


Aphid Resistance Segregates Independently of Cardenolide and Glucosinolate Content in an Erysimum cheiranthoides (Wormseed Wallflower) F2 Population

 , , , , and
, , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Phenotypic Differences between the Elbtalaue and Konstanz Accessions
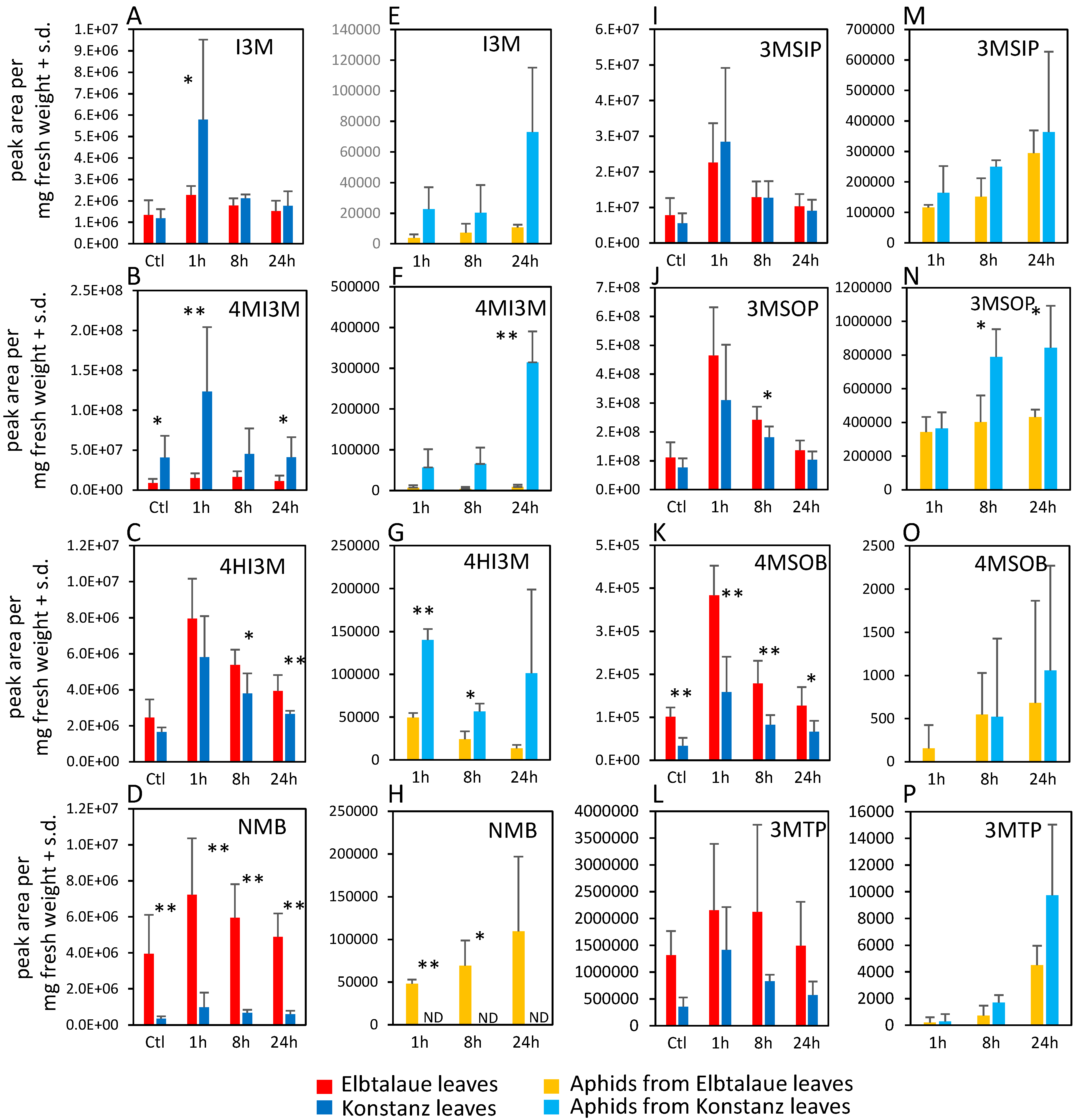
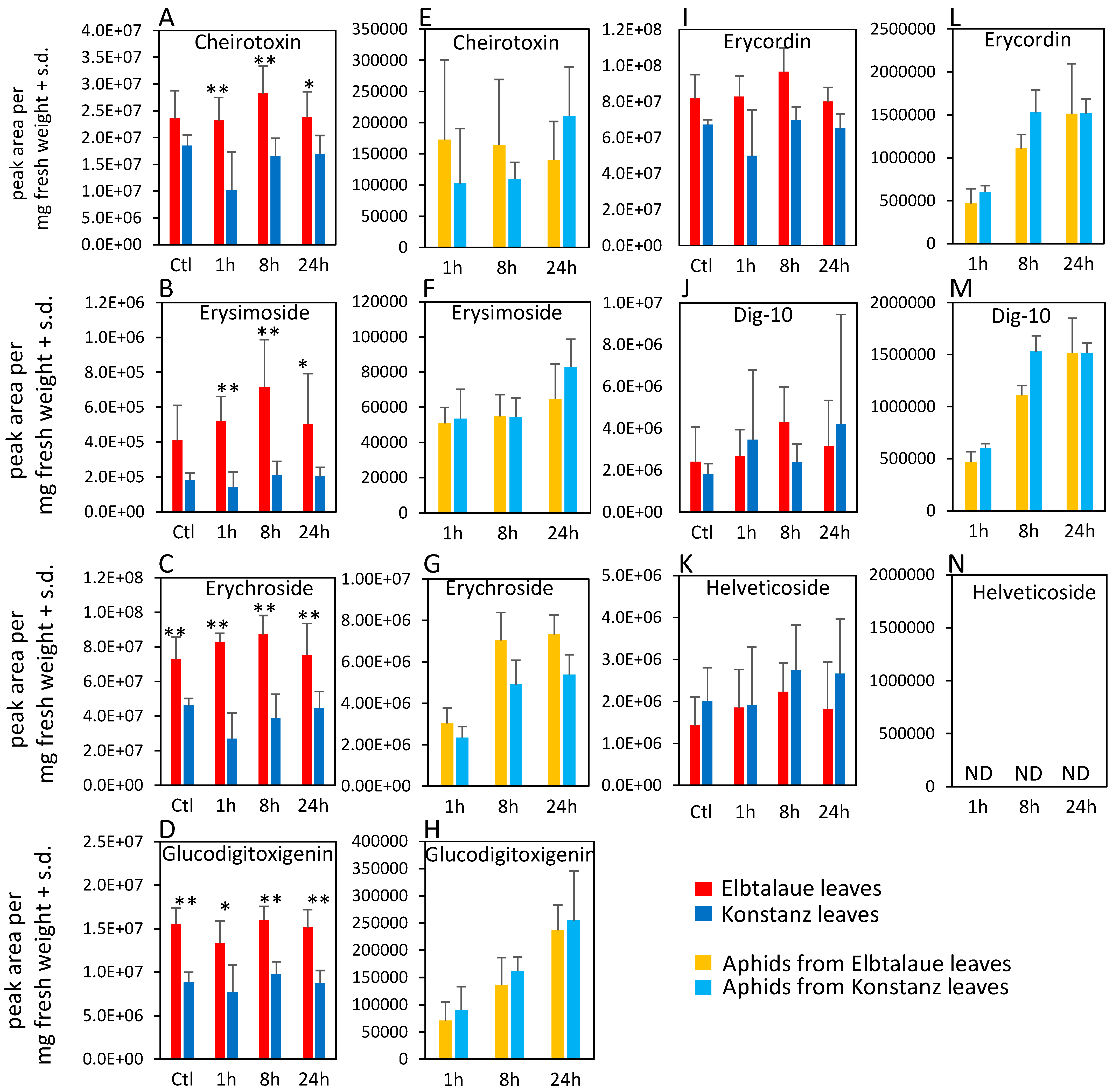
2.2. Correlation of Aphid Resistance with Glucosinolate and Cardenolide Content
2.3. E. cheiranthoides Genetic Map
2.4. Genetic Mapping of Defense Traits
3. Discussion
4. Materials and Methods
4.1. Plant and Insect Rearing
4.2. Insect Bioassays
4.3. Detection of Glucosinolates and Cardenolides
4.4. Transcriptome Sequencing
4.5. Genetic Map Construction and Assembly of E. cheiranthoides Genome v2.0
4.6. Genome Annotation
4.7. Coexpression Network Analysis
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Edger, P.P.; Heidel-Fischer, H.M.; Bekaert, M.; Rota, J.; Glockner, G.; Platts, A.E.; Heckel, D.G.; Der, J.P.; Wafula, E.K.; Tang, M.; et al. The butterfly plant arms-race escalated by gene and genome duplications. Proc. Natl. Acad. Sci. USA 2015, 112, 8362–8366. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Agerbirk, N.; Stauber, E.J.; Olsen, C.E.; Hippler, M.; Mitchell-Olds, T.; Gershenzon, J.; Vogel, H. Successful herbivore attack due to metabolic diversion of a plant chemical defense. Proc. Natl. Acad. Sci. USA 2004, 101, 4859–4864. [Google Scholar] [CrossRef]
- Ratzka, A.; Vogel, H.; Kliebenstein, D.J.; Mitchell-Olds, T.; Kroymann, J. Disarming the mustard oil bomb. Proc. Natl. Acad. Sci. USA 2002, 99, 11223–11228. [Google Scholar] [CrossRef]
- Francis, F.; Lognay, G.; Wathelet, J.P.; Haubruge, E. Characterisation of aphid myrosinase and degradation studies of glucosinolates. Arch. Insect Biochem. Physiol. 2002, 50, 173–182. [Google Scholar] [CrossRef]
- Jones, A.M.; Bridges, M.; Bones, A.M.; Cole, R.; Rossiter, J.T. Purification and characterisation of a non-plant myrosinase from the cabbage aphid Brevicoryne brassicae (L.). Insect Biochem. Mol. Biol. 2001, 31, 1–5. [Google Scholar] [CrossRef]
- Pontoppidan, B.; Ekbom, B.; Eriksson, S.; Meijer, J. Purification and characterization of myrosinase from the cabbage aphid (Brevicoryne brassicae), a Brassica herbivore. Eur. J. Biochem. 2001, 268, 1041–1048. [Google Scholar] [CrossRef]
