
Hormonal Regulation and Transcriptomic Insights into Flower Development in Hydrangea paniculata ‘Vanilla Strawberry’


 and
and
Abstract
1. Introduction
2. Results
2.1. Effect of Plant Growth Regulators on the Flowering Stage of Conical Hydrangea
2.2. Effects of Plant Growth Regulators on the Inflorescence of Hydrangea paniculata
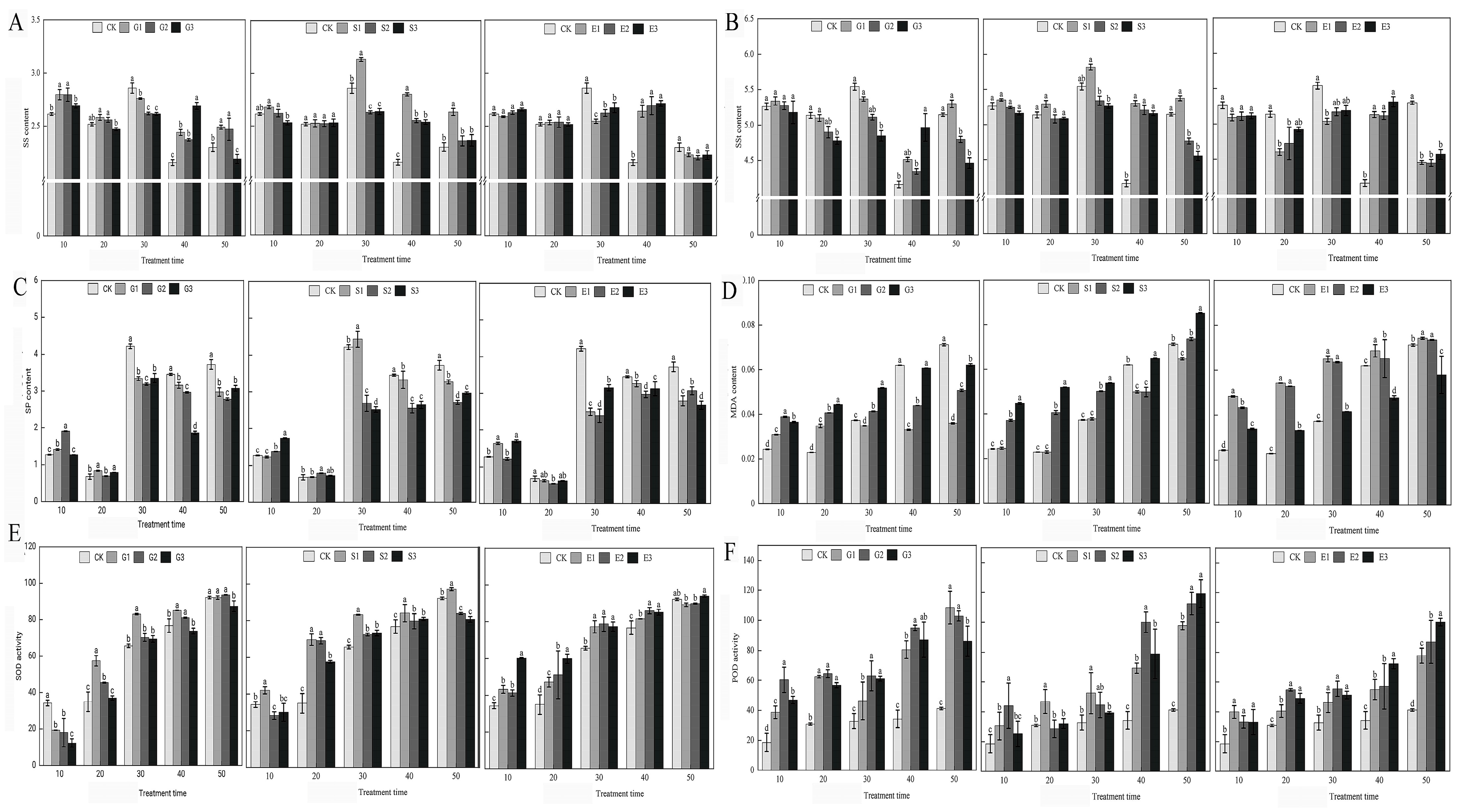
2.3. Physiological Responses to Plant Growth Regulators in Hydrangea paniculata
2.4. Library Construction and Transcriptome Sequencing
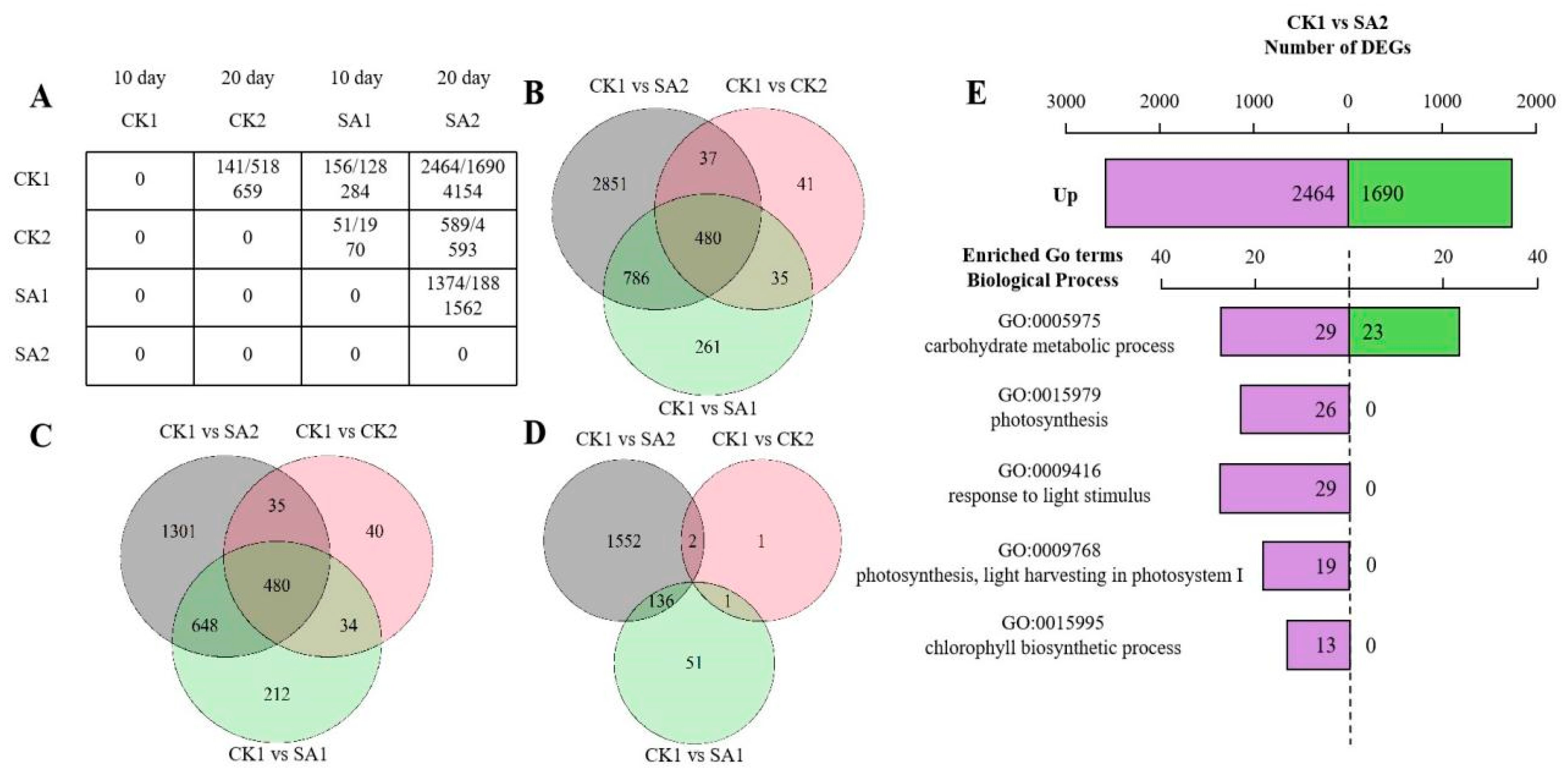
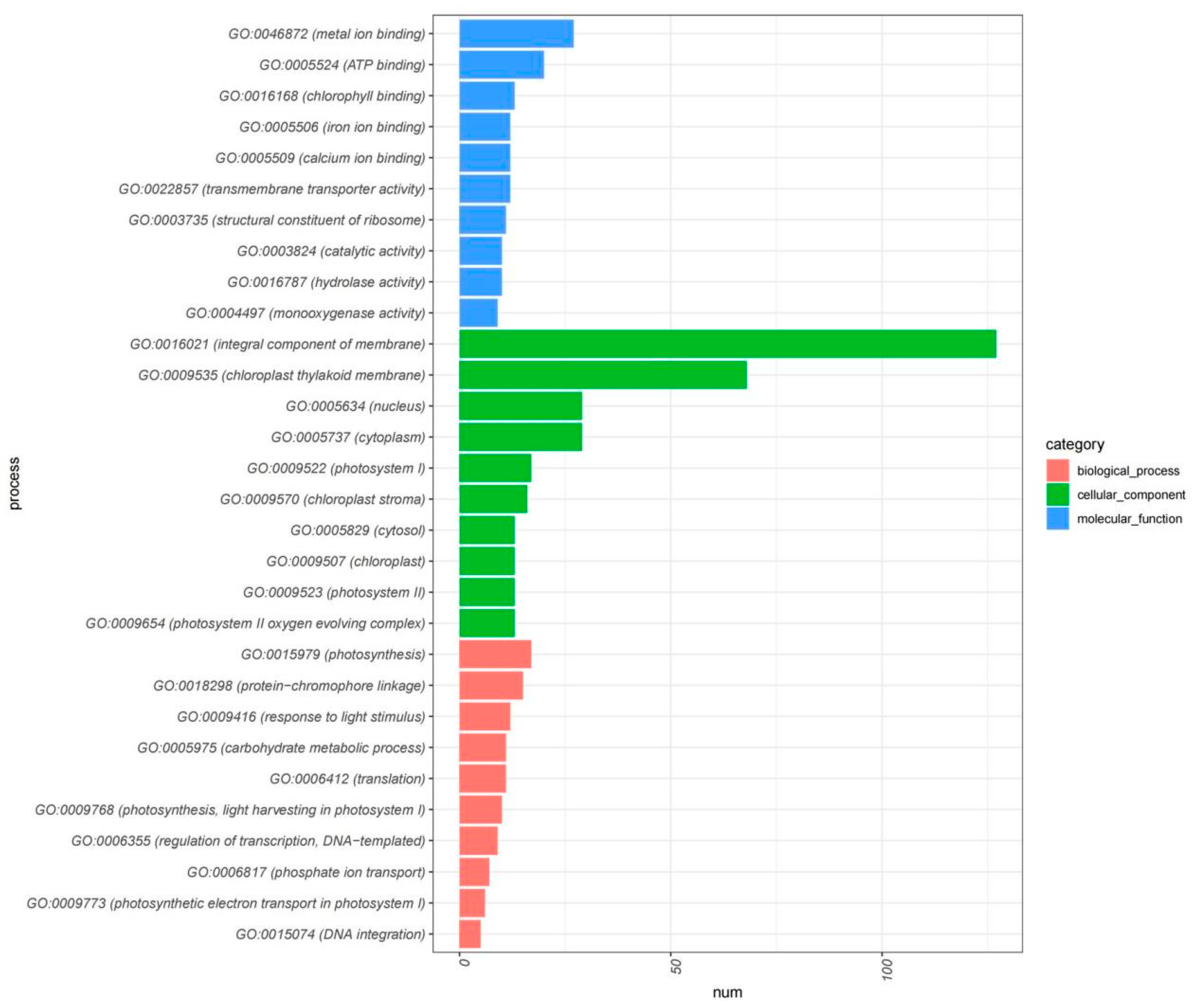
2.5. Transcriptomic Profiling Reveals Differential Gene Expression Dynamics in Flower Development and Opening
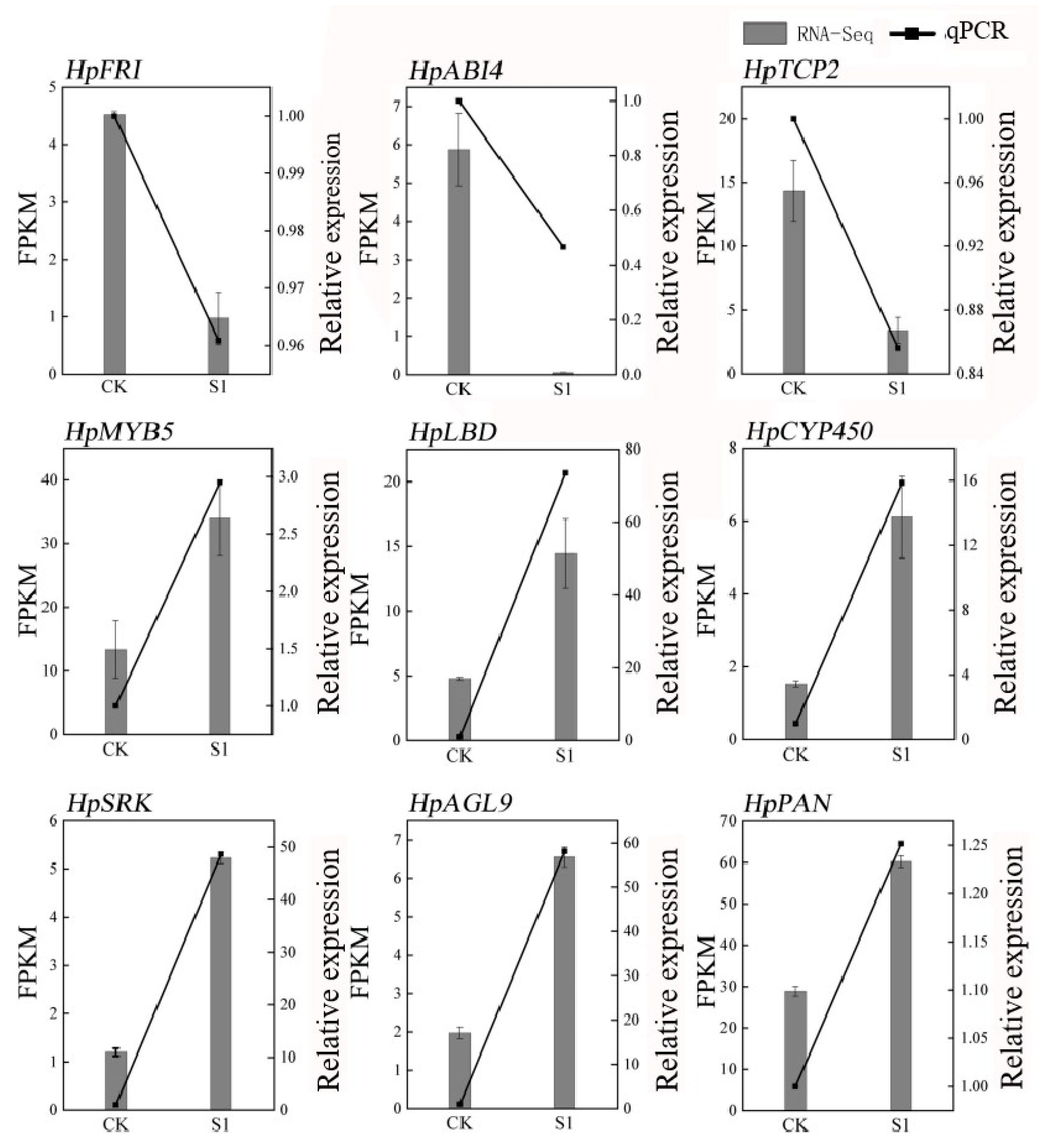
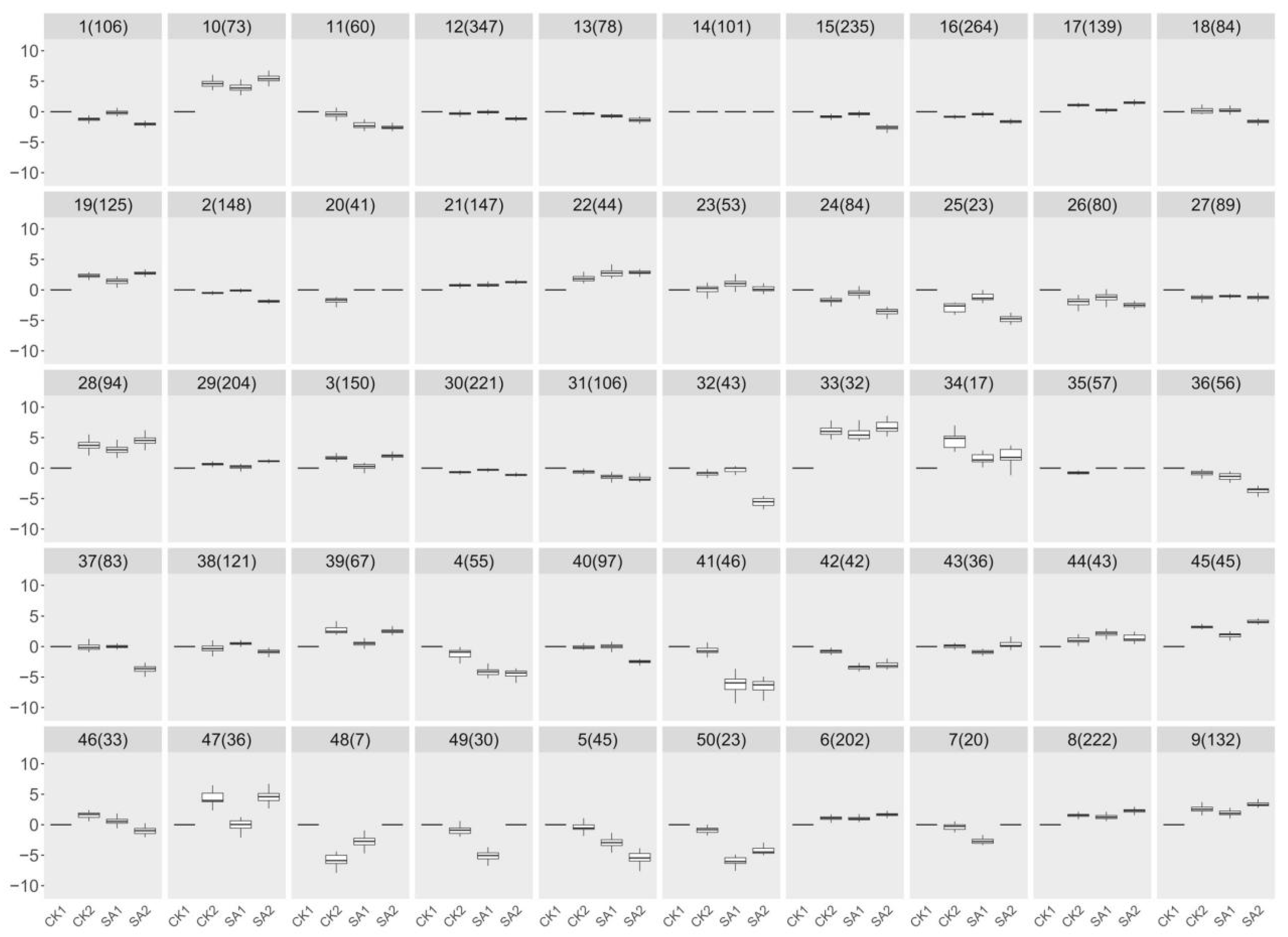
2.6. Elucidating Salicylic Acid-Induced Gene Expression Patterns in Flower Development: A Hierarchical Clustering Approach
2.7. Regulatory Dynamics of Myb Transcription Factors in Plant Development and Stress Response
3. Discussion
4. Materials and Methods
4.1. Plant Material
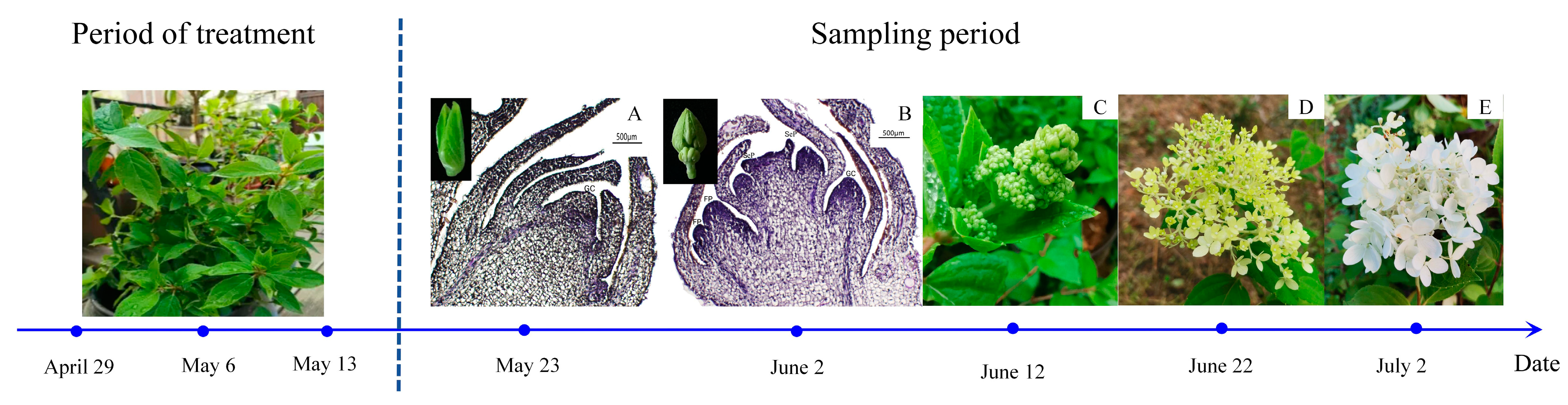
4.2. Plant Material Treatment and Sample Collection
4.3. Flowering Index Measurement
4.4. Physiological Index Measurement
4.5. RNA Extraction, Testing, and Transcriptome Sequencing
4.6. Data Assembly and Bioinformatics Analysis
4.7. qPCR Validation
4.8. Data Processing
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kitamura, Y.; Fudano, T.; Kawanishi, Y. Relation between floral initiations on apical buds of basal shoots after the autumn and cultivar differences in unseasonal flowering of hydrangea. Hortic. J. 2020, 89, 488–495. [Google Scholar] [CrossRef]
- Pagter, M.; Sergeant, K.; Moller, S.M.; Bertram, H.C.; Renaut, J. Changes in the proteome and water state in bark and xylem of Hydrangea paniculata during loss of freezing tolerance. Environ. Exp. Bot. 2014, 106, 99–111. [Google Scholar] [CrossRef]
- Greer, S.P.; Rinehart, T.A. Dormancy and germination in vitro response of hydrangea macrophylla and Hydrangea paniculata seed to light, cold-treatment and gibberellic acid. J. Environ. Hortic. 2010, 28, 41–47. [Google Scholar] [CrossRef]
