
Cabbage (Brassica oleracea var. capitata) Development in Time: How Differential Parenchyma Tissue Growth Affects Leafy Head Formation



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Cabbage Plants and Leaf Development in Planta and Histological Studies
2.3. Leaf Morphology Studies
3. Results
3.1. Whole Plant Development
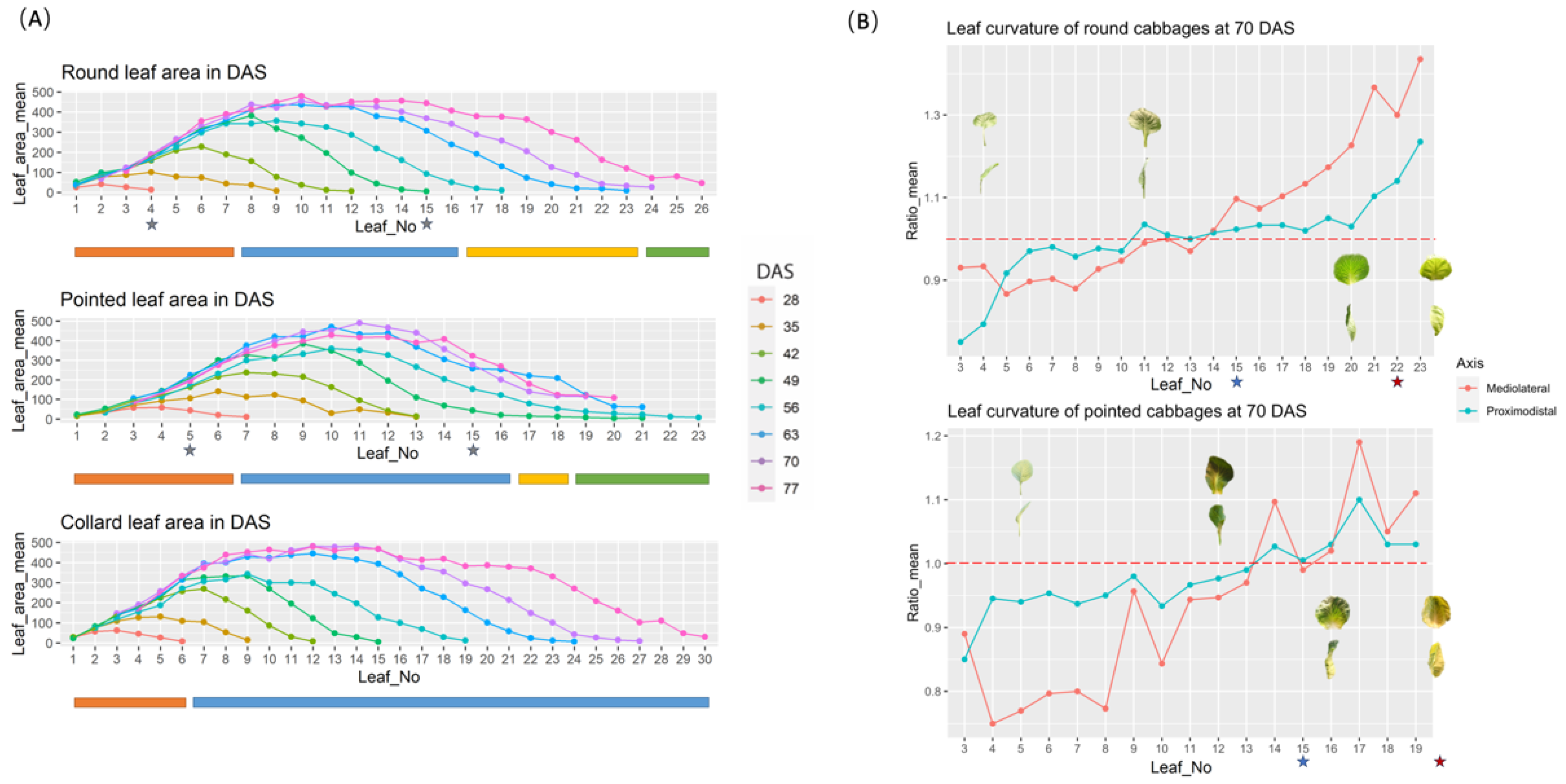
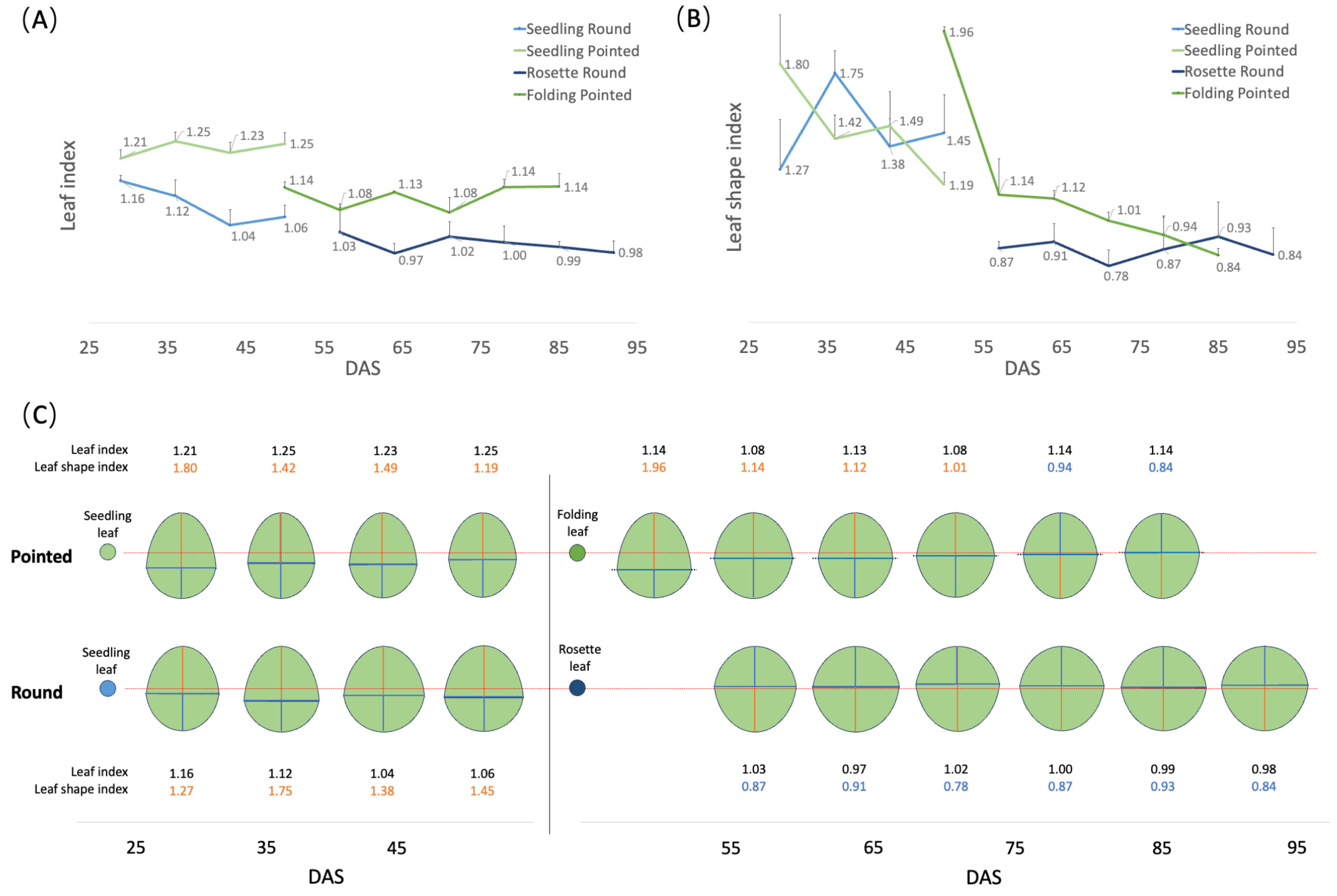
3.2. Changes in Leaf Morphology during Plant Development
3.3. Defining the Developmental Stages of Cabbage
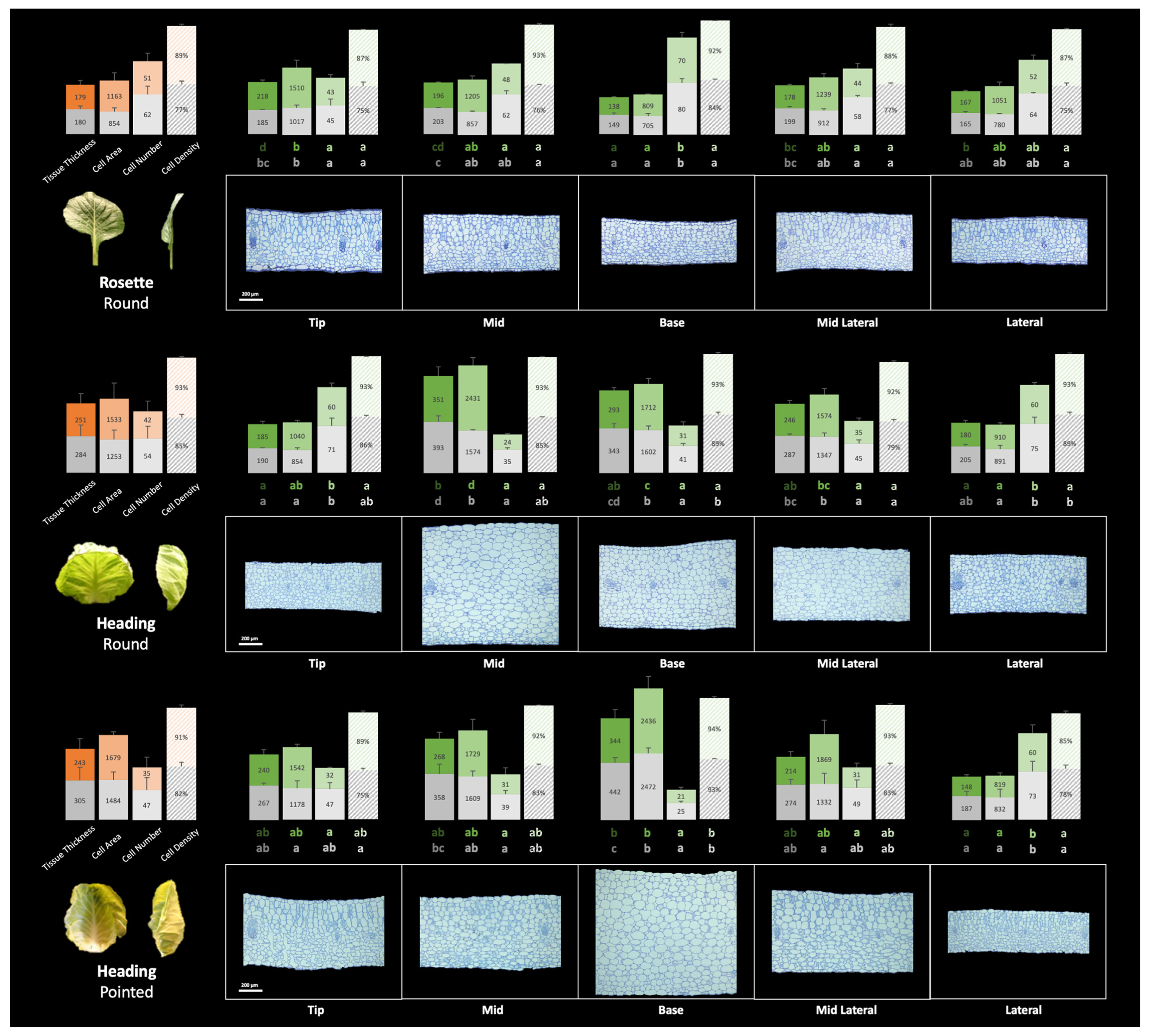
3.4. Histology of Leaf Development
3.4.1. Difference among Leaf Positions
3.4.2. Difference between Adaxial and Abaxial Sides
Tissue Thickness
Average Single Cell Area and Cell Density
3.4.3. Shoot Apical Meristem
4. Discussion
4.1. Characterization and Timing of Developmental Stages
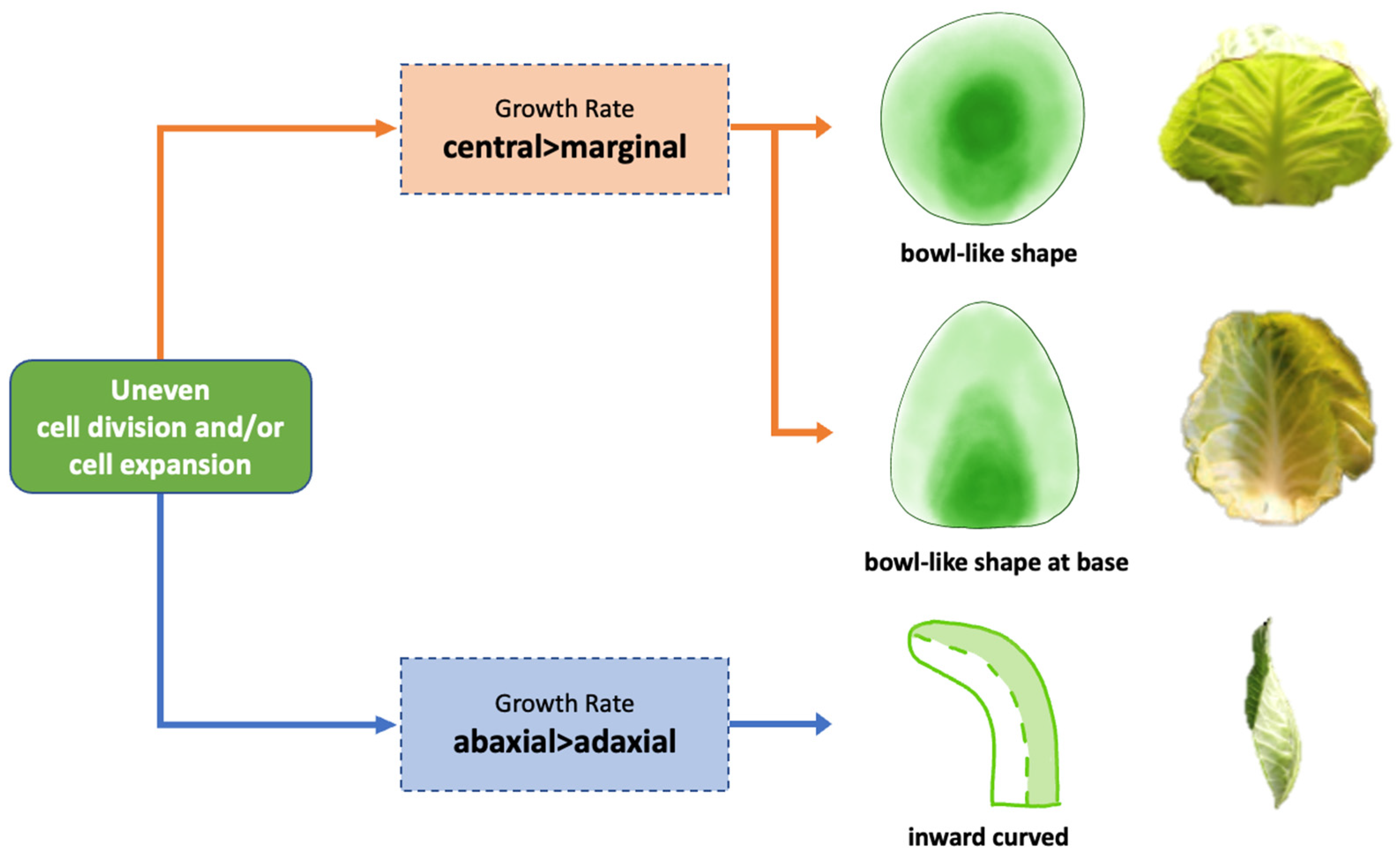
4.2. Differential Growth across the Leaf Blade and between the Mesophyll Cell Layers Is Associated with Leaf Curvature
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nawaz, H.; Shad, M.A.; Rauf, A. Optimization of extraction yield and antioxidant properties of Brassica oleracea Convar Capitata Var L. leaf extracts. Food Chem. 2018, 242, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liang, J.; Lin, R.; Zhang, L.; Wu, J.; Wang, X. Series-Spatial Transcriptome Profiling of Leafy Head Reveals the Key Transition Leaves for Head Formation in Chinese Cabbage. Front. Plant Sci. 2021, 12, 787826. [Google Scholar] [CrossRef] [PubMed]
