively Regulates Anthocyanin Biosynthesis and Determines Leaf Color in Chinese Tallow (Sapium Sebiferum Roxb.). Ind. Crops Prod. 2021, 164, 113335. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A Colorful Model for the Regulation and Evolution of Biochemical Pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Gantt, E. A Study in Scarlet: Enzymes of Ketocarotenoid Biosynthesis in the Flowers of Adonis aestivalis. Plant J. 2005, 41, 478–492. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in Human Nutrition and Health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, Chemical Fingerprints and Distribution among Organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid Metabolism and Regulation in Horticultural Crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef]
- Stanley, L.; Yuan, Y.-W. Transcriptional Regulation of Carotenoid Biosynthesis in Plants: So Many Regulators, So Little Consensus. Front. Plant Sci. 2019, 10, 1017. [Google Scholar] [CrossRef]
- Ma, S.; Hu, R.; Ma, J.; Fan, J.; Wu, F.; Wang, Y.; Huang, L.; Feng, G.; Li, D.; Nie, G.; et al. Integrative Analysis of the Metabolome and Transcriptome Provides Insights into the Mechanisms of Anthocyanins and Proanthocyanidins Biosynthesis in Trifolium Repens. Ind. Crops Prod. 2022, 187, 115529. [Google Scholar] [CrossRef]
- Kumimoto, R.W.; Zhang, Y.; Siefers, N.; Holt, B.F. NF-YC3, NF-YC4 and NF-YC9 Are Required for CONSTANS-Mediated, Photoperiod-Dependent Flowering in Arabidopsis Thaliana: NF-YC Function in Flowering Time. Plant J. 2010, 63, 379–391. [Google Scholar] [CrossRef]
- Zheng, P.-F.; Yang, Y.-Y.; Zhang, S.; You, C.-X.; Zhang, Z.-L.; Hao, Y.-J. Identification and Functional Characterization of MdPIF3 in Response to Cold and Drought Stress in Malus domestica. Plant Cell Tissue Organ Cult. 2021, 144, 435–447. [Google Scholar] [CrossRef]
- Suttipanta, N.; Pattanaik, S.; Kulshrestha, M.; Patra, B.; Singh, S.K.; Yuan, L. The Transcription Factor CrWRKY1 Positively Regulates the Terpenoid Indole Alkaloid Biosynthesis in Catharanthus roseus. Plant Physiol. 2011, 157, 2081–2093. [Google Scholar] [CrossRef] [PubMed]
- Flajoulot, S.; Ronfort, J.; Baudouin, P.; Barre, P.; Huguet, T.; Huyghe, C.; Julier, B. Genetic Diversity among Alfalfa (Medicago sativa) Cultivars Coming from a Breeding Program, Using SSR Markers. Theor. Appl. Genet. 2005, 111, 1420–1429. [Google Scholar] [CrossRef]
- Clifford, M.N. Anthocyanins—Nature, Occurrence and Dietary Burden. J. Sci. Food Agric. 2000, 80, 1063–1072. [Google Scholar] [CrossRef]
- Duan, H.-R.; Wang, L.-R.; Cui, G.-X.; Zhou, X.-H.; Duan, X.-R.; Yang, H.-S. Identification of the Regulatory Networks and Hub Genes Controlling Alfalfa Floral Pigmentation Variation Using RNA-Sequencing Analysis. BMC Plant Biol. 2020, 20, 110. [Google Scholar] [CrossRef]
- Schweiggert, R.M.; Steingass, C.B.; Heller, A.; Esquivel, P.; Carle, R. Characterization of Chromoplasts and Carotenoids of Red- and Yellow-Fleshed Papaya (Carica papaya L.). Planta 2011, 234, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Iijima, L.; Kishimoto, S.; Ohmiya, A.; Yagi, M.; Okamoto, E.; Miyahara, T.; Tsujimoto, T.; Ozeki, Y.; Uchiyama, N.; Hakamatsuka, T.; et al. Esterified Carotenoids Are Synthesized in Petals of Carnation (Dianthus caryophyllus) and Accumulate in Differentiated Chromoplasts. Sci. Rep. 2020, 10, 15256. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wan, R.; Shi, Z.; Ma, W.; Wang, H.; Chen, Y.; Bo, J.; Li, Y.; An, W.; Qin, K.; et al. Transcriptomics and Metabolomics Reveal the Critical Genes of Carotenoid Biosynthesis and Color Formation of Goji (Lycium barbarum L.) Fruit Ripening. Plants 2023, 12, 2791. [Google Scholar] [CrossRef]
