
Essential Oils Obtained from Sicilian Citrus reticulata Blanco By-Products: Antibacterial and Allelopathic Activity


 ,
,  ,
,  ,
, 

Abstract
1. Introduction
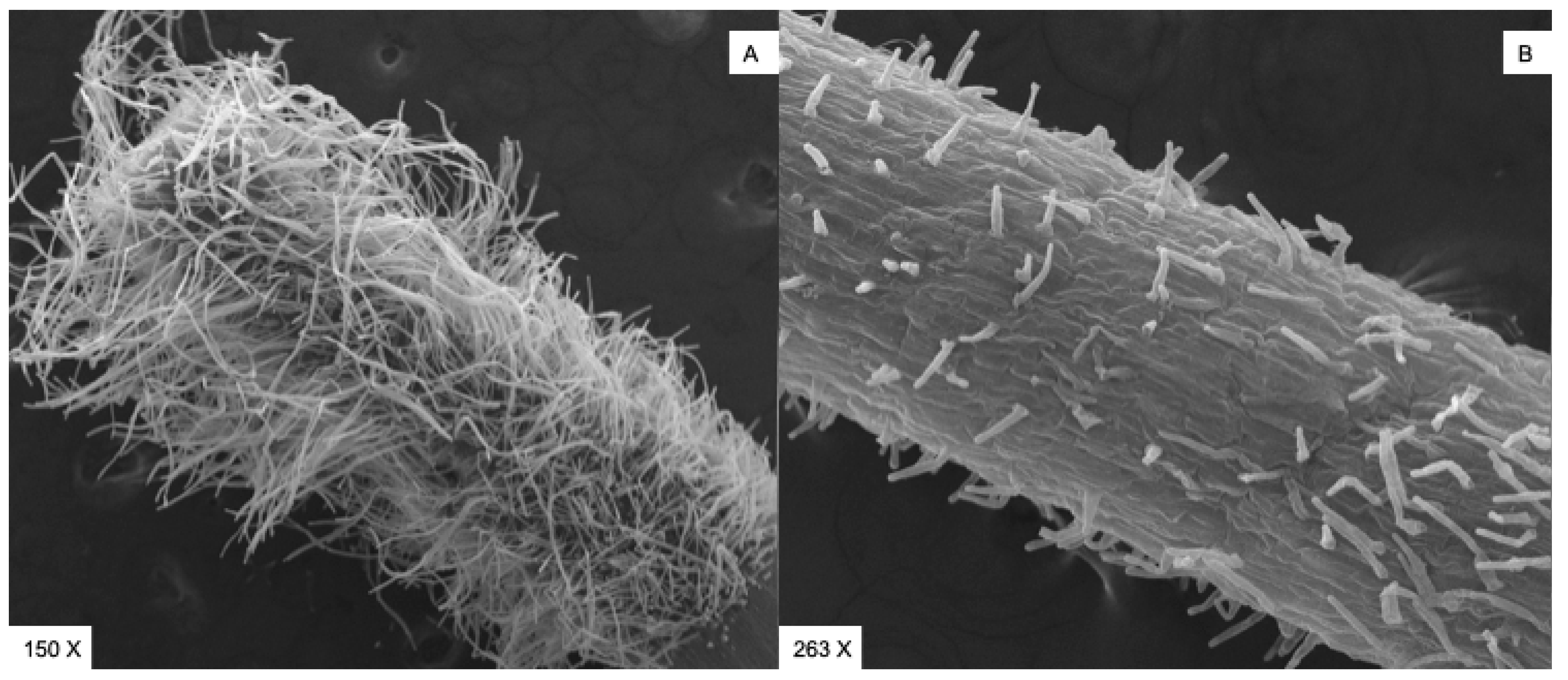
2. Results and Discussion
2.1. Chemical Composition of EOs (C1–C5)
2.2. Antibacterial Activity
2.3. Allelopathic Activity
2.3.1. Raphanus Sativus Seed Germination
2.3.2. Moss Spore Germination
3. Materials and Methods
3.1. Plant Materials
3.2. Extraction of EOs
3.3. GC and GC-MS Analyses
3.4. Antimicrobial Assay
3.5. Allelopathic Test
3.6. Seed Germination Tests
3.7. Moss Spore Germination Test
3.8. Evaluation of Oil Effects
3.9. Scanning Electron Microscopy
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding

Data Availability Statement
Conflicts of Interest
References
- Moulehi, I.; Bourgou, S.; Ourghemmi, I.; Tounsi, M.S. Variety and ripening impact on phenolic composition and antioxidant activity of mandarin (Citrus reticulate Blanco) and bitter orange (Citrus aurantium L.) seeds extracts. Ind. Crops Prod. 2012, 39, 74–80. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Heying, E.; Tanumihardjo, S.A. History, Global Distribution, and Nutritional Importance of Citrus Fruits. Compr. Rev. Food Sci. Food Saf. 2012, 11, 530–545. [Google Scholar] [CrossRef]
- Costanzo, G.; Vitale, E.; Iesce, M.R.; Naviglio, D.; Amoresano, A.; Fontanarosa, C.; Spinelli, M.; Ciaravolo, M.; Arena, C. Antioxidant Properties of Pulp, Peel and Seeds of Phlegrean Mandarin (Citrus reticulata Blanco) at Different Stages of Fruit Ripening. Antioxidants 2022, 11, 187. [Google Scholar] [CrossRef] [PubMed]
- Bentahar, A.; Bouaziz, A.; Djidel, S.; Khennouf, S. Phenolic content and antioxidant activity of ethanolic extracts from Citrus sinensis L. and Citrus reticulata L. fruits. J. Drug Deliv. Therap. 2020, 10, 308–313. [Google Scholar] [CrossRef]
- Nicolosi, E. Origin and taxonomy. In Citrus Genetics, Breeding and Biotechnology; Khan, I.A., Ed.; CAB International: Oxfordshire, UK, 2007; pp. 19–44. [Google Scholar]
- Luro, F.; Curk, F.; Froelicher, Y.; Ollitrault, P. Recent insights on Citrus diversity and phylogeny. In AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean: Acclimatization, Diversifications, Uses [Online]; Publications du Centre Jean Bérard: Naples, Italy, 2017; ISBN 9782918887775. [Google Scholar] [CrossRef]