- He, Y.K.; Xue, W.X.; Sun, Y.D.; Yu, X.H.; Liu, P.L. Leafy head formation of the progenies of transgenic plants of Chinese cabbage with exogenous auxin genes. Cell Res. 2000, 10, 151–160. [Google Scholar] [CrossRef]
- Sun, X.; Basnet, R.K.; Yan, Z.; Bucher, J.; Cai, C.; Zhao, J.; Bonnema, G. Genome-wide transcriptome analysis reveals molecular pathways involved in leafy head formation of Chinese cabbage (Brassica rapa). Hortic. Res. 2019, 6, 130. [Google Scholar] [CrossRef]
- Mao, Y.; Wu, F.; Yu, X.; Bai, J.; Zhong, W.; He, Y. MicroRNA319a-targeted Brassica rapa ssp. pekinensis TCP genes modulate head shape in chinese cabbage by differential cell division arrest in leaf regions. Plant Physiol. 2014, 164, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, F.; Bai, J.; He, Y. BrpSPL9 (Brassica rapa ssp. pekinensis SPL9) controls the earliness of heading time in Chinese cabbage. Plant Biotechnol. J. 2014, 12, 312–321. [Google Scholar] [CrossRef]
- Alemán-Báez, J.; Qin, J.; Cai, C.; Zou, C.; Bucher, J.; Paulo, M.-J.; Voorrips, R.E.; Bonnema, G. Genetic dissection of morphological variation in rosette leaves and leafy heads in cabbage (Brassica oleracea var. capitata). Theor. Appl. Genet. 2022, 135, 3611–3628. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, Y.; Wu, J.; Liang, J.; Chen, S.; Zhang, L.; Lv, H.; Yin, X.; Zhang, X.; Zhang, Y.; et al. A cluster of transcripts identifies a transition stage initiating leafy head growth in heading morphotypes of Brassica. Plant J. 2022, 110, 688–706. [Google Scholar] [CrossRef]
- Yue, X.; Su, T.; Xin, X.; Li, P.; Wang, W.; Yu, Y.; Zhang, D.; Zhao, X.; Wang, J.; Sun, L.; et al. The Adaxial/Abaxial Patterning of Auxin and Auxin Gene in Leaf Veins Functions in Leafy Head Formation of Chinese Cabbage. Front. Plant Sci. 2022, 13, 918112. [Google Scholar] [CrossRef]
- Oguchi, R.; Onoda, Y.; Terashima, I.; Tholen, D. Leaf anatomy and function. In The Leaf: A Platform for Performing Photosynthesis; Springer: Cham, Switzerland, 2018; pp. 97–139. [Google Scholar]
- Nath, U.; Crawford, B.C.; Carpenter, R.; Coen, E. Genetic control of surface curvature. Science 2003, 299, 1404–1407. [Google Scholar] [CrossRef]
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef]
- Liang, J.; Liu, B.; Wu, J.; Cheng, F.; Wang, X. Genetic variation and divergence of genes involved in leaf adaxial-abaxial polarity establishment in Brassica rapa. Front. Plant Sci. 2016, 7, 94. [Google Scholar] [CrossRef]
- Moon, J.; Hake, S. How a leaf gets its shape. Curr. Opin. Plant Biol. 2011, 14, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Wu, F.; Bai, J.; Li, X.; Yang, X.; Xue, W.; Liu, H.; He, Y. BcpLH organizes a specific subset of microRNAs to form a leafy head in Chinese cabbage (Brassica rapa ssp. pekinensis). Hortic. Res. 2020, 7, 1. [Google Scholar] [CrossRef]