- Nakamura, N.; Fukuchi-Mizutani, M.; Fukui, Y.; Ishiguro, K.; Suzuki, K.; Suzuki, H.; Okazaki, K.; Shibata, D.; Tanaka, Y. Generation of Pink Flower Varieties from Blue Torenia Hybrida by Redirecting the Flavonoid Biosynthetic Pathway from Delphinidin to Pelargonidin. Plant Biotechnol. 2010, 27, 375–383. [Google Scholar] [CrossRef]
- Katsumoto, Y.; Fukuchi-Mizutani, M.; Fukui, Y.; Brugliera, F.; Holton, T.A.; Karan, M.; Nakamura, N.; Yonekura-Sakakibara, K.; Togami, J.; Pigeaire, A.; et al. Engineering of the Rose Flavonoid Biosynthetic Pathway Successfully Generated Blue-Hued Flowers Accumulating Delphinidin. Plant Cell Physiol. 2007, 48, 1589–1600. [Google Scholar] [CrossRef]
- Deng, X.; Bashandy, H.; Ainasoja, M.; Kontturi, J.; Pietiäinen, M.; Laitinen, R.A.E.; Albert, V.A.; Valkonen, J.P.T.; Elomaa, P.; Teeri, T.H. Functional Diversification of Duplicated Chalcone Synthase Genes in Anthocyanin Biosynthesis of Gerbera hybrida. New Phytol. 2014, 201, 1469–1483. [Google Scholar] [CrossRef]
- Hong, S.; Wang, J.; Wang, Q.; Zhang, G.; Zhao, Y.; Ma, Q.; Wu, Z.; Ma, J.; Gu, C. Decoding the Formation of Diverse Petal Colors of Lagerstroemia Indica by Integrating the Data from Transcriptome and Metabolome. Front. Plant Sci. 2022, 13, 970023. [Google Scholar] [CrossRef]
- Okinaka, Y.; Shimada, Y.; Nakano-Shimada, R.; Ohbayashi, M.; Kiyokawa, S.; Kikuchi, Y. Selective Accumulation of Delphinidin Derivatives in Tobacco Using a Putative Flavonoid 3′,5′-Hydroxylase cDNA from Campanula medium. Biosci. Biotechnol. Biochem. 2003, 67, 161–165. [Google Scholar] [CrossRef]
- Tsuda, S.; Fukui, Y.; Nakamura, N.; Katsumoto, Y.; Yonekura-Sakakibara, K.; Fukuchi-Mizutani, M.; Ohira, K.; Ueyama, Y.; Ohkawa, H.; Holton, T.A.; et al. Flower Color Modification of Petunia Hybrida Commercial Varieties by Metabolic Engineering. Plant Biotechnol. 2004, 21, 377–386. [Google Scholar] [CrossRef]
- Zhou, X.-W.; Fan, Z.-Q.; Chen, Y.; Zhu, Y.-L.; Li, J.-Y.; Yin, H.-F. Functional Analyses of a Flavonol Synthase–like Gene from Camellia Nitidissima Reveal Its Roles in Flavonoid Metabolism during Floral Pigmentation. J. Biosci. 2013, 38, 593–604. [Google Scholar] [CrossRef]
- Holton, T.A.; Brugliera, F.; Tanaka, Y. Cloning and Expression of Flavonol Synthase from Petunia hybrida. Plant J. 1993, 4, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Cazzonelli, C.I.; Pogson, B.J. Source to Sink: Regulation of Carotenoid Biosynthesis in Plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Pollack, A.J.; Gong, X.; Pollack, J.R. A Common Phytoene Synthase Mutation Underlies White Petal Varieties of the California Poppy. Sci. Rep. 2019, 9, 11615. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, S.; Lu, Z.; He, Z.; Ye, Y.; Zhao, B.; Wang, L.; Jin, B. Cytological, Physiological, and Transcriptomic Analyses of Golden Leaf Coloration in Ginkgo biloba L. Hortic. Res. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-M.; To, K.-Y.; Lai, H.-M.; Jeng, S.-T. Modification of Flower Colour by Suppressing β-ring Carotene Hydroxylase Genes in Oncidium. Plant Biol. J. 2016, 18, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Schwinn, K.; Venail, J.; Shang, Y.; Mackay, S.; Alm, V.; Butelli, E.; Oyama, R.; Bailey, P.; Davies, K.; Martin, C. A Small Family of MYB -Regulatory Genes Controls Floral Pigmentation Intensity and Patterning in the Genus Antirrhinum. Plant Cell 2006, 18, 831–851. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, S.; Wang, Y.; Wang, Y.; Song, A.; Jiang, J.; Chen, S.; Guan, Z.; Chen, F. CmMYB3-like Negatively Regulates Anthocyanin Biosynthesis and Flower Color Formation during the Post-Flowering Stage in Chrysanthemum Morifolium. Hortic. Plant J. 2023, 10, 194–204. [Google Scholar] [CrossRef]