- Ollitrault, P.; Curk, F.; Krueger, R. The Genus Citrus. In Citrus Taxonomy; Manuel Talon, M., Marco Caruso, M., Fred, G., Gmitter, F.G., Jr., Eds.; Woodhead Publishing: Cambridge, UK, 2020. [Google Scholar] [CrossRef]
- Badalamenti, N.; Bruno, M.; Schicchi, R.; Geraci, A.; Leporini, M.; Tundis, R.; Loizzo, M.R. Reuse of Food Waste: The Chemical Composition and Health Properties of Pomelo (Citrus maxima) Cultivar Essential Oils. Molecules 2022, 27, 3273. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Curk, F.; Ancillo, G.; Ollitrault, F.; Perrier, X.; Jacquemoud-Collet, J.-P.; Garcia-Lor, A.; Navarro, L.; Ollitrault, P. Nuclear Species-Diagnostic SNP Markers Mined from 454 Amplicon Sequencing Reveal Admixture Genomic Structure of Modern Citrus Varieties. PLoS ONE 2015, 10, e0125628. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lor, A.; Luro, F.; Ollitrault, P.; Navarro, L. Genetic diversity and population-structure analysis of mandarin germplasm by nuclear, chloroplastic and mitochondrial markers. Tree Genet. Genomes 2015, 11, 123. [Google Scholar] [CrossRef]
- Webber, H.J. History and development of the citrus industry. In The Citrus Industry; Batchelor, L.D., Webber, H.J., Eds.; University of California Press: Berkeley, CA, USA, 1943. [Google Scholar]
- Ciriminna, R.; Angellotti, G.; Luque, R.; Pagliaro, M. The citrus economy in Sicily in the early bioeconomy era: A case study for bioeconomy practitioners. Biofuels Bioprod. Bioref. 2024, 18, 356–364. [Google Scholar] [CrossRef]
- Lo Piccolo, F. Palermo Dominante. Il soggiorno dei Borbone alla Favorita e a Boccadifalco (1798–1820); 40due Edizioni: Palermo, Italy, 2024; p. 368. ISBN ISBN 88-98115-84-9. EAN13: 9788898115846. [Google Scholar]
- Radan, M.; Parcina, A.; Burkul, F. Chemical composition and antioxidant activity of essential oil obtained from bitter orange peel (Citrus aurantium L.) using two methods. Croat. Chem. Acta 2018, 91, 125–128. [Google Scholar] [CrossRef]
- Bonesi, M.; Loizzo, M.R.; Leporini, M.; Tenuta, M.C.; Passalacqua, N.G.; Tundis, R. Comparative evaluation of petitgrain oils from six Citrus species alone and in combination as potential functional anti-radicals and antioxidant agents. Plant Biosyst. 2018, 152, 986–993. [Google Scholar] [CrossRef]