- Müller, C.; Wittstock, U. Uptake and turn-over of glucosinolates sequestered in the sawfly Athalia rosae. Insect Biochem. Mol. Biol. 2005, 35, 1189–1198. [Google Scholar] [CrossRef]
- Beran, F.; Pauchet, Y.; Kunert, G.; Reichelt, M.; Wielsch, N.; Vogel, H.; Reinecke, A.; Svatos, A.; Mewis, I.; Schmid, D.; et al. Phyllotreta striolata flea beetles use host plant defense compounds to create their own glucosinolate-myrosinase system. Proc. Natl. Acad. Sci. USA 2014, 111, 7349–7354. [Google Scholar] [CrossRef] [PubMed]
- Müller, R.; De Vos, M.; Sun, J.Y.; Sønderby, I.E.; Halkier, B.A.; Wittstock, U.; Jander, G. Differential effects of indole and aliphatic glucosinolates on lepidopteran herbivores. J. Chem. Ecol. 2010, 36, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jander, G. Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a deterrent indole glucosinolate. Plant J. 2007, 49, 1008–1019. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops; Wiley: Chichester, UK, 2000; p. 466. [Google Scholar]
- Makarevich, F.I.; Zhernoklev, K.V.; Slyusarskaya, T.B.; Yarmolenko, G.N. Cardenolide-containing plants of the family Cruciferae. Chem. Nat. Comp. 1994, 30, 275–289. [Google Scholar] [CrossRef]
- Züst, T.; Mirzaei, M.; Jander, G. Erysimum cheiranthoides, an ecological research system with potential as a genetic and genomic model for studying cardiac glycoside biosynthesis. Phytochem. Rev. 2018, 17, 1239–1251. [Google Scholar] [CrossRef]
- Younkin, G.C.; Alani, M.L.; Capador, A.P.; Fischer, H.D.; Mirzaei, M.; Hastings, A.P.; Agrawal, A.A.; Jander, G. Cardiac glycosides protect wormseed wallflower (Erysimum cheiranthoides) against some, but not all, glucosinolate-adapted herbivores. New Phytol. 2024; advance online publication. [Google Scholar] [CrossRef]
- Lei, Z.H.; Nakayama, H.; Kuniyasu, A.; Tai, B.S.; Nohara, T. Cardiac glycosides from Erysimum cheiranthoides. Chem. Pharm. Bull. 2002, 50, 861–862. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lei, Z.H.; Kuniyasu, A.; Tai, B.S.; Nakayama, H.; Nohara, T. Na+,K+-ATPase inhibiting activity of cardiac glycosides from Erysimum cheiranthoides. Planta Med. 2001, 67, 369–370. [Google Scholar] [CrossRef]