- Han, M.-H.; Goud, S.; Song, L.; Fedoroff, N. The Arabidopsis double-stranded RNA-binding protein HYL1 plays a role in microRNA-mediated gene regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Bucher, J.; Bakker, F.T.; Bonnema, G. Evidence for two domestication lineages supporting a middle-eastern origin for Brassica oleracea crops from diversified kale populations. Hortic. Res. 2022, 9, uhac033. [Google Scholar] [CrossRef]
- Juss, P.; Shaw, P. Protocol for Test on Distinctness, Uniformity and Stability; Community Plant Varienty Office (CPVO-TP): Angers, France, 2014; Volume 1, pp. 286–310. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Alemán-Báez, J.; Acevedo-Zamora, J.F.; Bucher, J.; Cai, C.; Voorrips, R.E.; Bonnema, G. Expression Changes of miRNA-Regulated Genes Associated with the Formation of the Leafy Head in Cabbage (Brassica oleracea var. capitata). Hortic. Plant J. 2024, accepted. [Google Scholar]
- Sun, X.; Luo, S.; Luo, L.; Wang, X.; Chen, X.; Lu, Y.; Shen, S.; Zhao, J.; Bonnema, G. Genetic analysis of Chinese cabbage reveals correlation between rosette leaf and leafy head variation. Front. Plant Sci. 2018, 9, 1455. [Google Scholar] [CrossRef]
- Yu, L.; Yu, X.; Shen, R.; He, Y. HYL1 gene maintains venation and polarity of leaves. Planta 2005, 221, 231–242. [Google Scholar] [CrossRef]