- Badalamenti, N.; Bruno, M.; Schicchi, R.; Geraci, A.; Leporini, M.; Gervasi, L.; Tundis, R.; Loizzo, M.R. Chemical Compositions and Antioxidant Activities of Essential Oils, and Their Combinations, Obtained from Flavedo By-Product of Seven Cultivars of Sicilian Citrus aurantium L. Molecules 2022, 27, 1580. [Google Scholar] [CrossRef]
- Maksoud, S.; Abdel-Massih, R.M.; Rajha, H.N.; Louka, N.; Chemat, F.; Barba, F.J.; Debs, E. Citrus aurantium L. active constituents, biological effects and extraction methods. an updated review. Molecules 2021, 26, 5832. [Google Scholar] [CrossRef] [PubMed]
- Ben Hsouna, A.; Hamdi, N.; Ben Halima, N.; Abdelkafi, S. Characterization of essential oil of Citrus aurantium L. flowers: Antimicrobial and antioxidant activities. J. Oleo Sci. 2013, 62, 763–772. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Biological activities and safety of Citrus spp. Essential Oils. Int. J. Mol. Sci. 2018, 19, 1966. [Google Scholar] [CrossRef] [PubMed]
- Moraes, T.M.; Kushima, H.; Moleiro, F.C.; Santos, R.C.; Machado Rocha, L.R.; Marques, M.O.; Vilegas, W.; Hiruma-Lima, C.A. Effects of limonene and essential oil from Citrus aurantium on gastric mucosa: Role of prostaglandins and gastric mucus secretion. Chem. Biol. Interact. 2009, 180, 499–505. [Google Scholar] [CrossRef]
- Narang, N.; Jiraungkoorskul, W. Anticancer Activity of Key Lime, Citrus aurantifolia. Pharmacogn. Rev. 2016, 10, 118–122. [Google Scholar]
- de Moraes Pultrini, A.; Almeida Galindo, L.; Costa, M. Effects of the essential oil from Citrus aurantium L. in experimental anxiety models in mice. Life Sci. 2006, 78, 1720–1725. [Google Scholar] [CrossRef]
- Fugh-Berman, A.; Myers, A. Citrus aurantium, an ingredient of dietary supplements marketed for weight loss: Current status of clinical and basic research. Exp. Biol. Med. 2004, 229, 698–704. [Google Scholar] [CrossRef]
- Russo, C.; Maugeri, A.; Lombardo, G.E.; Musumeci, L.; Barreca, D.; Rapisarda, A.; Cirmi, S.; Navarra, M. The second life of Citrus fruit waste: A valuable source of bioactive compounds. Molecules 2021, 26, 5991. [Google Scholar] [CrossRef]
- Nieto, G.; Fernández-López, J.; Pérez-Álvarez, J.A.; Peñalver, R.; Ros-Berruezo, G.; Viuda-Martos, M. Valorization of Citrus co-products: Recovery of bioactive compounds and application in meat and meat products. Plants 2021, 10, 1069. [Google Scholar] [CrossRef]
- FAOSTAT Database. Available online: http://faostat3.fao.org/home/index.html (accessed on 25 November 2024).