- Cochran, D.R.; Fulcher, A. Type and rate of plant growth regulator influence vegetative, floral growth, and quality of little lime™ hydrangea. Horttechnology 2013, 23, 306–311. [Google Scholar] [CrossRef]
- Liu, F.; Huang, L.-L.; Li, Y.; Reinhoud, P.; Jongsma, M.A.; Wang, C.-Y. Shoot organogenesis in leaf explants of Hydrangea macrophylla ‘Hyd1’ and assessing genetic stability of regenerants using ISSR markers. Plant Cell Tissue Organ Cult. 2011, 104, 111–117. [Google Scholar] [CrossRef]
- Khaing, M.T.; Jung, H.J.J.; Kim, J.B.; Han, T.-H. Characterization of hydrangea accessions based on morphological and molecular markers. Hortic. Sci. Technol. 2018, 36, 598–605. [Google Scholar] [CrossRef]
- Sheng, J.; Li, X.; Zhang, D. Gibberellins, brassinolide, and ethylene signaling were involved in flower differentiation and development in Nelumbo nucifera. Hortic. Plant J. 2022, 8, 243–250. [Google Scholar] [CrossRef]
- Kumar, S.; Nanda, K.K. Effect of gibberellic acid and salicylic acid on the activities and electrophoretic patterns of alkaline and acid phosphatases during floral induction in impatiens balsamina. Z. Pflanzenphysiol. 1981, 101, 159–168. [Google Scholar] [CrossRef]
- An, C.; Mou, Z. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef]
- Gad, M.; Abdul-Hafeez, E.; Ibrahim, O. Foliar application of salicylic acid and gibberellic acid enhances growth and flowering of Ixora coccinea L. Plants. J. Plant Prod. 2016, 7, 85–91. [Google Scholar] [CrossRef]
- Saks, Y.; Van Staden, J. The role of gibberellic acid in the senescence of carnation flowers. J. Plant Physiol. 1992, 139, 484–488. [Google Scholar] [CrossRef]