- Kreis, W. The foxgloves (Digitalis) revisited. Planta Med. 2017, 83, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A. Monarchs and Milkweed: A Migrating Butterfly, a Poisonous Plant, and Their Remarkable Story of Coevolution; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Munkert, J.; Costa, C.; Budeanu, O.; Petersen, J.; Bertolucci, S.; Fischer, G.; Muller-Uri, F.; Kreis, W. Progesterone 5beta-reductase genes of the Brassicaceae family as function-associated molecular markers. Plant Biol. 2015, 17, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Nagata, W.; Tamm, C.; Reichstein, T. Die Glykoside von Erysimum crepidifolium HGL Reichenbach. Glykoside und Aglykone 169. Mitteilung. Helv. Chim. Acta 1957, 40, 41–61. [Google Scholar] [CrossRef]
- Moazzeni, H.; Zarre, S.; Pfeil, B.E.; Bertrand, Y.J.K.; German, D.A.; Al-Shehbaz, I.A.; Mummenhoff, K.; Oxelman, B. Phylogenetic perspectives on diversification and character evolution in the species-rich genus Erysimum (Erysimeae; Brassicaceae) based on a densely sampled ITS approach. Bot. J. Linn. Soc. 2014, 175, 497–522. [Google Scholar] [CrossRef]
- Züst, T.; Strickler, S.R.; Powell, A.F.; Mabry, M.E.; An, H.; Mirzaei, M.; York, T.; Holland, C.K.; Kumar, P.; Erb, M.; et al. Rapid and independent evolution of ancestral and novel defenses in a genus of toxic plants (Erysimum, Brassicaceae). eLife 2020, 9, e51712. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Blazevic, I.; Montaut, S.; Burcul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Agrawal, A.A. Natural selection on common milkweed (Asclepias syriaca) by a community of specialized insect herbivores. Evol. Ecol. Res. 2005, 7, 651–667. [Google Scholar]
- Huang, X.; Renwick, J.A.; Sachdev-Gupta, K. A chemical basis for differential acceptance of Erysimum cheiranthoides by two Pieris species. J. Chem. Ecol. 1993, 19, 195–210. [Google Scholar] [CrossRef]
- Huang, X.P.; Renwick, J.A.A. Differential selection of host plants by two Pieris species: The role of oviposition stimulants and deterrents. Ent. Exp. Appl. 1993, 68, 59–69. [Google Scholar] [CrossRef]
- Feeny, P. Ecology of the Cruciferae. Ann. Mo. Bot. Gard. 1977, 64, 221–234. [Google Scholar] [CrossRef]
- Chew, F.S. Coevolution of pierid butterflies and their cruciferous foodplants. 2. Distribution of eggs on potential foodplants. Evolution 1977, 31, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Chew, F.S. Coevolution of pierid butterflies and their cruciferous food plants. 1. Relative quality of available resources. Oecologia 1975, 20, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Renwick, J.A.; Radke, C.D.; Sachdev-Gupta, K. Chemical constituents of Erysimum cheiranthoides deterring oviposition by the cabbage butterfly, Pieris rapae. J. Chem. Ecol. 1989, 15, 2161–2169. [Google Scholar] [CrossRef] [PubMed]