- González-Molina, E.; Domínguez-Perles, R.; Moreno, D.A.; GarcíaViguera, C. Natural bioactive compounds of Citrus limon for food and health. J. Pharm. Biomed. Anal. 2010, 51, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Palazzolo, E.; Guarrasi, V.; Aleo, A.; Mammina, C.; Moschetti, G.; Germanà, M.A. Inhibition of foodborne pathogen bacteria by essential oils extracted from Citrus fruits cultivated in Sicily. Food Contr. 2012, 26, 326–330. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A. Lipids and Essential Oils as Antimicrobial Agents; John Wiley & Sons, Ltd.: Sessex, UK, 2011; pp. 203–238. [Google Scholar]
- Luro, F.; Garcia Neves, C.; Costantino, G.; da Silva Gesteira, A.; Paoli, M.; Ollitrault, P.; Tomi, F.; Micheli, F.; Gibernau, M. Effect of Environmental Conditions on the Yield of Peel and Composition of Essential Oils from Citrus Cultivated in Bahia (Brazil) and Corsica (France). Agronomy 2020, 10, 1256. [Google Scholar] [CrossRef]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 2014, 153, 265–271. [Google Scholar] [CrossRef]
- Li, C.; Cai, Q.; Wu, X.; Tan, Z.; Huang, S.; Wei, C.; Zhang, W.; Chen, Z.; Zhang, L.; Xiang, X. Variation in Compositions and Biological Activities of Essential Oils from Four Citrus Species: Citrus limon, Citrus sinensis, Citrus paradisi, and Citrus reticulata. Chem. Biodivers. 2022, 19, e202100910. [Google Scholar] [CrossRef] [PubMed]
- Chutia, M.; Deka Bhuyan, P.; Pathak, M.G.; Sarma, T.C.; Boruah, P. Antifungal activity and chemical composition of Citrus reticulata Blanco essential oil against phytopathogens from North-East India. LWT 2009, 42, 777–780. [Google Scholar] [CrossRef]
- Fouad, H.A.; da Camara, C.A.G. Chemical composition and bioactivity of peel oils from Citrus aurantiifolia and Citrus reticulata and enantiomers of their major constituent against Sitophilus zeamais (Coleoptera: Curculionidae). J. Stored Prod. Res. 2017, 73, 30–36. [Google Scholar] [CrossRef]
- Hąc-Wydro, K.; Flasiński, M.; Romańczuk, K. Essential oils as food eco-preservatives: Model system studies on the effect of temperature on limonene antibacterial activity. Food Chem. 2017, 235, 127–135. [Google Scholar] [CrossRef]
- Han, Y.; Chen, W.; Sun, Z. Antimicrobial activity and mechanism of limonene against Staphylococcus aureus. J. Food Saf. 2021, 41, e12918. [Google Scholar] [CrossRef]
- Duckett, J.G.; Ligrone, R. A survey of diaspore liberation mechanism and germination patterns in mosses. J. Bryol. 1992, 17, 335–354. [Google Scholar] [CrossRef]