- Ren, W.; Wang, H.; Bai, J.; Wu, F.; He, Y. Association of microRNAs with types of leaf curvature in Brassica rapa. Front. Plant Sci. 2018, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Cai, C.; Ji, J.; Han, F.; Zhang, L.; Chen, S.; Zhang, L.; Yang, Y.; Tang, Q.; et al. Large-scale gene expression alterations introduced by structural variation drive morphotype diversification in Brassica oleracea. Nat. Genet. 2024, 1–13. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sowing Date | Plants in Total | Plants Utilized in the Experiment | Type of Experiment | Experiment | Cultivars | Material Scored | Scoring Timepoints (DAS) | Traits Scored |
|---|---|---|---|---|---|---|---|---|
| March | 18 (2 cabbage cultivars × 3 blocks × 3 plants) | 3 plants per cultivar (1 per block) | Non-destructive | Whole plant development | Round and pointed cabbage | Aerial part of cabbage plants | 30, 36, 43 51, 58, 65, 72, 78, 86, 91, 98, 106, 112, 120, and 126 | Plant growth |
| Leaf development in planta | 4th leaf for round cabbage, 5th leaf for pointed cabbage | 29, 36, 43, and 50 | Leaf blade area, leaf index, and leaf shape index | |||||
| 15th leaf for round and pointed cabbage | 50, 57, 64, 71, 78, 85, and 92 | |||||||
| 3 plants per cultivar (1 per block) | Histology of leaves | Round cabbage | Leaves 15th, 15th, 15th | t1 = 57 and t2 = 85 | Leaf tissue from 5 positions (tip, mid, base, mid-lateral, and lateral) | |||