- Alonso-Ramirez, A.; Rodriguez, D.; Reyes, D.; Angel Jimenez, J.; Nicolas, G.; Lopez-Climent, M.; Gomez-Cadenas, A.; Nicolas, C. Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in arabidopsis seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Sedighi, A.; Ershadi, A.; Sarikhani, H. Effect of pre- and postharvest treatments of salicylic and gibberellic acid on ripening and some physicochemical properties of ‘mashhad’ sweet cherry (Prunus avium L.) fruit. Acta Hort. 2010, 884, 257–264. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Khan, A.L.; Lee, I.-J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Masood, A.; Khan, M.I.R.; Fatma, M.; Asgher, M.; Per, T.S.; Khan, N.A. Involvement of ethylene in gibberellic acid-induced sulfur assimilation, photosynthetic responses, and alleviation of cadmium stress in mustard. Plant Physiol. Biochem. 2016, 104, 1–10. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Navarro, L.; Bari, R.; Jones, J.D.G. Pathological hormone imbalances. Curr. Opin. Plant Biol. 2007, 10, 372–379. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef]
- Chen, L.; Cao, T.; Zhang, J.; Lou, Y. Overexpression of osgid1 enhances the resistance of rice to the brown planthopper Nilaparvata lugens. Int. J. Mol. Sci. 2018, 19, 2744. [Google Scholar] [CrossRef]
- Nell, T.A.; Bodnaruk, W.H.; Joiner, J.N.; Sheehan, T.J. Ethylene evolution and flowering of cold- and GA-treated ‘Redwing’ azaleas. Hortscience 1983, 18, 454–455. [Google Scholar] [CrossRef]
- Pieterse, A.H. Is flowering in Lemnaceae stress-induced? A review. Aquat. Bot. 2013, 104, 1–4. [Google Scholar] [CrossRef]
- Elsadek, A. Improvement yield and quality of dahlia flowers by exogenous application of gibberellic acid and salicylic acid under sandy soil conditions. J. Plant Prod. 2018, 9, 289–297. [Google Scholar] [CrossRef]
- Khangjarakpam, G.; Mandal, S.; Singh, L.J. Influence of foliar application of gibberellic acid on growth, development, yield and biochemical constituents of African marigold cv. ‘Pusa Narangi Gainda’. J. Pharmacogn. Phytochem. 2019, 8, 1581–1585. [Google Scholar]