- Goode, J.A.; Stead, A.D.; Ligrone, R.; Duckett, J.G. Studies of protonemal morphogenesis in mosses IV. Aloina (Pottiales). J. Bryol. 1994, 18, 27–41. [Google Scholar] [CrossRef]
- Goode, J.A.; Stead, A.D.; Duckett, J.G. Redifferentiation of moss protonemata: An experimental and immunofluorescence study of brood cell formation. Can. J. Bot. 1993, 71, 1510–1519. [Google Scholar] [CrossRef]
- Basile, A.; Giordano, S.; Spagnuolo, V.; Alfano, F.; Castaldo Cobianchi, R. Effect of lead and colchicine on morphogenesis in proto- nemata of the moss Funaria hygrometrica. Ann. Bot. 1995, 76, 597–606. [Google Scholar] [CrossRef]
- Bopp, M.; Quader, H.; Thoni, C.; Sawidis, T.; Schnepf, E. Filament disruption in Funaria protonemata: Formation and disintegration of tmema cells. J. Plant Physiol. 1991, 137, 273–284. [Google Scholar] [CrossRef]
- Schnepf, E. Structure and development of tmema cells in protonemata of Funaria hygrometrica (Bryophyta). Criptogam. Bot. 1992, 3, 35–39. [Google Scholar]
- Rao, D.N. Response of bryophytes to air pollution. In Bryophyte Ecology; Smith, A.J., Ed.; Chapman & Hall: London, UK, 1982; pp. 445–471. [Google Scholar]
- El Sawi, S.A.; Ibrahim, M.E.; El-Rokiek, K.G.; El-Din, S.A.S. Allelopathic potential of essential oils isolated from peels of three citrus species. Ann. Agric. Sci. 2019, 64, 89–94. [Google Scholar] [CrossRef]
- Jalaei, Z.; Fattahi, M.; Aramideh, S. Allelopathic and insecticidal activities of essential oil of Dracocephalum kotschyi Boiss. from Iran: A new chemotype with highest limonene-10-al and limonene. Ind. Crops Prod. 2015, 73, 109–117. [Google Scholar] [CrossRef]
- Tang, J.S.; Jiang, C.Y.; Liu, Y.; Zhang, X.Y.; Shao, H.; Zhang, C. Allelopathic potential of volatile organic compounds released by Xanthium sibiricum Patrin ex Widder. Allelopath. J. 2019, 47, 233–242. [Google Scholar] [CrossRef]
- Vaughn, S.F. Phytotoxic and antimicrobial activity of 5,7-dihydroxychromone from Peanut shells. J. Chem. Ecol. 1995, 21, 107–115. [Google Scholar] [CrossRef]
- Basile, A.; Cobianchi, R.C.; Rigano, D.; Senatore, F.; Bruno, M.; Rosselli, S.; Conte, B.; Sorbo, S. Potential allelopathic activity of Sideritis italica (Miller) Greuter et Burdet essential oil. Plant Biosyst. 2011, 145, 241–247. [Google Scholar] [CrossRef]
- Basile, A.; Sorbo, S.; Giordano, S.; Ricciardi, L.; Ferrara, S.; Montesano, D.; Castaldo Cobianchi, R.; Vuotto, M.L.; Ferrara, L. Antibacterial and allelopathic activity of Castanea sativa Mill. leaves. Fitoterapia 2000, 71, S110–S116. [Google Scholar] [CrossRef]