- Sachdev-Gupta, K.; Renwick, J.A.; Radke, C.D. Isolation and identification of oviposition deterrents to cabbage butterfly, Pieris rapae, from Erysimum cheiranthoides. J. Chem. Ecol. 1990, 16, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Mertens, D.; Bouwmeester, K.; Poelman, E.H. Intraspecific variation in plant-associated herbivore communities is phylogenetically structured in Brassicaceae. Ecol. Lett. 2021, 24, 2314–2327. [Google Scholar] [CrossRef]
- Mirzaei, M.; Züst, T.; Younkin, G.C.; Hastings, A.P.; Alani, M.L.; Agrawal, A.A.; Jander, G. Less is more: A mutation in the chemical defense pathway of Erysimum cheiranthoides (Brassicaceae) reduces total cardenolide abundance but increases resistance to insect herbivores. J. Chem. Ecol. 2020, 46, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, B.W.; Schroeder, F.C.; Jander, G. Identification of indole glucosinolate breakdown products with antifeedant effects on Myzus persicae (green peach aphid). Plant J. 2008, 54, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hull, A.K.; Gupta, N.R.; Goss, K.A.; Alonso, J.; Ecker, J.R.; Normanly, J.; Chory, J.; Celenza, J.L. Trp-dependent auxin biosynthesis in Arabidopsis: Involvement of cytochrome P450s CYP79B2 and CYP79B3. Genes Dev. 2002, 16, 3100–3112. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Jander, G. Arabidopsis myrosinases TGG1 and TGG2 have redundant function in glucosinolate breakdown and insect defense. Plant J. 2006, 46, 549–562. [Google Scholar] [CrossRef]
- Zhang, Q.; Du, Y.; Bai, S.; Yuan, W. Complete chloroplast genome sequence and phylogenetic analysis of Erysimum cheiranthoides. Mitochondrial DNA B Resour. 2019, 4, 2920–2921. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.V.; Song, B.H.; Olson-Manning, C.; Anderson, J.T.; Lee, C.R.; Schranz, M.E.; Windsor, A.J.; Clauss, M.J.; Manzaneda, A.J.; Naqvi, I.; et al. A gain-of-function polymorphism controlling complex traits and fitness in nature. Science 2012, 337, 1081–1084. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Borowsky, A.T.; Tzin, V.; Jander, G.; Kliebenstein, D.J.; Rokas, A. A global coexpression network approach for connecting genes to specialized metabolic pathways in plants. Plant Cell 2017, 29, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Sønderby, I.E.; Burow, M.; Rowe, H.C.; Kliebenstein, D.J.; Halkier, B.A. A complex interplay of three R2R3 MYB transcription factors determines the profile of aliphatic glucosinolates in Arabidopsis. Plant Physiol. 2010, 153, 348–363. [Google Scholar] [CrossRef]
- Pfalz, M.; Vogel, H.; Kroymann, J. The gene controlling the indole glucosinolate modifier1 quantitative trait locus alters indole glucosinolate structures and aphid resistance in Arabidopsis. Plant Cell 2009, 21, 985–999. [Google Scholar] [CrossRef]
- Pfalz, M.; Mikkelsen, M.D.; Bednarek, P.; Olsen, C.E.; Halkier, B.A.; Kroymann, J. Metabolic engineering in Nicotiana benthamiana reveals key enzyme functions in Arabidopsis indole glucosinolate modification. Plant Cell 2011, 23, 716–729. [Google Scholar] [CrossRef]
- Pfalz, M.; Mukhaimar, M.; Perreau, F.; Kirk, J.; Hansen, C.I.; Olsen, C.E.; Agerbirk, N.; Kroymann, J. Methyl transfer in glucosinolate biosynthesis mediated by indole glucosinolate O-methyltransferase 5. Plant Physiol. 2016, 172, 2190–2203. [Google Scholar] [CrossRef]
- Hansen, B.G.; Kliebenstein, D.J.; Halkier, B.A. Identification of a flavin-monooxygenase as the S-oxygenating enzyme in aliphatic glucosinolate biosynthesis in Arabidopsis. Plant J. 2007, 50, 902–910. [Google Scholar] [CrossRef]
- Li, J.; Hansen, B.G.; Ober, J.A.; Kliebenstein, D.J.; Halkier, B.A. Subclade of flavin-monooxygenases involved in aliphatic glucosinolate biosynthesis. Plant Physiol. 2008, 148, 1721–1733. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Goggin, F.L.; Milligan, S.B.; Kaloshian, I.; Ullman, D.E.; Williamson, V.M. The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc. Natl. Acad. Sci. USA 1998, 95, 9750–9754. [Google Scholar] [CrossRef] [PubMed]
- Dogimont, C.; Chovelon, V.; Pauquet, J.; Boualem, A.; Bendahmane, A. The Vat locus encodes for a CC-NBS-LRR protein that confers resistance to Aphis gossypii infestation and A. gossypii-mediated virus resistance. Plant J. 2014, 80, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Oswald, S.; Zollner, U. Die Wirtseinigung von Rapssorten unterschiedlichen Glucosinolatgehalts fuer Brevicoryne brassicae (L.) und Myzus persicae (Sulzer) (Hemiptera, Aphididae). Z. Pflanzenkrankh. Pflanzenschutz 1986, 93, 113–124. [Google Scholar]