- Ayesha, R.; Hassan, I.; Abbasi, N.A.; Khan, K.S. Regulation of morpho-physiological and vase quality attributes of carnation (Dianthus caryophyllus) cv. ‘Tabasco’ mediated by GA3. Pak. J. Bot. 2020, 52, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Palevitch, D.; Milo, J.; Lavie, D. Effect of gibberellic acid on flowering and the thebaine yield of different clones of Papaver bracteatum. Plant Grouth Regul. 1986, 4, 153–157. [Google Scholar] [CrossRef]
- Angitha, S.; Singh, D.; Fatmi, U. Effect of plant growth regulators giberellic acid (GA3) and salicylic acid (SA) on growth and yield of carnation (Dianthus caryophyllus) under naturally ventilated polyhouse. Int. J. Plant Soil Sci. 2022, 34, 416–423. [Google Scholar]
- Glazinska, P.; Zienkiewicz, A.; Wojciechowski, W.; Kopcewicz, J. The putative miR172 target gene InAPETALA2-like is involved in the photoperiodic flower induction of Ipomoea nil. J. Plant Physiol. 2009, 166, 1801–1813. [Google Scholar] [CrossRef]
- Quebedeaux, B.; Beyer, E.M.J. Chemically induced parthenocarpy in cucumber by a new inhibitor of auxin transport. Hortscience 1972, 7, 474–476. [Google Scholar] [CrossRef]
- Christopher, D.A.; Loy, J.B. Influence of foliarly applied growth regulators on sex expression in watermelon. J. Am. Soc. Hortic. Sci. 1982, 107, 401–404. [Google Scholar] [CrossRef]
- Takahiro, H.; Royal, D.H.; Arthur, C.C.; William, H.C. Ethephon influences flowering, height, and branching of several herbaceous perennials. Sci. Hortic. 2001, 91, 305–324. [Google Scholar]
- Farazi, E.; Afshari, H.; Abadi, H.H. Effect of different concentrations of brassinosteroid on physiomorphological characteristics of five pistachio genotypes (Pistacia vera. L). J. Nuts 2015, 6, 143–153. [Google Scholar]
- Zhang, X.; Wang, S.; Ren, Y.; Gan, C.; Li, B.; Fan, Y.; Zhao, X.; Yuan, Z. Identification, analysis and gene cloning of the sweet gene family provide insights into sugar transport in pomegranate (Punica granatum). Int. J. Mol. Sci. 2022, 23, 2471. [Google Scholar] [CrossRef]
- Prat, L.; Botti, C.; Fichet, T. Effect of plant growth regulators on floral differentiation and seed production in Jojoba (Simmondsia chinensis (Link) Schneider). Ind. Crops Prod. 2008, 27, 44–49. [Google Scholar] [CrossRef]