- Gagliano Candela, R.; Ilardi, V.; Badalamenti, N.; Bruno, M.; Rosselli, S.; Maggi, F. Essential oil compositions of Teucrium fruticans, T. scordium subsp. scordioides and T. siculum growing in Sicily and Malta. Nat. Prod. Res. 2021, 35, 3460–3469. [Google Scholar] [CrossRef]
- Council of Europe (EDQM). European Pharmacopoeia, 6th ed.; EDQM: Strasbourg, France, 2008. [Google Scholar]
- Rigano, D.; Formisano, C.; Rosselli, S.; Badalamenti, N.; Bruno, M. GC and GC-MS Analysis of Volatile Compounds from Ballota nigra subsp. uncinata collected in Aeolian Islands, Sicily (Southern Italy). Nat. Prod. Commun. 2020, 15, 1934578X2092048. [Google Scholar] [CrossRef]
- Erickson, H.H.; Sherris, J.C. Antibiotic sensitivity testing report of international collaborative study. Acta Pathol. Microbiol. Scand. 1971, 217B, 1–9. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| No. | Compounds a | LRI b | LRI c | Area (%) d | ||||
|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | ||||
| 1 | Hexanal | 803 | 798 | - | - | - | t | - |
| 2 | 3-Hexen-1-ol | 861 | 857 | - | - | t | t | - |
| 3 | Nonane | 902 | 900 | - | - | - | t | - |
| 4 | α-Pinene | 936 | 938 | 3.12 ± 0.13 | 3.35 ± 0.15 | 1.46 ± 0.04 | 1.65 ± 0.05 | 3.69 ± 0.11 |
| 5 | β-Pinene | 978 | 980 | 1.83 ± 0.06 | 1.71 ± 0.05 | 0.79 ± 0.02 | 0.81 ± 0.03 | 2.02 ± 0.08 |
| 6 | β-Myrcene | 1003 | 1005 | 2.21 ± 0.08 | 2.09 ± 0.07 | 2.57 ± 0.08 | 2.62 ± 0.07 | 2.08 ± 0.06 |
| 7 | Limonene | 1067 | 1063 | 72.89 ± 3.06 | 70.74 ± 2.91 | 86.80 ± 3.38 | 82.22 ± 3.19 | 71.32 ± 2.94 |
| 8 | γ-Terpinene | 1083 | 1081 | 17.75 ± 0.57 | 18.26 ± 0.58 | 5.87 ± 0.24 | 8.99 ± 0.29 | 17.80 ± 0.64 |
| 9 | Terpinolene | 1095 | 1089 | 0.98 ± 0.04 | 0.89 ± 0.04 | 0.29 ± 0.01 | 0.40 ± 0.02 | 1.00 ± 0.03 |
| 10 | β-Linalool | 1109 | 1103 | t | 0.23 ± 0.00 | 1.11 ± 0.04 | 0.38 ± 0.01 | t |
| 11 | Nonanal | 1112 | 1008 | - | t | t | t | - |
| 12 | Camphor | 1147 | 1145 | - | - | t | - | t |
| 13 | β-Citronellal | 1158 | 1154 | - | t | t | t | t |
| 14 | Terpinen-4-ol | 1181 | 1183 | 0.22 ± 0.01 | 0.20 ± 0.00 | t | t | 0.25 ± 0.01 |
| 15 | p-Cymene-8-ol | 1188 | 1189 | - | - | - | t | - |
| 16 | α-Terpinol | 1196 | 1192 | 0.38 ± 0.01 | 0.43 ± 0.02 | t | t | 0.52 ± 0.02 |
| 17 | Decanal | 1209 | 1210 | t | 0.11 ± 0.00 | t | t | t |
| 18 | Octyl acetate | 1216 | 1215 | - | - | - | t | - |
| 19 | Thymol methyl ether | 1231 | 1228 | - | - | t | - | - |
| 20 | 2-Decenal | 1265 | 1267 | - | - | t | - | - |
| 21 | Perillal | 1272 | 1271 | - | t | t | t | - |
| 22 | 1-Decanol | 1277 | 1275 | - | - | - | - | t |
| 23 | p-Mentha-1(7),8(10)-dien-9-ol | 1289 | 1287 | - | t | - | - | - |