- Feng, H.; Chen, W.; Hussain, S.; Shakir, S.; Tzin, V.; Adegbayi, F.; Ugine, T.; Fei, Z.; Jander, G. Horizontally transferred genes as RNA interference targets for aphid and whitefly control. Plant Biotechnol. J. 2023, 21, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Febvay, G.; Bonnin, J.; Rahbe, Y.; Bournoville, R.; Delrot, S.; Bonnemain, J.L. Resistance of different lucerne cultivars to the pea aphid Acyrthosiphon pisum: Influence of phloem composition on aphid fecundity. Ent. Exp. Appl. 1988, 48, 127–134. [Google Scholar] [CrossRef]
- Douglas, A.E.; Minto, L.B.; Wilkinson, T.L. Quantifying nutrient production by the microbial symbionts in an aphid. J. Exp. Biol. 2001, 204, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Kremling, K.A.G.; Chen, S.Y.; Su, M.H.; Lepak, N.K.; Romay, M.C.; Swarts, K.L.; Lu, F.; Lorant, A.; Bradbury, P.J.; Buckler, E.S. Dysregulation of expression correlates with rare-allele burden and fitness loss in maize. Nature 2018, 555, 520–523. [Google Scholar] [CrossRef]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high confidence variant calls: The Genome Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.11–11.10.33. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Gonda, I.; Ashrafi, H.; Lyon, D.A.; Strickler, S.R.; Hulse-Kemp, A.M.; Ma, Q.; Sun, H.; Stoffel, K.; Powell, A.F.; Futrell, S.; et al. Sequencing-based bin map construction of a tomato mapping population, facilitating high-resolution quantitative trait loci detection. Plant Genome 2019, 12, 180010. [Google Scholar] [CrossRef]
- Wu, Y.; Bhat, P.R.; Close, T.J.; Lonardi, S. Efficient and accurate construction of genetic linkage maps from the minimum spanning tree of a graph. PLoS Genet. 2008, 4, e1000212. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Soorni, A.; Haak, D.; Zaitlin, D.; Bombarely, A. Organelle_PBA, a pipeline for assembling chloroplast and mitochondrial genomes from PacBio DNA sequencing data. BMC Genom. 2017, 18, 49. [Google Scholar] [CrossRef]
- Zhang, Y.; An, D.; Li, C.; Zhao, Z.; Wang, W. The complete chloroplast genome of greater duckweed (Spirodela polyrhiza 7498) using PacBio long reads: Insights into the chloroplast evolution and transcription regulation. BMC Genom. 2020, 21, 76. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 23 December 2023).