- Aliyu, O.M.; Adeigbe, O.O.; Awopetu, J.A. Foliar application of the exogenous plant hormones at pre-blooming stage improves flowering and fruiting in cashew (Anacardium occidentale L.). J. Crop Sci. Biotechnol. 2011, 14, 143–150. [Google Scholar] [CrossRef]
- Kumar, P.; Singh, D.-V.; Johar, V.; Kumar, A. Uses of plant growth regulators and biofertilizers in fruit crops: A review. Int. J. Environ. Clim. Chang. 2022, 12, 314–326. [Google Scholar] [CrossRef]
- Patel, M.; Patel, R.; Chawla, S.; Patil, S.; Patel, D.K. Influence of ga3 and sa on growth and yield of Limonium var. misty blue. Int. J. Chem. Stud. 2018, 6, 1575–1577. [Google Scholar]
- Naor, V.; Kigel, J.; Ziv, M.; Flaishman, M. A developmental pattern of flowering in colored Zantedeschia spp.: Effects of bud position and gibberellin. J. Plant Growth Regul. 2005, 23, 269–279. [Google Scholar] [CrossRef]
- Chen, W.S.; Liu, H.Y.; Liu, Z.H.; Yang, L.N.; Chen, W.H. Gibberellin and temperature influence carbohydrate content and flowering in Phalaenopsis. Physiol. Plant 1994, 90, 391–395. [Google Scholar] [CrossRef]
- Cheng, C.; Jiao, C.; Singer, S.D.; Gao, M.; Xu, X.; Zhou, Y.; Li, Z.; Fei, Z.; Wang, Y.; Wang, X. Gibberellin-induced changes in the transcriptome of grapevine (Vitis labrusca × V. vinifera) cv. Kyoho flowers. BMC Genom. 2015, 16, 128. [Google Scholar] [CrossRef]
- Paroussin, G.; Voyiatzis, D.G.; Parousis, E.; Drogoudia, P.D. Growth, flowering and yield responses to GA3 of strawberry grown under different environmental conditions. Sci. Hortic. 2002, 96, 103–113. [Google Scholar] [CrossRef]
- Kepinski, S. Integrating hormone signaling and patterning mechanisms in plant development. Curr. Opin. Plant Biol. 2006, 9, 28–34. [Google Scholar] [CrossRef]
- Munoz-Fambuena, N.; Mesejo, C.; Carmen Gonzalez-Mas, M.; Iglesias, D.J.; Primo-Millo, E.; Agusti, M. Gibberellic acid reduces flowering intensity in sweet orange [Citrus sinensis (L.) osbeck] by repressing cift gene expression. J. Plant Growth Regul. 2012, 31, 529–536. [Google Scholar] [CrossRef]