| Leaves 22nd, 22nd, 24th | t1 = 98 and t2 = 126 | |||||||
| Pointed cabbage | Leaves 15th, 15th, 17th | t1 = 57 and t2 = 85 | ||||||
| Leaves 17th, 21st, 22nd | t1 = 71 and t2 = 99 | |||||||
| August | 90 (3 cultivars × 3 blocks × 10 plants) | 3 plants per timepoint per cultivar (1 per block) | Destructive | Leaf morphology and development | Round and pointed cabbage and collard | Leaves | 28, 35, 42, 49, 56, 63, 70, and 77 | Total number of leaves and their leaf area |
| Round and pointed cabbage | Leaf blade | 70 | Leaf curvature | |||||
| Leafy head | - | First heading leaf |
| Stages | Seedling | Rosette | Folding | Heading | |
|---|---|---|---|---|---|
| Round (Excalibur hybrid variety) | DAS | ~10 to ~38 | ~38 to ~60 | ~60 to ~70 (latest 84) | ~70 to cracking |
| Duration (days) | ~28 | ~22 | ~10 | >56 | |
| Leaf number | 1st to 7th | 8th to ~15th | ~16th to ~23rd (latest 24th) | ~24th to … | |
| Pointed (Sonsma hybrid variety) | DAS | ~8 to ~33 | ~33 to ~48 | ~48 to ~56 (latest 84) | ~56 to cracking |
| Duration (days) | ~25 | ~15 | ~8 | >56 | |
| Leaf number | 1st to 6th | 7th to ~14th | ~15th to ~17th (latest 21st) | ~18th to … |
 Rosette Round | P/S (t1) | P/S (t2) | ∆P/∆S | |
| Tissue Thickness | 1.00 | 1.00 | 1.00 | |
| Single Cell Area | 1.02 | 1.36 | 1.48 | |
| Cell Density | 1.02 | 1.15 | -0.24 | |
 Heading Round | P/S (t1) | P/S (t2) | ∆P/∆S | |