| 24 | Thymol | 1293 | 1292 | t | t | t | - | t |
| 25 | Undecanal | 1308 | 1306 | - | - | t | t | t |
| 26 | δ-Elemene | 1334 | 1337 | - | - | t | t | - |
| 27 | Citronellol acetate | 1352 | 1354 | - | - | - | t | - |
| 28 | Neryl acetate | 1361 | 1363 | - | - | t | t | - |
| 29 | α-Copaene | 1368 | 1370 | t | - | t | t | t |
| 30 | β-Elemene | 1383 | 1381 | 0.20 ± 0.00 | 0.74 ± 0.03 | t | 0.35 ± 0.01 | 0.28 ± 0.01 |
| 31 | β-Caryophyllene | 1417 | 1420 | t | t | - | t | t |
| 32 | α-Caryophyllene | 1445 | 1452 | t | - | t | t | - |
| 33 | α-Selinene | 1485 | 1483 | t | t | - | 0.21 ± 0.00 | - |
| 34 | (Z,E)-α-Farnesene | 1502 | 1497 | t | t | t | 0.54 ± 0.02 | t |
| 35 | δ-Cadinene | 1510 | 1508 | t | - | t | t | t |
| 36 | α-Sinensal | 1740 | 1745 | - | 0.22 ± 0.00 | - | - | t |
| Monoterpene Hydrocarbons | 98.78 ± 3.94 | 97.04 ± 3.80 | 97.78 ± 3.79 | 96.69 ± 3.65 | 97.91 ± 3.86 | |||
| Oxygenated Monoterpenes | 0.60 ± 0.02 | 0.74 ± 0.02 | 1.11 ± 0.04 | 0.38 ± 0.01 | 0.77 ± 0.03 | |||
| Sesquiterpene Hydrocarbons | 0.20 ± 0.00 | 0.74 ± 0.03 | - | 1.10 ± 0.03 | 0.28 ± 0.01 | |||
| Oxygenated Sesquiterpenes | - | 0.22 ± 0.00 | - | - | - | |||
| Others | - | 0.11 ± 0.00 | - | - | - | |||
| Total | 99.58 ± 3.96 | 98.85 ± 3.85 | 98.89 ± 3.83 | 98.17 ± 3.69 | 98.96 ± 3.90 | |||
| C1 | C2 | C3 | C4 | C5 | Lim | Ce | Pe | Te | |
|---|---|---|---|---|---|---|---|---|---|
| Staphylococcus aureus | 16 | 32 | 32 | 32 | 16 | 4 | R | 2 | 0.03 |
| Proteus vulgaris | 32 | 128 | 128 | 128 | 32 | 8 | R | 16 | 2 |
| Klebsiella pneumoniae | 128 | 128 | 256 | 256 | 128 | 8 | R | 0.1 | R |
| Enterobacter cloacae | 64 | 128 | 128 | 128 | 64 | 16 | R | R | R |
| Escherichia coli | 64 | 64 | 128 | 128 | 128 | 8 | R | 0.1 | 64 |
| Pseudomonas aeruginosa | 128 | 256 | 128 | 256 | 128 | 4 | R | R | 16 |
| Salmonella typhi | 128 | 256 | 256 | 256 | 128 | 4 | 0.5 | 1 | 4 |
| Enterobacter aerogens | 64 | 128 | 128 | 128 | 64 | 16 | R | R | R |
| Samples | R. sativus | T. muralis | |||
|---|---|---|---|---|---|
| Hypocotyl-Root Axis Length in cm | Length of the Hair Zone | Cell Number of Main Protonemata Filaments | Number of Brood Cells | Number of Tmema Cells | |
| control | 2 ± 0.1 a | 0.5 ± 0.1 a’ | 25 ± 2.3 a” | 0 | 0 |
| C1 a | 0 | 0 | 0 | 0 | 0 |
| C1 b | 0 | 0 | 0 | 0 | 0 |
| C1 c | 0.3 ± 0.1 b | 0 | 7 ± 1.3 b” | 1 ± 0.2 a’’’ | 0 |
| C1 d | 0.8 ± 0.4 c | 0 | 15 ± 2.6 c” | 3 ± 0.9 b’’’ | 1 ± 0.1 a’’’’ |
| C1 e | 1.7 ± 0.3 a | 0.3 ± 0.1 b’ | 22 ± 3.1 a” | 6 ± 1.1 c’’’ | 3 ± 0.2 b’’’’ |
| C2 a | 0 | 0 | 0 | 0 | 0 |
| C2 b | 0 | 0 | 0 | 0 | 0 |