- Sabaghian, E.; Drebert, Z.; Inze, D.; Saeys, Y. An integrated network of Arabidopsis growth regulators and its use for gene prioritization. Sci. Rep. 2015, 5, 17617. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bao, Z.; Clancy, M.A.; Carvalho, R.F.; Elliott, K.; Folta, K.M. Identification of novel growth regulators in plant populations expressing random peptides. Plant Physiol. 2017, 175, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Ebinezer, L.B.; Franchin, C.; Trentin, A.R.; Carletti, P.; Trevisan, S.; Agrawal, G.K.; Rakwal, R.; Quaggiotti, S.; Arrigoni, G.; Masi, A. Quantitative proteomics of maize roots treated with a protein hydrolysate: A comparative study with transcriptomics highlights the molecular mechanisms responsive to biostimulants. J. Agric. Food Chem. 2020, 68, 7541–7553. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Gray, W.M. Plant hormone receptors: New perceptions. Genes Dev. 2008, 22, 2139–2148. [Google Scholar] [CrossRef] [PubMed]
- Biesaga-Koscielniak, J.; Koscielniak, J.; Filek, M.; Marcinska, I.; Krekule, J.; Machackova, I.; Kubon, M. The effect of plant growth regulators and their interaction with electric current on winter wheat development. Acta Physiol. Plant. 2010, 32, 987–995. [Google Scholar] [CrossRef]
- Davies, P.J. Plant hormones: Biosynthesis, signal transduction, action! S. Afr. J. Bot. 2006, 72, 185–194. [Google Scholar]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of Salinity-Induced Oxidative Stress in Plants by Salicylic Acid: Physiological and Biochemical Insights. In Salicylic Acid: A Multifaceted Hormone; Springer: Berlin/Heidelberg, Germany, 2020; pp. 179–198. [Google Scholar]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB Transcription Factor Genes as Regulators for Plant Responses: An Overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nie, Y.; Wang, D.; Xie, L. Effects of ETH and PP333 on the growth, florescence and physiological properties of bougainvillea spectabilis. Sci. Silvae Sin. 2018, 54, 46–55. [Google Scholar]