- Wang, L.; Tang, W.; Hu, Y.; Zhang, Y.; Sun, J.; Guo, X.; Lu, H.; Yang, Y.; Fang, C.; Niu, X.; et al. AMYB/bHLH Complex Regulates Tissue-specific Anthocyanin Biosynthesis in the Inner Pericarp of Red-centered Kiwifruit Actinidia chinensis Cv. Hongyang. Plant J. 2019, 99, 359–378. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Y.; Yu, L.; Jiang, H.; Guo, Z.; Xu, H.; Jiang, S.; Fang, H.; Zhang, J.; Su, M.; et al. MdWRKY11 Participates in Anthocyanin Accumulation in Red-Fleshed Apples by Affecting MYB Transcription Factors and the Photoresponse Factor MdHY5. J. Agric. Food Chem. 2019, 67, 8783–8793. [Google Scholar] [CrossRef]
- Welsch, R.; Maass, D.; Voegel, T.; DellaPenna, D.; Beyer, P. Transcription Factor RAP2.2 and Its Interacting Partner SINAT2: Stable Elements in the Carotenogenesis of Arabidopsis Leaves. Plant Physiol. 2007, 145, 1073–1085. [Google Scholar] [CrossRef]
- Zhu, F.; Luo, T.; Liu, C.; Wang, Y.; Yang, H.; Yang, W.; Zheng, L.; Xiao, X.; Zhang, M.; Xu, R.; et al. An R2R3-MYB Transcription Factor Represses the Transformation of A- and Β-branch Carotenoids by Negatively Regulating Expression of CrBCH2 and CrNCED5 in Flavedo of Citrus reticulate. New Phytol. 2017, 216, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Ortiz, G.; Johansson, H.; Lee, K.P.; Bou-Torrent, J.; Stewart, K.; Steel, G.; Rodríguez-Concepción, M.; Halliday, K.J. The HY5-PIF Regulatory Module Coordinates Light and Temperature Control of Photosynthetic Gene Transcription. PLoS Genet. 2014, 10, e1004416. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wu, M.; Cao, L.; Yuan, W.; Dong, M.; Wang, X.; Chen, W.; Shang, F. Characterization of OfWRKY3, a Transcription Factor That Positively Regulates the Carotenoid Cleavage Dioxygenase Gene OfCCD4 in Osmanthus Fragrans. Plant Mol. Biol. 2016, 91, 485–496. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of DAMs | Upregulated | Downregulated |
|---|---|---|---|
| P vs. C | 42 | 33 | 9 |
| Y vs. C | 35 | 15 | 20 |
| P vs. Y | 51 | 38 | 13 |
| Group | Number of DAMs | Upregulated | Downregulated |
|---|---|---|---|
| P vs. C | 20 | 11 | 9 |
| Y vs. C | 38 | 33 | 5 |
| P vs. Y | 39 | 32 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Liu, L.; Qiang, X.; Meng, Y.; Li, Z.; Huang, F. Integrated Metabolomic and Transcriptomic Profiles Provide Insights into the Mechanisms of Anthocyanin and Carotenoid Biosynthesis in Petals of Medicago sativa ssp. sativa and Medicago sativa ssp. falcata. Plants 2024, 13, 700. https://doi.org/10.3390/plants13050700
Huang X, Liu L, Qiang X, Meng Y, Li Z, Huang F. Integrated Metabolomic and Transcriptomic Profiles Provide Insights into the Mechanisms of Anthocyanin and Carotenoid Biosynthesis in Petals of Medicago sativa ssp. sativa and Medicago sativa ssp. falcata. Plants. 2024; 13(5):700. https://doi.org/10.3390/plants13050700
Chicago/Turabian StyleHuang, Xiuzheng, Lei Liu, Xiaojing Qiang, Yuanfa Meng, Zhiyong Li, and Fan Huang. 2024. "Integrated Metabolomic and Transcriptomic Profiles Provide Insights into the Mechanisms of Anthocyanin and Carotenoid Biosynthesis in Petals of Medicago sativa ssp. sativa and Medicago sativa ssp. falcata" Plants 13, no. 5: 700. https://doi.org/10.3390/plants13050700
APA StyleHuang, X., Liu, L., Qiang, X., Meng, Y., Li, Z., & Huang, F. (2024). Integrated Metabolomic and Transcriptomic Profiles Provide Insights into the Mechanisms of Anthocyanin and Carotenoid Biosynthesis in Petals of Medicago sativa ssp. sativa and Medicago sativa ssp. falcata. Plants, 13(5), 700. https://doi.org/10.3390/plants13050700