| C2 c | 0.5 ± 0.1 b | 0 | 9 ± 1.8 b” | 1 ± 0.1 a’’’ | 0 |
| C2 d | 1.4 ± 0.4 c | 0.2 ± 0.1 b’ | 18 ± 2.2 c” | 2 ± 0.3 b’’’ | 1 ± 0.1 a’’’’ |
| C2 e | 1.9 ± 0.3 a | 0.3 ± 0.1 b’ | 24 ± 3.3 a” | 5 ± 0.4 c’’’ | 2 ± 0.3 c’’’’ |
| C3 a | 0 | 0 | 0 | 0 | 0 |
| C3 b | 0 | 0 | 0 | 0 | 0 |
| C3 c | 0.5 ± 0.1 b | 0 | 8 ± 1.2 b” | 1 ± 0.2 a’’’ | 0 |
| C3 d | 1.4 ± 0.4 c | 0.2 ± 0.1 b’ | 17 ± 2.3 c” | 3 ± 0.4 b’’’ | 1 ± 0.2 a’’’’ |
| C3 e | 1.8 ± 0.3 a | 0.3 ± 0.1 b’ | 24 ± 2.7 a” | 4 ± 0.3 c’’’ | 2 ± 0.3 c’’’’ |
| C4 a | 0 | 0 | 0 | 0 | 0 |
| C4 b | 0 | 0 | 0 | 0 | 0 |
| C4 c | 0.5 ± 0.1 b | 0 | 9 ± 2.1 b” | 1 ±0.2 a’’’ | 0 |
| C4 d | 1.5 ± 0.4 c | 0.1 ± 0.1 b’ | 16 ± 2.3 c” | 3 ± 0.5 b’’’ | 1 ± 0.4 a’’’’ |
| C4 e | 1.9 ± 0.3 a | 0.2 ± 0.1 b’ | 24 ± 2.5 a” | 4 ± 1.2 c’’’ | 2 ± 0.6 c’’’’ |
| C5 a | 0 | 0 | 0 | 0 | 0 |
| C5 b | 0 | 0 | 0 | 0 | 0 |
| C5 c | 0.4 ± 0.2 b | 0 | 6 ± 1.1 d” | 1 ± 0.3 a’’’ | 0 |
| C5 d | 0.9 ± 0.3 c | 0 | 13 ± 2.8 c” | 3 ± 0.7 b’’’ | 1 ± 0.1 a’’’’ |
| C5 e | 1.7 ± 0.4 a | 0.3 ± 0.1 b’ | 23 ± 2.2 a” | 5 ± 0.3 c’’’ | 2 ± 0.2 c’’’’ |
| Lim a, b, c | 0 | 0 | 0 | 0 | 0 |
| Lim d | 0.6 ± 0.3 b | 0.2 ± 0.1 b’ | 12 ± 1.8 c” | 3 ± 0.4 b’’’ | 1 ± 0.1 a’’’’ |
| Lim e | 1.2 ± 0.2 d | 0.3 ± 0.1 b’ | 19 ± 2.1 c” | 6 ± 1.3 c’’’ | 3 ± 0.2 b’’’’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geraci, A.; Postiglione, A.; Sgadari, F.; Schicchi, R.; Badalamenti, N.; Bruno, M.; Basile, A.; Dentato, M.; Maresca, V. Essential Oils Obtained from Sicilian Citrus reticulata Blanco By-Products: Antibacterial and Allelopathic Activity. Plants 2024, 13, 3527. https://doi.org/10.3390/plants13243527
Geraci A, Postiglione A, Sgadari F, Schicchi R, Badalamenti N, Bruno M, Basile A, Dentato M, Maresca V. Essential Oils Obtained from Sicilian Citrus reticulata Blanco By-Products: Antibacterial and Allelopathic Activity. Plants. 2024; 13(24):3527. https://doi.org/10.3390/plants13243527
Chicago/Turabian StyleGeraci, Anna, Alessia Postiglione, Francesco Sgadari, Rosario Schicchi, Natale Badalamenti, Maurizio Bruno, Adriana Basile, Martina Dentato, and Viviana Maresca. 2024. "Essential Oils Obtained from Sicilian Citrus reticulata Blanco By-Products: Antibacterial and Allelopathic Activity" Plants 13, no. 24: 3527. https://doi.org/10.3390/plants13243527
APA StyleGeraci, A., Postiglione, A., Sgadari, F., Schicchi, R., Badalamenti, N., Bruno, M., Basile, A., Dentato, M., & Maresca, V. (2024). Essential Oils Obtained from Sicilian Citrus reticulata Blanco By-Products: Antibacterial and Allelopathic Activity. Plants, 13(24), 3527. https://doi.org/10.3390/plants13243527