- Sun, Q.; Yu, X.M.; Hou, X.F.; Sun, J.J. Study on characteristics for tests guidelines of distinctness, unifornity and stability for varieties of hydrangea. In Advances in Ornamental Horticulture of China; China Forestry Publishing House: Beijiing, China, 2018; pp. 106–110. [Google Scholar]
- Wang, H.M.; Yang, S.; Huang, H.R.; Ding, L.M.; Fei, Y.; Wu, Q. Ornamental property of cultivated varieties of Hydrangea paniculata. Fujian J. Agric. Sci. 2020, 35, 286–294. [Google Scholar]
- Ling, J.; Tan, J.; Chen, H.; Yang, Z.; Luo, Q.; Jia, J. Physiology, transcriptome and root exudates analysis of response to aluminum stress in Pinus massoniana. Forests 2023, 14, 1410. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Peng, J.; Xue, C.; Dong, X.; Zeng, C.; Wu, Y.; Cao, F. Gene cloning and analysis of the pattern of expression of the transcription factor HymMYB2 related to blue flower formation in Hydrangea macrophylla. Euphytica 2021, 217, 115. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D.L. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Initial Flowering Date | Full Flowering Date | Final Flowering Date | Days Advanced for Initial Flowering (d) | Increased Days of Total Flowering Duration (d) |
|---|---|---|---|---|---|
| CK | 11 June 2021 | 18 June 2021 | 29 August 2021 | 0 ± 1.89 b | 0 ± 1.70 b |
| G1 | 14 June 2021 | 21 June 2021 | 29 August 2021 | −3 ± 1.41 b | −3 ± 1.24 b |
| G2 | 20 June 2021 | 28 June 2021 | 27 August 2021 | −9 ± 0.67 d | −11 ± 0.82 c |
| G3 | 23 June 2021 | 30 June 2021 | 19 August 2021 | −12 ± 1.76 e | −21 ± 1.49 d |
| S1 | 6 June 2021 | 12 June 2021 | 4 September 2021 | 5 ± 0.47 a | 10 ± 0.82 a |
| S2 | 17 June 2021 | 25 June 2021 | 23 August 2021 | −6 ± 1.15 c | −11 ± 0.99 c |
| S3 | 20 June 2021 | 27 June 2021 | 20 August 2021 | −9 ± 0.94 d | −17 ± 1.05 e |
| E1 | 21 June 2021 | 27 June 2021 | 26 August 2021 | −10 ± 0.67 de | −12 ± 0.82 c |
| E2 | 22 June 2021 | 30 June 2021 | 27 August 2021 | −11 ± 0.82 de | −12 ± 1.05 c |