| Tissue Thickness | 0.88 | 0.86 | 0.79 | |
| Single Cell Area | 1.23 | 1.21 | 1.17 | |
| Cell Density | 1.05 | 1.08 | 0.46 | |
 Heading Pointed | P/S (t1) | P/S (t2) | ∆P/∆S | |
| Tissue Thickness | 0.79 | 0.79 | 0.74 | |
| Single Cell Area | 1.03 | 1.13 | 1.41 | |
| Cell Density | 1.06 | 1.10 | 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Alemán-Báez, J.; Visser, R.G.F.; Bonnema, G. Cabbage (Brassica oleracea var. capitata) Development in Time: How Differential Parenchyma Tissue Growth Affects Leafy Head Formation. Plants 2024, 13, 656. https://doi.org/10.3390/plants13050656
Liu Z, Alemán-Báez J, Visser RGF, Bonnema G. Cabbage (Brassica oleracea var. capitata) Development in Time: How Differential Parenchyma Tissue Growth Affects Leafy Head Formation. Plants. 2024; 13(5):656. https://doi.org/10.3390/plants13050656
Chicago/Turabian StyleLiu, Zihan, Jorge Alemán-Báez, Richard G. F. Visser, and Guusje Bonnema. 2024. "Cabbage (Brassica oleracea var. capitata) Development in Time: How Differential Parenchyma Tissue Growth Affects Leafy Head Formation" Plants 13, no. 5: 656. https://doi.org/10.3390/plants13050656
APA StyleLiu, Z., Alemán-Báez, J., Visser, R. G. F., & Bonnema, G. (2024). Cabbage (Brassica oleracea var. capitata) Development in Time: How Differential Parenchyma Tissue Growth Affects Leafy Head Formation. Plants, 13(5), 656. https://doi.org/10.3390/plants13050656