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef]
- Wu, T.D.; Watanabe, C.K. GMAP: A genomic mapping and alignment program for mRNA and EST sequences. Bioinformatics 2005, 21, 1859–1875. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Pertea, M. GffRead and GffCompare [version 2; peer review: 3 approved]. RF1000Research 2020, 9, 304. [Google Scholar] [CrossRef]
- Managadze, D. App to Add Explicit UTR Exon Records to GFF3 Data, Version 1.0.0. Available online: https://ftp.ncbi.nlm.nih.gov/genomes/TOOLS/add_utrs_to_gff/add_utrs_to_gff.py (accessed on 12 January 2024).
- Strickler, S.R.; Powell, A.F.; Mueller, L.A.; Zust, T.; Jander, G. NCBI BioProject ID PRJNA563696. Rapid and Independent Evolution of Ancestral and Novel Chemical Defenses in a Genus of Toxic Plants (Erysimum, Brassicaceae). Available online: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA563696/ (accessed on 23 December 2023).
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Morris, J.H.; Demchak, B.; Bader, G.D. Biological network exploration with Cytoscape 3. Curr. Protoc. Bioinforma. 2014, 47, 8.13.1–8.13.24. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.M.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer; Version 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2012; Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 23 December 2023).
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Templeton, G.F. A two-step approach for transforming continuous variables to normal: Implications and recommendations for IS research. Commun. Assoc. Inf. Syst. 2011, 28, 4. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirzaei, M.; Younkin, G.C.; Powell, A.F.; Alani, M.L.; Strickler, S.R.; Jander, G. Aphid Resistance Segregates Independently of Cardenolide and Glucosinolate Content in an Erysimum cheiranthoides (Wormseed Wallflower) F2 Population. Plants 2024, 13, 466. https://doi.org/10.3390/plants13040466
Mirzaei M, Younkin GC, Powell AF, Alani ML, Strickler SR, Jander G. Aphid Resistance Segregates Independently of Cardenolide and Glucosinolate Content in an Erysimum cheiranthoides (Wormseed Wallflower) F2 Population. Plants. 2024; 13(4):466. https://doi.org/10.3390/plants13040466
Chicago/Turabian StyleMirzaei, Mahdieh, Gordon C. Younkin, Adrian F. Powell, Martin L. Alani, Susan R. Strickler, and Georg Jander. 2024. "Aphid Resistance Segregates Independently of Cardenolide and Glucosinolate Content in an Erysimum cheiranthoides (Wormseed Wallflower) F2 Population" Plants 13, no. 4: 466. https://doi.org/10.3390/plants13040466
APA StyleMirzaei, M., Younkin, G. C., Powell, A. F., Alani, M. L., Strickler, S. R., & Jander, G. (2024). Aphid Resistance Segregates Independently of Cardenolide and Glucosinolate Content in an Erysimum cheiranthoides (Wormseed Wallflower) F2 Population. Plants, 13(4), 466. https://doi.org/10.3390/plants13040466