| E3 | 20 June 2021 | 27 June 2021 | 26 August 2021 | −9 ± 1.15 d | −11 ± 1.25 c |
| Treatment | Inflorescence Length (mm) | Inflorescence Width (mm) | Number of Inflorescences | Petal Area (cm2) |
|---|---|---|---|---|
| CK | 123.67 ± 1.25 c | 99.67 ± 2.05 c | 12 ± 0.99 d | 1.53 ± 0.02 b |
| G1 | 133.00 ± 2.16 b | 111.67 ± 2.87 b | 9 ± 0.82 e | 1.57 ± 0.11 b |
| G2 | 118.67 ± 4.50 cd | 97.33 ± 4.71 cd | 10 ± 1.15 de | 1.42 ± 0.09 b |
| G3 | 110.00 ± 8.16 d | 93.33 ± 2.49 d | 12 ± 1.15 d | 1.02 ± 0.04 c |
| S1 | 151.67 ± 2.36 a | 126.33 ± 1.25 a | 12 ± 0.82 d | 1.95 ± 0.08 a |
| S2 | 113.00 ± 2.16 d | 91.67 ± 2.36 d | 13 ± 1.05 d | 1.12 ± 0.03 c |
| S3 | 94.00 ± 0.94 e | 71.00 ± 1.41 f | 15 ± 0.47 c | 0.65 ± 0.03 e |
| E1 | 92.00 ± 2.45 e | 81.67 ± 2.36 e | 19 ± 0.82 a | 0.75 ± 0.02 de |
| E2 | 93.33 ± 2.49 e | 85.67 ± 4.19 e | 17 ± 1.05 b | 0.87 ± 0.03 d |
| E3 | 119.37 ± 4.19 cd | 98.67 ± 1.89 cd | 12 ± 1.15 d | 1.43 ± 0.09 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, C.; Wen, Y.; Sheng, S.; Gao, Y.; Zhang, Y.; Chen, T.; Peng, J.; Cao, S. Hormonal Regulation and Transcriptomic Insights into Flower Development in Hydrangea paniculata ‘Vanilla Strawberry’. Plants 2024, 13, 486. https://doi.org/10.3390/plants13040486
Xue C, Wen Y, Sheng S, Gao Y, Zhang Y, Chen T, Peng J, Cao S. Hormonal Regulation and Transcriptomic Insights into Flower Development in Hydrangea paniculata ‘Vanilla Strawberry’. Plants. 2024; 13(4):486. https://doi.org/10.3390/plants13040486
Chicago/Turabian StyleXue, Chao, Yuxing Wen, Song Sheng, Yu Gao, Yaoyi Zhang, Tingfeng Chen, Jiqing Peng, and Shoujin Cao. 2024. "Hormonal Regulation and Transcriptomic Insights into Flower Development in Hydrangea paniculata ‘Vanilla Strawberry’" Plants 13, no. 4: 486. https://doi.org/10.3390/plants13040486
APA StyleXue, C., Wen, Y., Sheng, S., Gao, Y., Zhang, Y., Chen, T., Peng, J., & Cao, S. (2024). Hormonal Regulation and Transcriptomic Insights into Flower Development in Hydrangea paniculata ‘Vanilla Strawberry’. Plants, 13(4), 486. https://doi.org/10.3390/plants13040486